

Soil Properties of Different Planting Combinations of Zanthoxylum planispinum var. dintanensis Plantations and Their Effect on Stoichiometry

Abstract
:1. Introduction
2. Materials and Methods
2.1. Overview of the Research Site
2.2. Sample Plot Setting
2.3. Soil Sample Collection
2.4. Index Analysis Methods
2.5. Data Processing
3. Results
3.1. Soil Properties of Z. planispinum Plantations with Different Planting Combinations
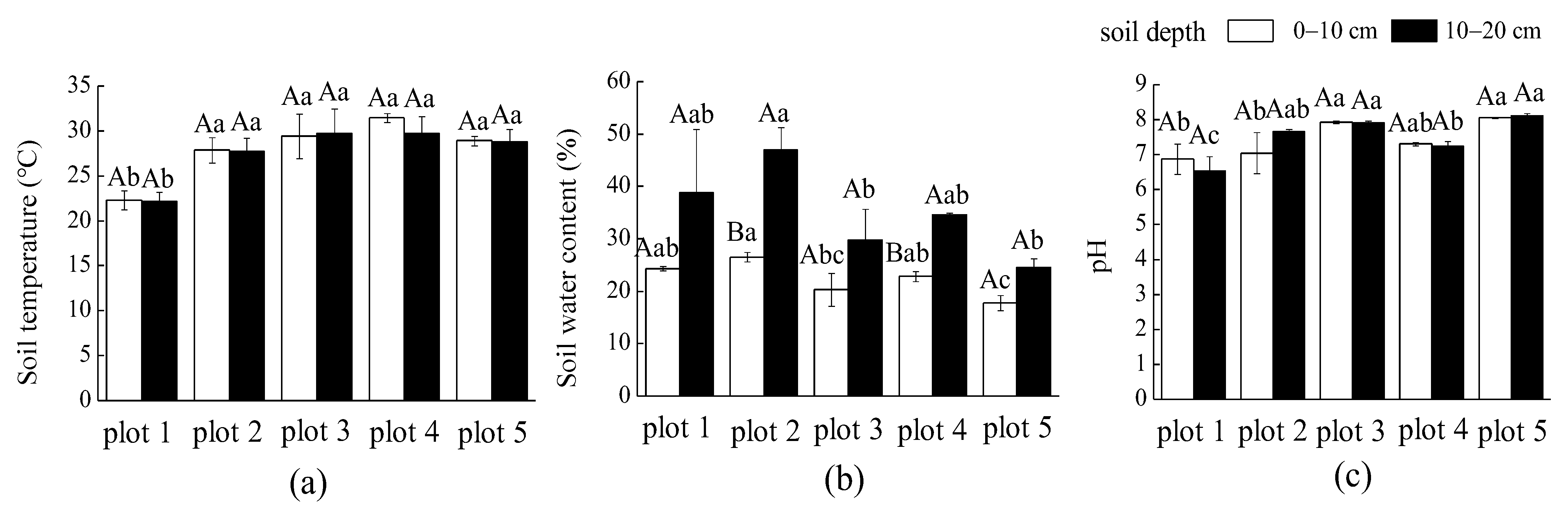
3.1.1. Soil Temperature, Water Content, and pH Value
3.1.2. Soil Available Nutrient Elements
3.2. Comprehensive Evaluation of Soil Quality of Z. planispinum Plantations with Different Planting Combinations
3.3. Soil Stoichiometry of Z. planispinum Plantations with Different Planting Combinations
3.4. Effect of Soil Quality on Stoichiometry
4. Discussion
4.1. Comparison of Soil Quality of Z. planispinum Plantations with Different Planting Combinations
4.2. The Driving Force of Soil Quality on Stoichiometry of Z. planispinum Plantations with Different Planting Combinations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bao, Q.; Yang, R.; Li, W.H.; Nie, C.J.; Lu, F.J. Soil ecological effects of different restoration models in the karst plateau gorge region. J. Soil Water Conserv. 2017, 31, 154–161. (In Chinese) [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, L.T.; Feng, Y.X.; Guo, S.S.; Pang, X.; Zhang, D.; Geng, Z.F.; Du, S.S. Insecticidal and repellent efficacy against stored-product insects of oxygenated monoterpenes and 2-dodecanone of the essential oil from Zanthoxyium planispinum var. dintanensis. Environ. Sci. Pollut. Res. 2019, 26, 24988–24997. [Google Scholar] [CrossRef] [PubMed]
- Huuskonena, S.; Domischb, T.; Finérb, L.; Hantulaa, J.; Hynynena, J.; Matalab, J.; Miinab, J.; Neuvonenc, S.; Nevalainenb, S.; Niemistöd, P.; et al. What is the potential for replacing monocultures with mixed-species stands to enhance ecosystem services in boreal forests in Fennoscandia? For. Ecol. Manag. 2021, 479, 118558. [Google Scholar] [CrossRef]
- Thoms, C.; Gattinger, A.; Jacob, M.; Thomas, F.M.; Gleixner, G. Direct and indirect effects of tree diversity drive soil microbial diversity in temperate deciduous forest. Soil Biol. Biochem. 2010, 42, 1558–1565. [Google Scholar] [CrossRef]
- Wang, L.X.; Pang, X.Y.; Li, N.; Qi, K.B.; Huang, J.S.; Yin, C.Y. Effects of vegetation type, fine and coarse roots on soil microbial communities and enzyme activities in eastern Tibetan plateau. Catena 2020, 194, 104694. [Google Scholar] [CrossRef]
- Zeng, Q.C.; Li, X.; Dong, Y.H.; An, S.S.; Darboux, F. Soil and plant components ecological stoichiometry in four steppe communities in the Loess Plateau of China. Catena 2016, 147, 481–488. [Google Scholar] [CrossRef]
- Ren, Z.; Niu, D.C.; Ma, P.P.; Wang, Y.; Fu, H.; Elser, J.J. Cascading influences of grassland degradation on nutrient limitation in a high mountain lake and its inflow streams. Ecology 2019, 100, e02755. [Google Scholar] [CrossRef]
- Lu, L.H.; Li, H.; Chen, L.; Ming, A.G.; Yang, Y.J. Effects of management patterns on soil physicochemical properties and ecological stoichiometric ratio in Pinus massoniana near-mature forest. Chin. J. Ecol. 2021, 40, 654–663. (In Chinese) [Google Scholar] [CrossRef]
- Stocker, C.M.; Bamberg, A.L.; Stumpf, L.; Monteiro, A.B.; Cardoso, J.H.; de Lima, A.C.R. Short-term soil physical quality improvements promoted by an agroforestry system. Agrofor. Syst. 2020, 94, 2053–2064. [Google Scholar] [CrossRef]
- Ma, Y.H.; Fu, S.L.; Zhang, X.P.; Zhao, K.; Chen, H.Y.H. Intercropping improves soil nutrient availability, soil enzyme activity and tea quantity and quality. Appl. Soil Ecol. 2017, 119, 171–178. [Google Scholar] [CrossRef]
- Zhou, L.; Sun, Y.J.; Saeed, S.; Zhang, B.; Luo, M. The difference of soil properties between pure and mixed Chinese fir (Cunninghamia lanceolata) plantations depends on tree species. Glob. Ecol. Conserv. 2020, 22, e01009. [Google Scholar] [CrossRef]
- Chen, J.Z.; Rong, L.; Xiong, K.N. Effect of Interplanting Patterns on Concentrations of Soil Mineral Elements of Zanthoxylum bungeanum. Southwest China J. Agric. Sci. 2019, 32, 763–769. (In Chinese) [Google Scholar] [CrossRef]
- Hou, X.L.; Han, H.; Tigabu, M.; Cai, L.P.; Meng, F.R.; Liu, A.Q.; Ma, X.Q. Changes in soil physico-chemical properties following vegetation restoration mediate bacterial community composition and diversity in Changting, China. Ecol. Eng. 2019, 138, 171–179. [Google Scholar] [CrossRef]
- Raiesi, F. A minimum data set and soil quality index to quantify the effect of land use conversion on soil quality and degradation in native rangelands of upland arid and semiarid regions. Ecol. Indic. 2017, 75, 307–320. [Google Scholar] [CrossRef]
- Rowley, M.C.; Grand, S.; Verrecchia, E.P. Calcium-mediated stabilisation of soil organic carbon. Biogeochemistry 2018, 137, 27–49. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Ul Hassan, M.; Nadeem, F.; Wu, L.Q.; Zhang, F.S.; Li, X.X. Magnesium Fertilization Improves Crop Yield in Most Production Systems: A Meta-Analysis. Front. Plant Sci. 2020, 10, 1727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Q.Q.; Wu, Y.H.; Bing, H.J.; Zhou, J.; Wang, J.P. Vegetation type rather than climate modulates the variation in soil enzyme activities and stoichiometry in subalpine forests in the eastern Tibetan Plateau. Geoderma 2020, 374, 114424. [Google Scholar] [CrossRef]
- Huang, R.; Lan, T.; Song, X.; Li, J.; Ling, J.; Deng, O.P.; Wang, C.Q.; Gao, X.S.; Li, Q.Q.; Tang, X.Y.; et al. Soil labile organic carbon impacts C:N:P stoichiometry in urban park green spaces depending on vegetation types and time after planting. Appl. Soil Ecol. 2021, 163, 103926. [Google Scholar] [CrossRef]
- Zou, J.; Yu, L.F.; Huang, Z.S. Variation of Leaf Carbon Isotope in Plants in Different Lithological Habitats in a Karst Area. Forest 2019, 10, 356. [Google Scholar] [CrossRef] [Green Version]
- Wei, C.S.; Zuo, Z.L. Analysis and countermeasure research on the cause of the decline of Zanthoxyhum planispiunum var dingtanensis industry. Guizhou For. Sci. Technol. 2016, 44, 60–64. (In Chinese) [Google Scholar] [CrossRef]
- Li, Y.T.; Yu, Y.H.; Song, Y.P. Stoichiometry of Soil, Microorganisms, and Extracellular Enzymes of Zanthoxylum planispinum var. dintanensis Plantations for Different Allocations. Agronomy 2022, 12, 1709. [Google Scholar] [CrossRef]
- Bao, S.D. Soil Agrochemical Analysis, 3rd ed.; China Agriculture Press: Beijing, China, 2000; pp. 22–173. (In Chinese) [Google Scholar]
- Lu, R.K. Methods for Soil and Agriculture Chemistry Analysis, 1st ed.; Chinese Agricultural Science and Technology Press: Beijing, China, 1999. (In Chinese) [Google Scholar]
- Sinsabaugh, R.L.; Hill, B.H.; Shah, J.J.F. Ecoenzymatic stoichiometry of microbial organic nutrient acquisition in soil and sediment. Nature 2009, 462, 795–798. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, G.; Loffredo, E.; Senesi, N. Anticlastogenic, antitoxic and sorption effects of humic substances on the mutagen maleic hydrazide tested in leguminous plants. Eur. J. Soil Sci. 2004, 55, 449–458. [Google Scholar] [CrossRef]
- Ferrara, G.; Loffredo, E.; Simeone, R.; Senesi, N. Evaluation of Antimutagenic and Desmutagenic Effects of Humic and Fulvic Acids on Root Tips of Vicia faba. Environ. Toxicol. 2000, 15, 513–517. [Google Scholar] [CrossRef]
- Jin, Z.Z.; Lei, J.Q.; Xu, X.W.; Li, S.Y.; Zhao, S.F.; Qiu, Y.Z.; Xu, B. Evaluation of soil fertility of the shelter-Forest land along the Tarim Desert Highway. Chin. Sci. Bull. 2008, 53, 125–136. [Google Scholar] [CrossRef] [Green Version]
- Joswig, J.S.; Wirth, C.; Schuman, M.C.; Kattge, J.; Reu, B.; Wright, I.J.; Sippel, S.D.; Rüger, N.; Richter, R.; Schaepman, M.E.; et al. Climatic and soil factors explain thetwo-dimensional spectrum of global planttrait variation. Nat. Ecol. Evol. 2022, 6, 36–50. [Google Scholar] [CrossRef]
- Schloter, M.; Nannipieri, P.; Sørensen, S.J.; van Elsas, J.D. Microbial indicators for soil quality. Biol. Fertil. Soils 2018, 54, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Smith, M.S.; Fridley, J.D.; Goebe, M.; Bauerle, T.L. Links between Belowground and Aboveground Resource-Related Traits Reveal Species Growth Strategies that Promote Invasive Advantages. PLoS ONE 2014, 9, e104189. [Google Scholar] [CrossRef]
- López-Vicente, M.; Calvo-Seas, E.; Álvarez, S.; Cerdà, A. Effectiveness of Cover Crops to Reduce Loss of Soil Organic Matter in a Rainfed Vineyard. Land 2020, 9, 230. [Google Scholar] [CrossRef]
- Ch’ng, H.Y.; Ahmed, O.H.; Majid, N.M.A. Improving phosphorus availability in an acid soil using organic amendments produced from agroindustrial wastes. Sci. World J. 2014, 506356, 1–6. [Google Scholar] [CrossRef]
- Green, S.M.; Dungait, J.A.J.; Tu, C.L.; Buss, H.L.; Sanderson, N.; Hawkes, S.J.; Xing, K.X.; Yue, F.J.; Hussey, V.L.; Peng, J.; et al. Soil functions and ecosystem services research in the Chinese karst Critical Zone. Chem. Geol. 2019, 527, 119107. [Google Scholar] [CrossRef] [Green Version]
- White, P.J.; Broadley, M.R. Calcium in plants. Ann. Bot. 2003, 92, 487–511. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.K.; Wang, S.L. Soil microbial properties and nutrients in pure and mixed Chinese fir plantations. J. For. Res. 2008, 19, 131–135. [Google Scholar] [CrossRef]
- Udawatta, R.P.; Kremer, R.J.; Nelson, K.A.; Jose, S.; Bardhan, S. Soil quality of a mature alley cropping agroforestry system in temperate North America. Commun. Soil Sci. Plant Anal. 2014, 45, 1539–2551. [Google Scholar] [CrossRef]
- Qi, K.B.; Pang, X.Y.; Yang, B.; Bao, W.K. Soil carbon, nitrogen and phosphorus ecological stoichiometry shifts with tree species in subalpine plantations. PeerJ 2020, 8, e9702. [Google Scholar] [CrossRef]
- Rawata, M.; Arunachalam, K.; Arunachalam, A.; Alatalo, J.M.; Kumar, U.; Simon, B.; Hufnagel, L.; Micheli, E.; Pandey, R. Relative contribution of plant traits and soil properties to the functioning of a temperate forest ecosystem in the Indian Himalayas. Catena 2020, 194, 104671. [Google Scholar] [CrossRef]
- Tian, L.M.; Zhao, L.; Wu, X.D.; Hu, G.J.; Fang, H.B.; Zhao, Y.H.; Shen, Y.; Chen, J.; Wu, J.C.; Li, W.P.; et al. Variations in soil nutrient availability across Tibetan grassland from the 1980s to 2010s. Geoderma 2019, 338, 197–205. [Google Scholar] [CrossRef]
- Tian, D.S.; Reich, P.B.; Chen, H.Y.H.; Xiang, Y.Z.; Luo, Y.Q.; Shen, Y.; Meng, C.; Han, W.X.; Niu, S.L. Global changes alter plant multi-element stoichiometric coupling. New Phytol. 2019, 221, 807–817. [Google Scholar] [CrossRef]
- Muhammad, I.; Yang, L.; Ahmad, S.; Zeeshan, M.; Farooq, S.; Ali, I.; Khan, A.; Zhou, X.B. Irrigation and Nitrogen Fertilization Alter Soil Bacterial Communities, Soil Enzyme Activities, and Nutrient Availability in Maize Crop. Front. Microbiol. 2022, 13, 833758. [Google Scholar] [CrossRef]
- Palansooriya, K.N.; Shaheen, S.M.; Chen, S.S.; Tsang, D.C.W.; Hashimoto, Y.; Hou, D.Y.; Bolan, N.S.; Rinklebe, J.; Ok, Y.S. Soil amendments for immobilization of potentially toxic elements in contaminated soils: A critical review. Environ. Int. 2020, 134, 105046. [Google Scholar] [CrossRef]
- Wei, L.; Ge, T.D.; Zhu, Z.K.; Luo, Y.; Yang, Y.H.; Xiao, M.L.; Yan, Z.F.; Li, Y.H.; Wu, J.S.; Kuzyakov, Y. Comparing carbon and nitrogen stocks in paddy and upland soils: Accumulation, stabilization mechanisms, and environmental drivers. Geoderma 2021, 398, 115121. [Google Scholar] [CrossRef]
- Khan, K.S.; Joergensen, R.G. Stoichiometry of the soil microbial biomass in response to amendments with varying C/N/P/S ratios. Biol. Fertil. Soils 2019, 55, 265–274. [Google Scholar] [CrossRef]
- Wu, L.K.; Lin, X.M.; Lin, W.X. Advances and perspective in research on plant-soil-microbe interactions mediated by root exudates. Chin. J. Plant Ecol. 2014, 38, 298–310. (In Chinese) [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Plantation Type | Species Combinations | Longitude | Latitude | Growing Area (ha) | Altitude (m asl) | Density (m) | Height (m) | Crown Width (m) | Coverage (%) |
|---|---|---|---|---|---|---|---|---|---|
| Plot 1 | Z. planispinum + P. salicina | 105°40′28.33″ E | 25°37′57.41″ N | 1.34 | 764 | 3 × 3 | 3.5 | 2 × 2.3 | 70 |
| Plot 2 | Z. planispinum + S. tonkinensis | 105°40′19.79″ E | 25°39′25.75″ N | 0.67 | 728 | 2 × 2 | 2.0 | 1.2 × 1.8 | 60 |
| Plot 3 | Z. planispinum + A. hypogaea | 105°38′36.32″ E | 25°39′23.64″ N | 0.67 | 791 | 2 × 2 | 2.5 | 2.5 × 2.8 | 85 |
| Plot 4 | Z. planispinum + L. japonica | 105°38′36.35″ E | 25°39′22.29″ N | 6.67 | 814 | 3.5 × 3 | 2.5 | 1.5 × 2.5 | 70 |
| Plot 5 | Z. planispinum | 105°38′35.64″ E | 25°39′23.35″ N | 33.35 | 788 | 3 × 4 | 2.2 | 2.5 × 2.3 | 65 |
| Soil Parameters | Soil Depth (cm) | Plot 1 | Plot 2 | Plot 3 | Plot 4 | Plot 5 |
|---|---|---|---|---|---|---|
| Soil organic carbon (g·kg−1) | 0–10 | 36.05 ± 9.83 Aab | 32.45 ± 2.05 Aab | 28.75 ± 15.49 Ab | 59.55 ± 16.48 Aa | 27.05 ± 1.34 Ab |
| 10–20 | 39.40 ± 4.81 Aa | 26.35 ± 0.92 Aa | 27.60 ± 12.59 Aa | 42.10 ± 10.18 Aa | 25.95 ± 3.04 Aa | |
| Total nitrogen (g·kg−1) | 0–10 | 36.05 ± 9.83 Aab | 2.78 ± 0.04 Ab | 2.79 ± 0.93 Ab | 5.07 ± 0.25 Aa | 2.84 ± 0.23 Ab |
| 10–20 | 39.40 ± 4.81 Aa | 2.51 ± 0.11 Ab | 2.78 ± 0.56 Ab | 4.13 ± 0.64 Aa | 2.69 ± 0.23 Ab | |
| Total phosphorus (g·kg−1) | 0–10 | 1.46 ± 0.13 Aab | 0.99 ± 0.21 Ab | 1.17 ± 0.52 Aab | 1.73 ± 0.20 Aa | 1.32 ± 0.01 Aab |
| 10–20 | 1.27 ± 0.08 Aa | 0.66 ± 0.13 Ab | 1.04 ± 0.35 Aab | 1.32 ± 0.15 Aa | 1.21 ± 0.06 Aa | |
| Total potassium (g·kg−1) | 0–10 | 7.15 ± 0.35 Ab | 6.64 ± 2.79 Ab | 11.65 ± 0.78 Aa | 11.65 ± 0.50 Aa | 11.80 ± 0.57 Aa |
| 10–20 | 6.75 ± 1.04 Ac | 5.58 ± 0.23 Ac | 13.00 ± 0.28 Aa | 12.10 ± 0.57 Aa | 9.96 ± 0.62 Ab | |
| Total calcium (g·kg−1) | 0–10 | 1.20 ± 0.85 Aa | 2.35 ± 2.05 Aa | 6.05 ± 6.72 Aa | 1.65 ± 0.21 Aa | 9.40 ± 5.37 Aa |
| 10–20 | 1.25 ± 0.50 Ab | 1.95 ± 1.34 Ab | 4.65 ± 4.60 Aab | 2.10 ± 0.57 Ab | 8.90 ± 2.97 Aa | |
| Total magnesium (g·kg−1) | 0–10 | 5.30 ± 2.55 Ab | 5.65 ± 3.32 Ab | 7.85 ± 1.91 Ab | 8.45 ± 1.06 Ab | 15.45 ± 0.21 Aa |
| 10–20 | 5.10 ± 0.57 Ab | 4.25 ± 0.21 Ab | 11.45 ± 4.88 Aa | 8.75 ± 1.20 Aab | 12.25 ± 2.05 Aa |
| Soil Parameters | Soil Depth (cm) | Plot 1 | Plot 2 | Plot 3 | Plot 4 | Plot 5 |
|---|---|---|---|---|---|---|
| Fungal concentration (×103 CFU·g−1) | 0–10 | 11.75 ± 4.60 Aa | 11.00 ± 1.41 Aa | 9.60 ± 4.81 Aa | 12.00 ± 1.41 Aa | 8.40 ± 2.26 Aa |
| 10–20 | 12.50 ± 3.54 Aa | 9.00 ± 2.83 Aa | 5.95 ± 0.21 Aa | 12.20 ± 5.37 Aa | 7.45 ± 3.61 Aa | |
| Bacterial concentration (×106 CFU·g−1) | 0–10 | 2.35 ± 0.09 Abc | 5.30 ± 0.71 Aa | 3.00 ± 0.99 Abc | 3.15 ± 0.49 Ab | 1.20 ± 0.00 Ac |
| 10–20 | 4.85 ± 1.06 Aa | 1.80 ± 0.42 Bbc | 2.20 ± 1.56 Aabc | 4.40 ± 1.13 Aab | 1.55 ± 0.78 Ac | |
| Actinomycete concentration (×105 CFU·g−1) | 0–10 | 21.50 ± 12.02 Aa | 26.50 ± 0.71 Aa | 14.65 ± 6.15 Aa | 25.00 ± 7.07 Aa | 9.30 ± 0.57 Aa |
| 10–20 | 26.50 ± 13.44 Aa | 9.95 ± 0.07 Ba | 15.00 ± 7.07 Aa | 32.15 ± 11.10 Aa | 10.10 ± 5.52 Aa | |
| Microbial biomass carbon (mg·kg−1) | 0–10 | 265.00 ± 15.56 Aa | 252.00 ± 8.49 Aa | 249.50 ± 20.51 Aa | 245.00 ± 19.80 Aa | 262.00 ± 9.90 Aa |
| 10–20 | 221.00 ± 5.66 Ab | 257.50 ± 3.54 Aab | 254.50 ± 14.85 Aab | 280.50 ± 23.33 Aa | 262.50 ± 43.13 Aab | |
| Microbial biomass nitrogen (mg·kg−1) | 0–10 | 12.75 ± 3.18 Aa | 14.90 ± 1.27 Aa | 15.65 ± 0.35 Aa | 13.70 ± 0.42 Aa | 13.30 ± 1.56 Aa |
| 10–20 | 12.05 ± 0.21 Aa | 12.25 ± 1.34 Aa | 13.10 ± 1.56 Aa | 14.10 ± 1.70 Aa | 14.85 ± 1.91 Aa | |
| Microbial biomass phosphorus (mg·kg−1) | 0–10 | 120.00 ± 21.21 Ab | 150.00 ± 5.66 Aab | 152.00 ± 14.14 Aab | 161.00 ± 9.90 Aa | 126.00 ± 11.31 Ab |
| 10–20 | 136.00 ± 25.46 Aa | 139.00 ± 4.24 Aa | 144.00 ± 2.83 Aa | 148.00 ± 16.97 Aa | 152.00 ± 4.24 Aa | |
| β-1,4-glucosidase [μmol·(min·g)−1] | 0–10 | 6.38 ± 0.03 Aa | 6.24 ± 0.33 Aa | 6.49 ± 0.32 Aa | 6.38 ± 0.74 Aa | 6.28 ± 0.74 Aa |
| 10–20 | 5.67 ± 0.02 Bb | 6.05 ± 0.17 Ab | 6.18 ± 0.28 Ab | 6.96 ± 0.02 Aa | 6.30 ± 0.45 Ab | |
| β-1,4-N-acetylglucosaminidase [μmol·(min·g)−1] | 0–10 | 35.39 ± 0.17 Aa | 34.41 ± 2.25 Aa | 36.12 ± 2.25 Aa | 35.39 ± 5.19 Aa | 34.66 ± 5.20 Aa |
| 10–20 | 30.37 ± 0.17 Bb | 33.07 ± 1.21 Ab | 33.98 ± 1.99 Ab | 39.43 ± 0.18 Aa | 34.78 ± 3.12 Ab | |
| Leucine aminopeptidase [μmol·(min·g)−1] | 0–10 | 30.14 ± 0.12 Aa | 29.47 ± 1.55 Aa | 30.64 ± 1.54 Aa | 30.14 ± 3.56 Aa | 34.63 ± 3.51 Aa |
| 10–20 | 26.70 ± 0.12 Bb | 28.54 ± 0.83 Ab | 29.18 ± 1.36 Ab | 32.91 ± 0.12 Aa | 29.72 ± 2.14 Ab | |
| Acid phosphatase [μmol·(min·g)−1] | 0–10 | 7.51 ± 0.03 Aa | 7.35 ± 0.35 Aa | 7.63 ± 0.36 Aa | 7.51 ± 0.83 Aa | 7.39 ± 0.83 Aa |
| 10–20 | 6.70 ± 0.03 Bb | 7.14 ± 0.19 Ab | 7.29 ± 0.32 Ab | 8.15 ± 0.03 Aa | 7.41 ± 0.50 Ab |
| Soil Parameters | Load Matrix of Principal Components | |||||
|---|---|---|---|---|---|---|
| PC1 | PC2 | PC3 | PC4 | PC5 | PC6 | |
| pH | −0.579 | 0.673 | 0.081 | 0.194 | −0.036 | −0.016 |
| Soil water content | −0.178 | −0.423 | −0.258 | 0.073 | 0.031 | 0.768 |
| Soil temperature | −0.020 | 0.526 | 0.303 | 0.648 | 0.145 | 0.060 |
| Soil organic carbon | 0.916 | −0.003 | −0.097 | 0.279 | 0.155 | 0.042 |
| Total nitrogen | 0.924 | 0.067 | 0.039 | 0.267 | 0.120 | 0.043 |
| Total phosphorus | 0.828 | 0.352 | 0.056 | −0.035 | 0.155 | −0.279 |
| Total potassium | 0.075 | 0.564 | 0.359 | 0.415 | −0.041 | −0.341 |
| Total calcium | −0.248 | 0.865 | 0.015 | −0.112 | −0.108 | −0.013 |
| Total magnesium | −0.108 | 0.874 | 0.096 | 0.074 | −0.073 | −0.303 |
| Available nitrogen | 0.939 | −0.086 | 0.054 | 0.164 | 0.072 | 0.058 |
| Soil available phosphorus | 0.745 | −0.246 | 0.062 | −0.241 | −0.194 | −0.146 |
| Available potassium | 0.616 | −0.361 | 0.042 | −0.413 | −0.482 | −0.138 |
| Available calcium | 0.189 | 0.622 | 0.031 | 0.602 | 0.325 | 0.206 |
| Available magnesium | 0.169 | 0.850 | 0.224 | 0.222 | 0.014 | −0.005 |
| Fungi concentration | 0.636 | −0.235 | 0.177 | −0.184 | 0.189 | 0.455 |
| Bacterial concentration | 0.314 | −0.410 | −0.035 | 0.035 | 0.793 | 0.120 |
| Actinomycetes concentration | 0.527 | −0.207 | 0.071 | −0.087 | 0.746 | 0.216 |
| Microbial biomass carbon | 0.077 | 0.364 | 0.505 | −0.225 | −0.044 | 0.525 |
| Microbial biomass nitrogen | −0.169 | 0.308 | 0.236 | 0.016 | 0.729 | −0.253 |
| Microbial biomass phosphorus | 0.140 | −0.068 | 0.154 | 0.844 | −0.111 | −0.119 |
| β-1,4-glucosidase | 0.038 | 0.093 | 0.969 | 0.145 | 0.059 | −0.027 |
| β-1,4-N-acetylglucosaminidase | 0.037 | 0.092 | 0.969 | 0.144 | 0.061 | −0.028 |
| Leucine aminopeptidase | −0.028 | 0.561 | 0.562 | −0.168 | 0.038 | −0.140 |
| Acid phosphatase | 0.035 | 0.093 | 0.969 | 0.143 | 0.059 | −0.029 |
| Eigenvalue | 6.918 | 5.883 | 2.629 | 2.404 | 1.587 | 1.266 |
| Variance contribution rate (%) | 23.104 | 20.899 | 16.176 | 9.864 | 9.430 | 6.721 |
| Cumulative variance contribution rate (%) | 23.104 | 44.002 | 60.179 | 70.043 | 79.473 | 86.194 |
| Plantation Type | Factor Score | Soil Quality Parameters | Rank | |||||
|---|---|---|---|---|---|---|---|---|
| F1 | F2 | F3 | F4 | F5 | F6 | |||
| Plot 1 | 1.589 | −3.042 | −2.419 | −2.678 | −0.625 | 0.652 | −0.939 | 5 |
| Plot 2 | −1.198 | −1.746 | −1.471 | −0.703 | 0.001 | 1.542 | −0.845 | 4 |
| Plot 3 | −1.235 | 0.630 | 0.430 | 0.801 | −0.260 | −1.023 | −0.098 | 3 |
| Plot 4 | 2.587 | 1.046 | 2.578 | 2.237 | 2.095 | 0.386 | 1.678 | 1 |
| Plot 5 | −1.742 | 3.113 | 0.882 | 0.343 | −1.210 | −1.558 | 0.206 | 2 |
| Soil Parameters | Soil Depth (cm) | Plot 1 | Plot 2 | Plot 3 | Plot 4 | Plot 5 |
|---|---|---|---|---|---|---|
| C:N | 0–10 | 10.15 ± 1.49 Aa | 11.70 ± 0.89 Aa | 10.51 ± 1.05 Aa | 11.69 ± 2.68 Aa | 9.54 ± 0.29 Aa |
| 10–20 | 11.15 ± 0.09 Aa | 10.50 ± 0.11 Aa | 9.69 ± 2.59 Aa | 10.12 ± 0.91 Aa | 9.65 ± 0.29 Aa | |
| C:P | 0–10 | 25.08 ± 8.92 Aab | 33.45 ± 4.88 Aab | 25.70 ± 0.57 Aab | 34.10 ± 5.62 Aa | 20.50 ± 1.24 Ab |
| 10–20 | 30.97 ± 1.72 Aab | 40.85 ± 7.13 Aa | 26.08 ± 3.45 Ab | 31.78 ± 4.15 Aab | 21.63 ± 3.67 Ab | |
| N:P | 0–10 | 2.43 ± 0.52 Aa | 2.88 ± 0.64 Aa | 2.46 ± 0.30 Aa | 2.94 ± 0.19 Aa | 2.15 ± 0.19 Aa |
| 10–20 | 2.78 ± 0.18 Ab | 3.89 ± 0.64 Aa | 2.74 ± 0.38 Ab | 3.13 ± 0.13 Aab | 2.24 ± 0.31 Ab | |
| C:K | 0–10 | 5.08 ± 1.63 Aa | 5.29 ± 1.91 Aa | 2.60 ± 1.26 Aa | 5.15 ± 1.63 Aa | 2.30 ± 0.22 Aa |
| 10–20 | 5.86 ± 0.19 Aa | 4.73 ± 0.36 Aab | 2.11 ± 0.92 Ac | 3.50 ± 1.01 Abc | 2.60 ± 0.14 Ac | |
| N:K | 0–10 | 0.49 ± 0.09 Aa | 0.46 ± 0.20 Aa | 0.24 ± 0.10 Aa | 0.44 ± 0.04 Aa | 0.24 ± 0.03 Aa |
| 10–20 | 0.53 ± 0.01 Aa | 0.45 ± 0.04 Aa | 0.21 ± 0.04 Ac | 0.34 ± 0.07 Ab | 0.27 ± 0.01 Abc | |
| P:K | 0–10 | 0.20 ± 0.01 Aa | 0.16 ± 0.03 Aab | 0.10 ± 0.05 Ab | 0.15 ± 0.02 Aab | 0.11 ± 0.00 Ab |
| 10–20 | 0.19 ± 0.02 Aa | 0.12 ± 0.03 Ab | 0.08 ± 0.02 Ab | 0.11 ± 0.02 Ab | 0.12 ± 0.01 Ab | |
| C:Ca | 0–10 | 50.38 ± 15.96 Aa | 25.39 ± 3.95 Aabc | 16.41 ± 0.58 Abc | 37.04 ± 14.75 Aab | 4.59 ± 0.10 Ac |
| 10–20 | 37.62 ± 14.05 Aa | 16.81 ± 5.60 Ab | 14.28 ± 1.31 Ab | 20.12 ± 0.57 Aab | 4.26 ± 1.07 Ab | |
| Ca:Mg | 0–10 | 0.16 ± 0.02 Ac | 0.24 ± 0.03 Ab | 0.23 ± 0.04 Abc | 0.20 ± 0.00 Abc | 0.38 ± 0.03 Aa |
| 10–20 | 0.22 ± 0.08 Ab | 0.39 ± 0.10 Aab | 0.20 ± 0.03 Ab | 0.25 ± 0.10 Ab | 0.52 ± 0.16 Aa | |
| MBC:MBN | 0–10 | 21.61 ± 6.62 Aa | 16.95 ± 0.88 Aa | 15.96 ± 1.67 Aa | 17.87 ± 0.89 Aa | 19.88 ± 3.07 Aa |
| 10–20 | 18.34 ± 0.14 Aa | 21.17 ± 2.61 Aa | 19.63 ± 3.46 Aa | 19.94 ± 0.75 Aa | 17.64 ± 0.64 Aa | |
| MBC:MBP | 0–10 | 2.23 ± 0.27 Aa | 1.69 ± 0.12 Abc | 1.64 ± 0.01 Ac | 1.53 ± 0.21 Ac | 2.09 ± 0.11 Aab |
| 10–20 | 1.66 ± 0.35 Aa | 1.86 ± 0.04 Aa | 1.77 ± 0.14 Aa | 1.92 ± 0.37 Aa | 1.74 ± 0.33 Aa | |
| MBN:MBP | 0–10 | 0.11 ± 0.04 Aa | 0.10 ± 0.01 Aa | 0.11 ± 0.01 Aa | 0.09 ± 0.01 Aa | 0.11 ± 0.02 Aa |
| 10–20 | 0.09 ± 0.01 Aa | 0.09 ± 0.01 Aa | 0.09 ± 0.01 Aa | 0.10 ± 0.02 Aa | 0.10 ± 0.01 Aa | |
| BG:(NAG + LAP) | 0–10 | 0.10 ± 0.00 Aa | 0.10 ± 0.00 Aa | 0.10 ± 0.00 Aa | 0.10 ± 0.00 Aa | 0.09 ± 0.01 Aa |
| 10–20 | 0.10 ± 0.00 Aa | 0.10 ± 0.00 Aa | 0.10 ± 0.00 Aa | 0.10 ± 0.00 Aa | 0.10 ± 0.00 Aa | |
| BG:AP | 0–10 | 0.85 ± 0.00 Aa | 0.85 ± 0.00 Aa | 0.85 ± 0.00 Aa | 0.85 ± 0.00 Aa | 0.85 ± 0.00 Aa |
| 10–20 | 0.85 ± 0.00 Aa | 0.85 ± 0.00 Aa | 0.85 ± 0.00 Aa | 0.85 ± 0.00 Aa | 0.85 ± 0.00 Aa | |
| (NAG + LAP):AP | 0–10 | 8.73 ± 0.01 Aa | 8.69 ± 0.10 Aa | 8.75 ± 0.08 Aa | 8.72 ± 0.20 Aa | 9.42 ± 0.83 Aa |
| 10–20 | 8.52 ± 0.01 Bb | 8.63 ± 0.06 Ab | 8.67 ± 0.08 Ab | 8.88 ± 0.01 Aa | 8.7 ± 0.13 Aab |
| Soil Parameters | Importance Ranking | Factor Interpretation (%) | p | F |
|---|---|---|---|---|
| Total magnesium | 1 | 27.3 | 0.002 | 10.5 |
| Total calcium | 2 | 26.0 | 0.002 | 9.8 |
| pH | 3 | 23.9 | 0.002 | 8.8 |
| Leucine aminopeptidase | 4 | 23.4 | 0.002 | 8.6 |
| Available potassium | 5 | 18.9 | 0.002 | 6.5 |
| Total potassium | 6 | 18.9 | 0.002 | 6.5 |
| Soil organic carbon | 7 | 15.2 | 0.002 | 5.0 |
| Fungi concentration | 8 | 15.1 | 0.002 | 5.0 |
| Bacterial concentration | 9 | 14.7 | 0.002 | 4.8 |
| Available magnesium | 10 | 14.1 | 0.002 | 4.6 |
| Available nitrogen | 11 | 14.1 | 0.006 | 4.6 |
| Microbial biomass phosphorus | 12 | 13.2 | 0.002 | 4.2 |
| Soil water content | 13 | 13.1 | 0.004 | 4.2 |
| Soil available phosphorus | 14 | 13.0 | 0.008 | 4.2 |
| Actinomycetes concentration | 15 | 12.6 | 0.006 | 4.0 |
| Microbial biomass nitrogen | 16 | 11.8 | 0.006 | 3.7 |
| Acid phosphatase | 17 | 10.5 | 0.010 | 3.3 |
| β-1,4-glucosidase | 18 | 10.5 | 0.012 | 3.3 |
| β-1,4-N-acetylglucosaminidase | 19 | 10.5 | 0.012 | 3.3 |
| Total nitrogen | 20 | 10.1 | 0.006 | 3.2 |
| Soil temperature | 21 | 10.1 | 0.010 | 3.2 |
| Microbial biomass carbon | 22 | 9.2 | 0.030 | 2.8 |
| Total phosphorus | 23 | 6.2 | 0.086 | 1.9 |
| Soil quality index | 24 | 6.2 | 0.088 | 1.9 |
| Available calcium | 25 | 6.0 | 0.114 | 1.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Yu, Y.; Song, Y. Soil Properties of Different Planting Combinations of Zanthoxylum planispinum var. dintanensis Plantations and Their Effect on Stoichiometry. Agronomy 2022, 12, 2562. https://doi.org/10.3390/agronomy12102562
Li Y, Yu Y, Song Y. Soil Properties of Different Planting Combinations of Zanthoxylum planispinum var. dintanensis Plantations and Their Effect on Stoichiometry. Agronomy. 2022; 12(10):2562. https://doi.org/10.3390/agronomy12102562
Chicago/Turabian StyleLi, Yitong, Yanghua Yu, and Yanping Song. 2022. "Soil Properties of Different Planting Combinations of Zanthoxylum planispinum var. dintanensis Plantations and Their Effect on Stoichiometry" Agronomy 12, no. 10: 2562. https://doi.org/10.3390/agronomy12102562
APA StyleLi, Y., Yu, Y., & Song, Y. (2022). Soil Properties of Different Planting Combinations of Zanthoxylum planispinum var. dintanensis Plantations and Their Effect on Stoichiometry. Agronomy, 12(10), 2562. https://doi.org/10.3390/agronomy12102562