

The Effect of Paraffin Oil Spraying and Powdery Mildew Infection on Leaf Gas Exchange and Yield of Chardonnay and Kékfrankos (Vitis vinifera L.) in Hungary


Abstract
:1. Introduction
2. Materials and Methods
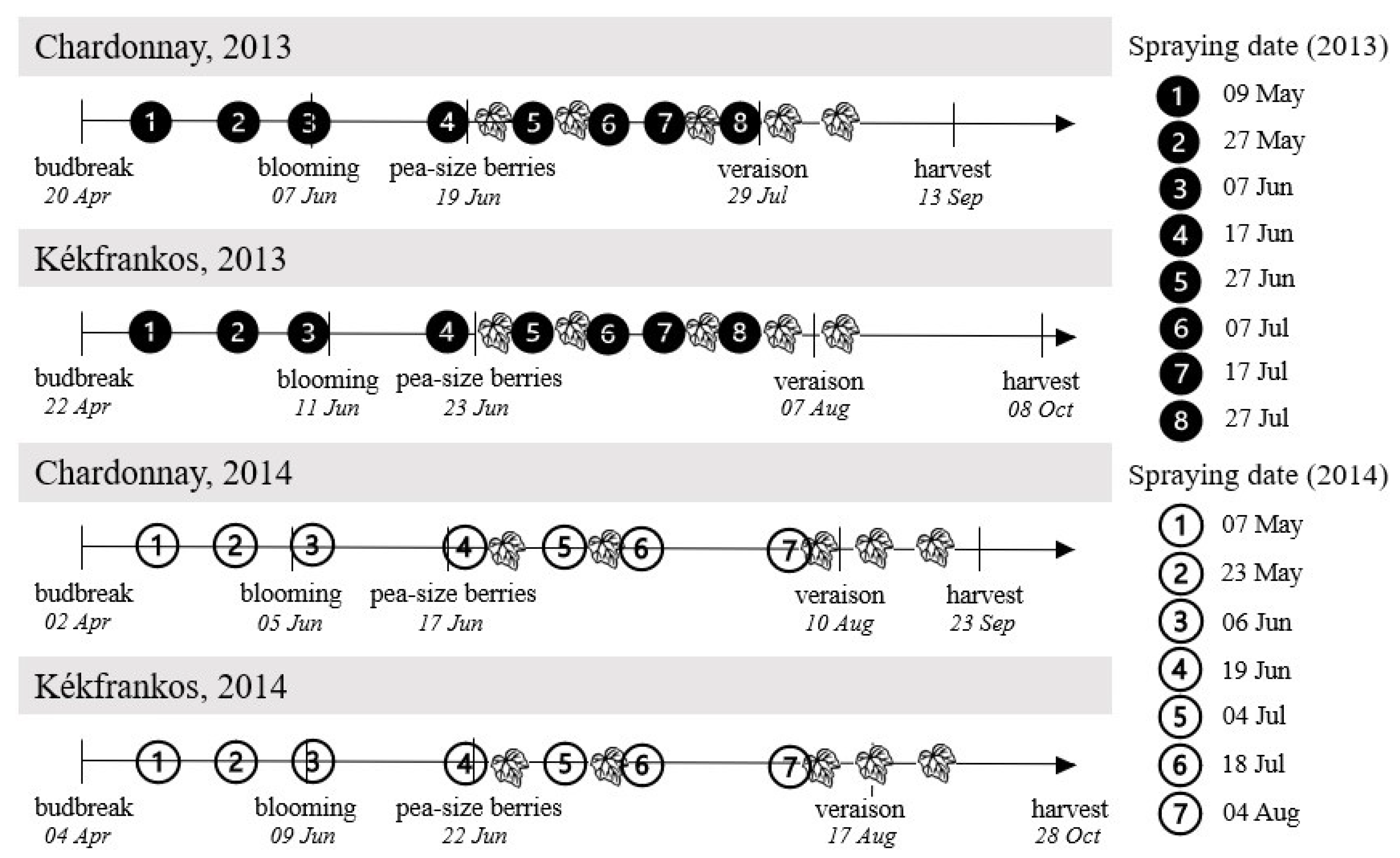
2.1. Experimental Design and Spraying
2.2. Gas Exchange Measurements, Monitoring of GPM Symptom Intensity, and Yield Parameters
2.3. Statistical Analysis
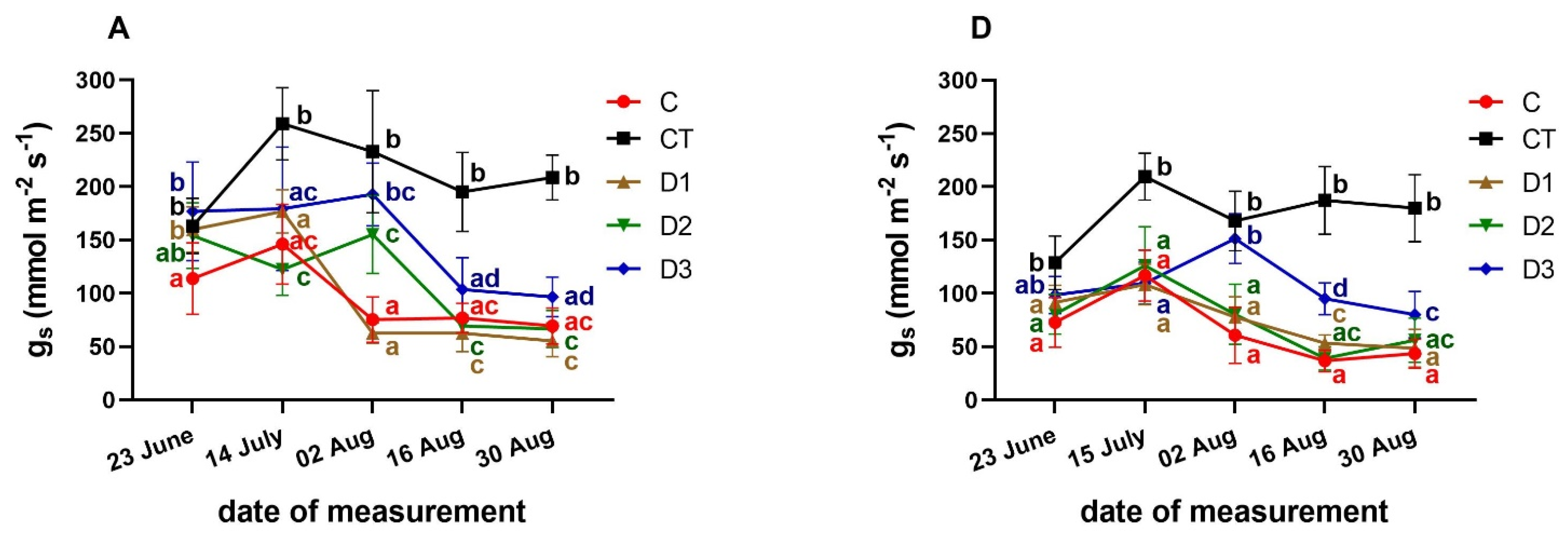
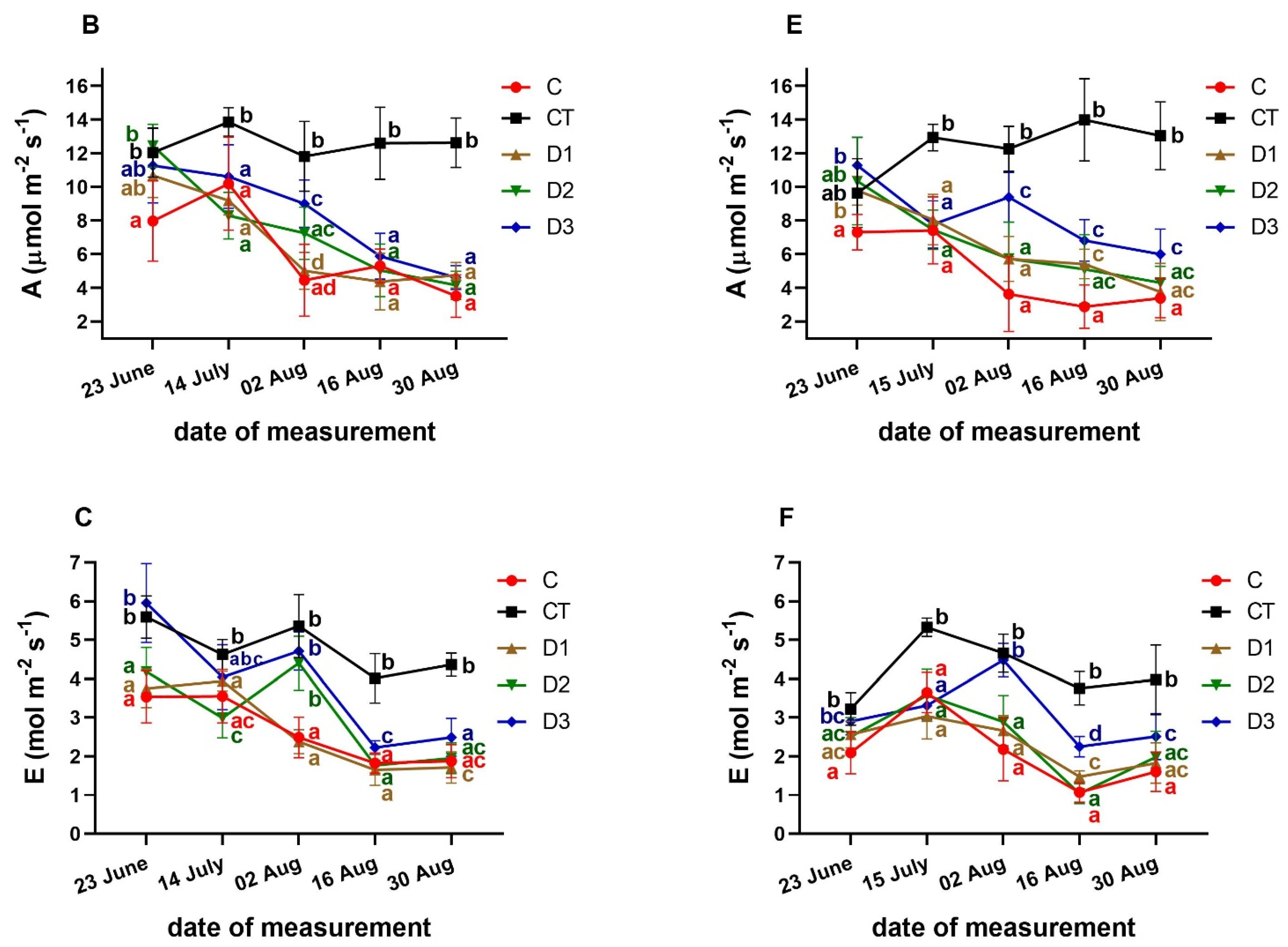
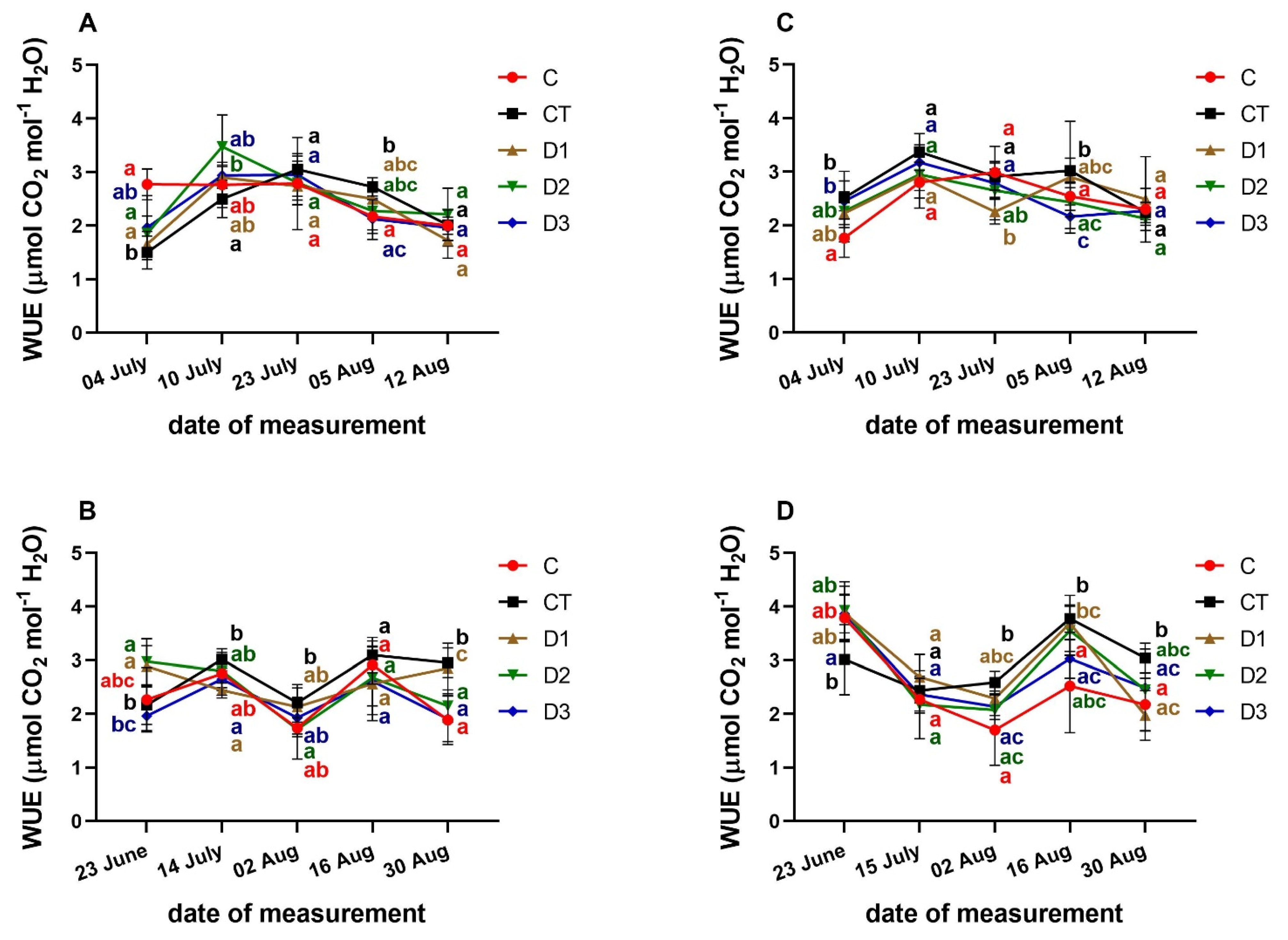
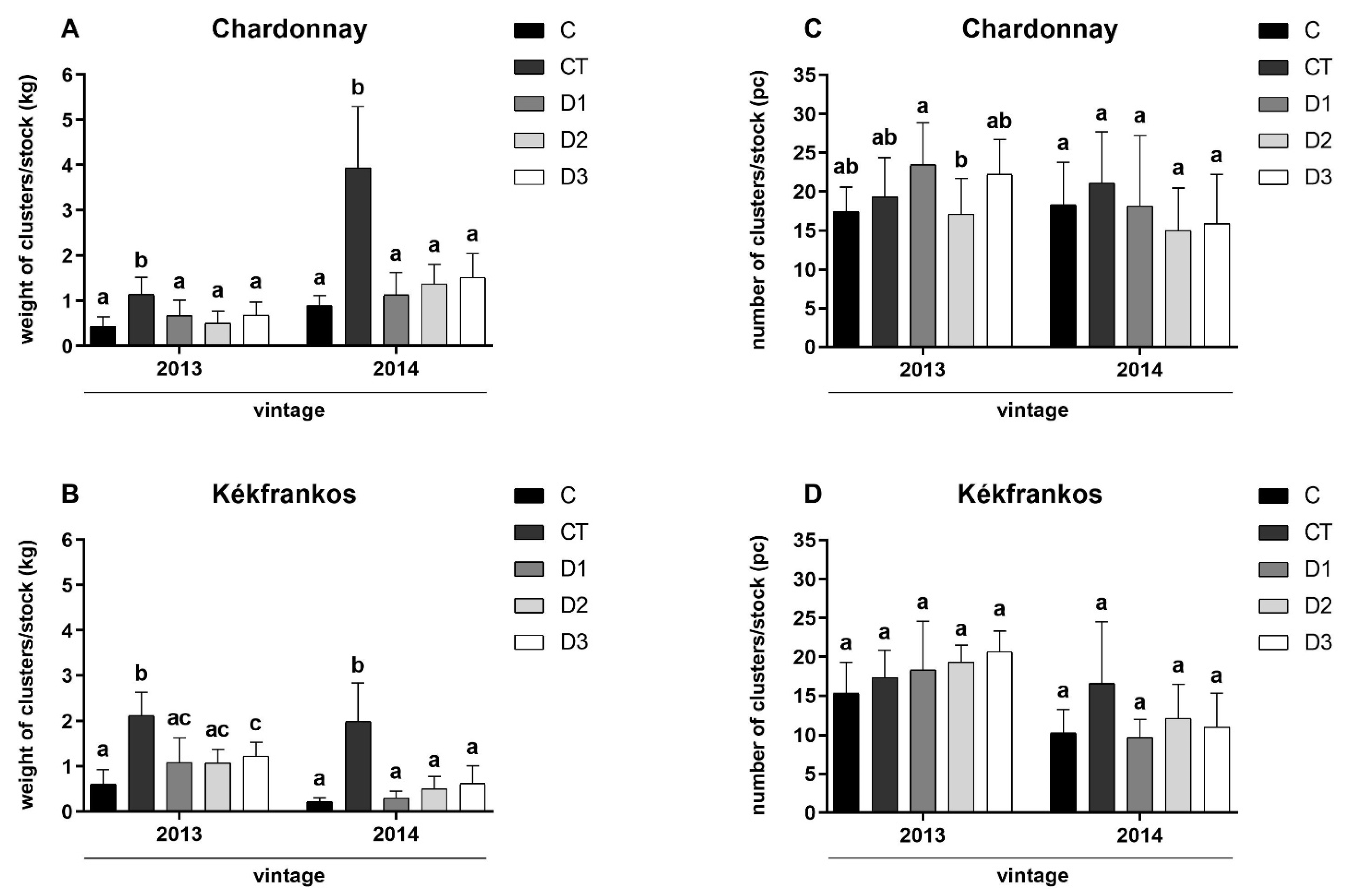
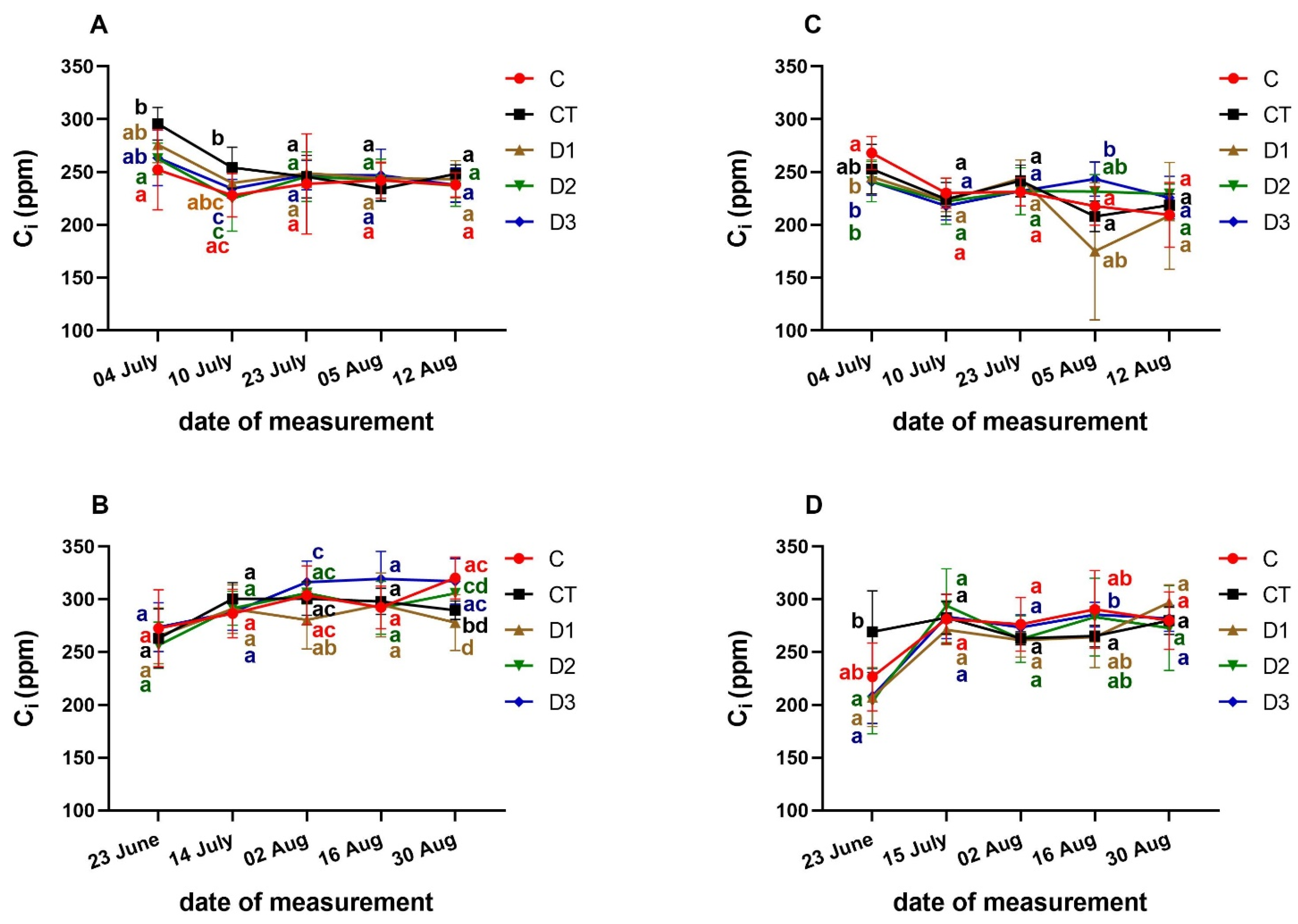
3. Results and Discussion
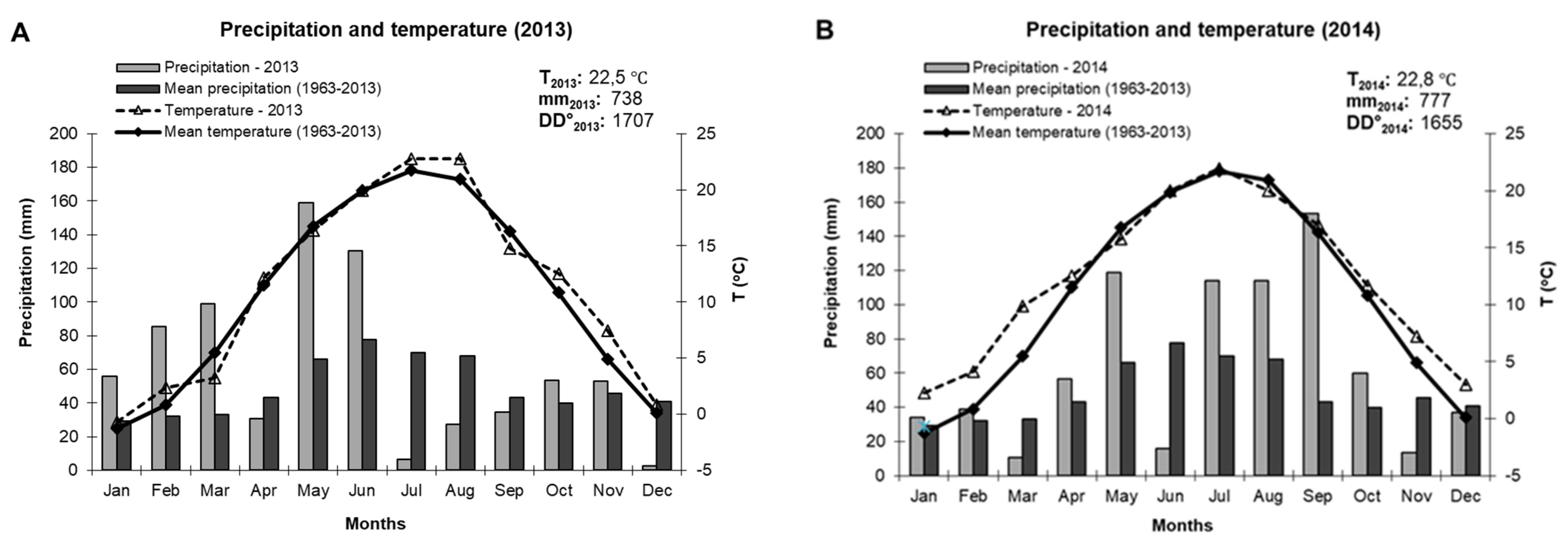
3.1. Climatic Conditions of the Experimental Years (2013–2014)
3.2. Effects of PFO Treatments on GPM Infection
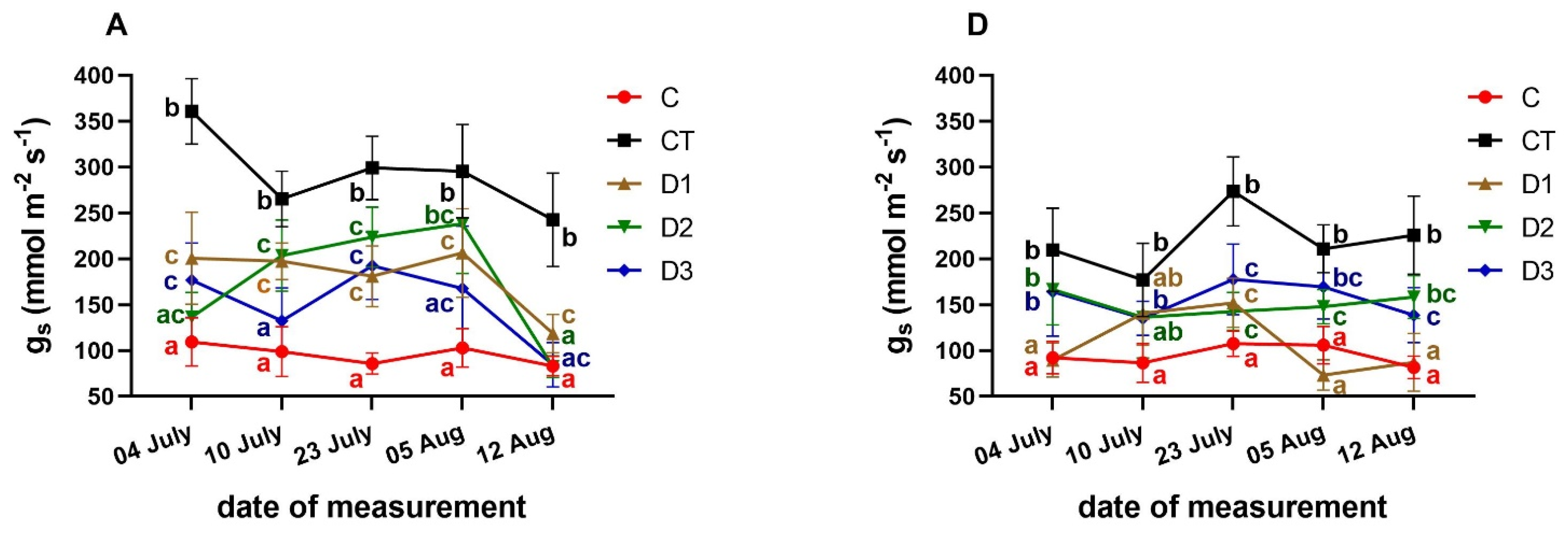
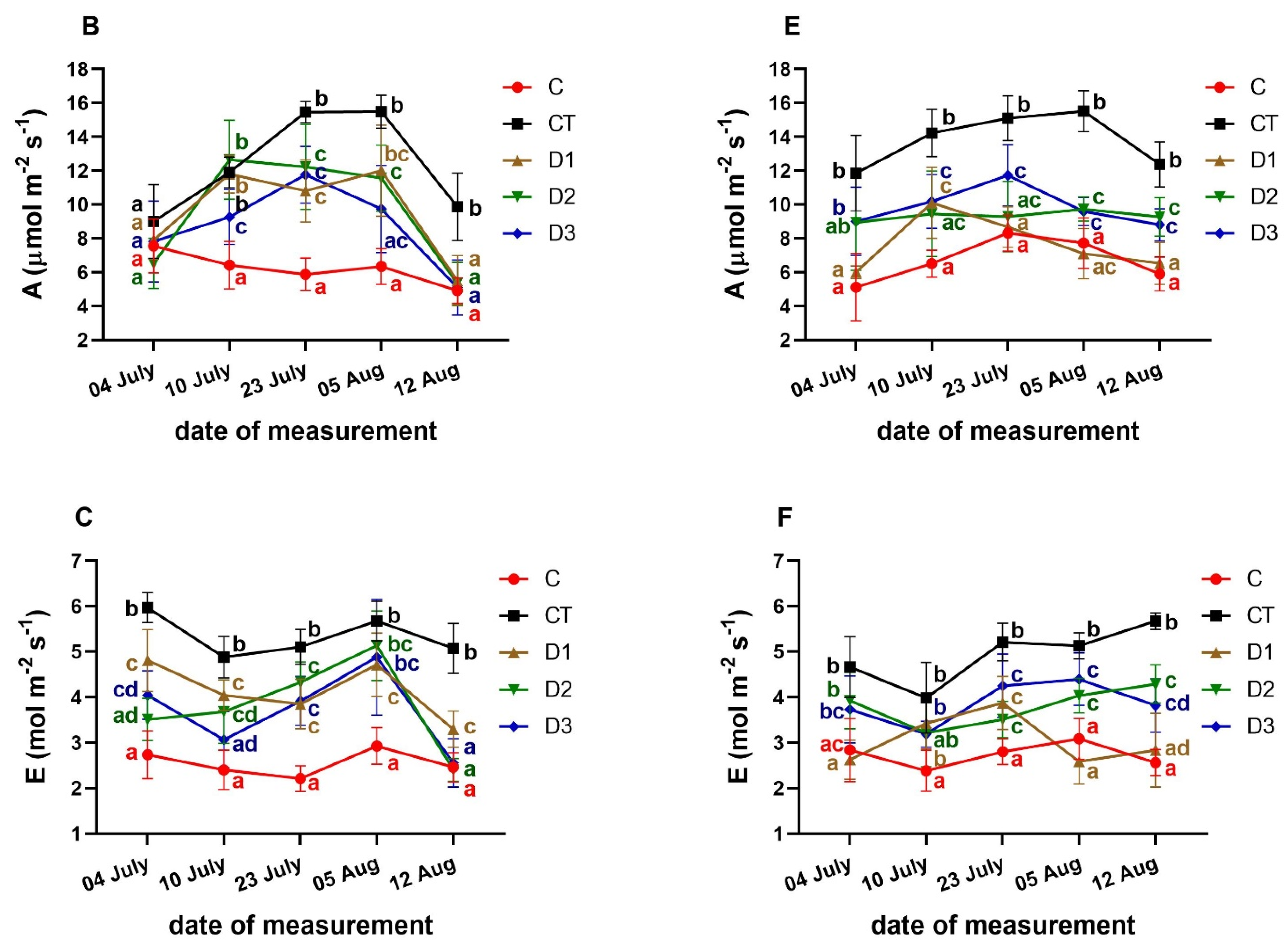
3.3. Effects of PFO Treatments on Gas Exchange Parameters
3.4. Yield Parameters of Spraying Experiment at Harvest
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2013 | 2014 |
|---|---|
| Fungicides | |
| Collis SC (boscalid, kresoxim-methyl) Falcon 460 EC (tebuconazole, triadimenol, spiroxamine) Folpan 80 WDG (folpet) Kumulus S WG (sulphur) Kocide 2000 (WG) Manzate 75 DF WG (mancozeb) Tanos 50 DP (cymoxanil, famoxadone) | Champion WG (copper hydroxide) Curzate F SC (cymoxanil, folpet) Dynali DC (cyflufenamid, difenoconazole) Falcon 460 EC (tebuconazole, triadimenol, spiroxamine) Folpan 80 WDG (folpet) Karathane Star EC (metyldinocap), Kocide 2000 (WG) Kumulus S WG (sulphur) Manzate 75 DF WG (mancozeb) Talendo EC (proquinazid) |
| Other pesticides | |
| Actara 25 WG (thiamethoxam) Pyranica 20 WP (tebufenpyrad) Pyrinex 25 CS (chlorpyrifos) Nonit SL (dioctyl sodium sulfosuccinate) | Actara 25 WG (thiamethoxam) Pyranica 20 WP (tebufenpyrad) Pyrinex 25 CS (chlorpyrifos) Nonit SL (dioctyl sodium sulfosuccinate) Spur LC (trisiloxane; modified with polyether + Pluronic L62) |
| Chardonnay 2013 | gs vs. Ci | A vs. Ci | ||||||||
| C | CT | D1 | D2 | D3 | C | CT | D1 | D2 | D3 | |
| Pearson r | 0.470 | 0.716 | 0.293 | −0.173 | 0.656 | 0.533 | −0.746 | −0.328 | −0.441 | 0.102 |
| p value | 0.425 | 0.173 | 0.632 | 0.780 | 0.229 | 0.359 | 0.148 | 0.590 | 0.457 | 0.871 |
| Significance | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| Kékfrankos 2013 | gs vs. Ci | A vs. Ci | ||||||||
| C | CT | D1 | D2 | D3 | C | CT | D1 | D2 | D3 | |
| Pearson r | 0.055 | 0.307 | 0.607 | 0.803 | 0.777 | −0.467 | −0.446 | 0.147 | −0.698 | −0.183 |
| p value | 0.930 | 0.616 | 0.277 | 0.102 | 0.122 | 0.428 | 0.452 | 0.814 | 0.190 | 0.768 |
| Significance | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| Chardonnay 2014 | gs vs. Ci | A vs. Ci | ||||||||
| C | CT | D1 | D2 | D3 | C | CT | D1 | D2 | D3 | |
| Pearson r | −0.687 | 0.817 | −0.200 | −0.427 | −0.568 | −0.783 | 0.408 | −0.469 | −0.856 | −0.841 |
| p value | 0.200 | 0.091 | 0.747 | 0.473 | 0.318 | 0.117 | 0.496 | 0.426 | 0.064 | 0.074 |
| Significance | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| Kékfrankos 2014 | gs vs. Ci | A vs. Ci | ||||||||
| C | CT | D1 | D2 | D3 | C | CT | D1 | D2 | D3 | |
| Pearson r | −0.179 | 0.468 | −0.473 | 0.074 | 0.058 | −0.618 | 0.130 | −0.841 | −0.728 | −0.858 |
| p value | 0.773 | 0.407 | 0.421 | 0.906 | 0.927 | 0.267 | 0.836 | 0.074 | 0.163 | 0.063 |
| Significance | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |

References
- Pautasso, M.; Dehnen-Schmutz, K.; Holdenrieder, O.; Pietravalle, S.; Salama, N.; Jeger, M.J.; Lange, E.; Hehl-Lange, S. Plant health and global change—Some implications for landscape management. Biol. Rev. 2010, 85, 728–755. [Google Scholar] [CrossRef]
- Rosenzweig, C.; Iglesius, A.; Yang, X.B.; Epstein, P.R.; Chivian, E. Climate change and extreme weather events—Implications for food production, plant diseases, and pests. NASA Publ. 24 Glob. Chang. Hum. Health 2001, 2, 90–104. [Google Scholar] [CrossRef]
- Özkara, A.; Akyil, D.; Konuk, M. Pesticides, environmental pollution, and health. In Environmental Health Risk—Hazardous Factors to Living Species; Larramendy, M.L., Soloneski, S., Eds.; IntechOpen: London, UK, 2011; Available online: https://www.intechopen.com/books/environmental-health-risk-hazardous-factors-to-living-species/pesticides-environmental-pollution-and-health (accessed on 13 September 2022). [CrossRef] [Green Version]
- Pertot, I.; Caffi, T.; Rossi, V.; Mugnai, L.; Hoffmann, C.; Grando, M.S.; Gary, C.; Lafond, D.; Duso, C.; Thiery, D.; et al. A critical review of plant protection tools for reducing pesticide use on grapevine and new perspectives for the implementation of IPM in viticulture. Crop Prot. 2017, 97, 70–84. [Google Scholar] [CrossRef]
- Rantsiou, K.; Giacosa, S.; Pugliese, M.; Englezos, V.; Ferrocino, I.; Río Segade, S.; Monchiero, M.; Gribaudo, I.; Gambino, G.; Gullino, M.L.; et al. Impact of chemical and alternative fungicides applied to grapevine cv Nebbiolo on microbial ecology and chemical-physical grape characteristics at harvest. Front. Plant Sci. 2020, 11, 700. [Google Scholar] [CrossRef]
- Essling, M.; McKay, S.; Petrie, P.R. Fungicide programs used to manage powdery mildew (Erysiphe necator) in Australian vineyards. Crop Prot. 2021, 139, 105369. [Google Scholar] [CrossRef]
- OIV (2019): Statistical Report on World Vitiviniculture. Available online: http://www.oiv.int/public/medias/6782/oiv-2019-statistical-report-on-world-vitiviniculture.pdf (accessed on 13 September 2022).
- Gessler, C.; Pertot, I.; Perazzolli, M. Plasmopara viticola: A review of knowledge on downy mildew of grapevine and effective disease management. Phytopathol. Mediterr. 2011, 50, 3–44. [Google Scholar]
- Sosa-Zuniga, V.; Vidal Valenzuela, A.; Barba, P.; Espinoza Cancino, C.; Romero-Romero, J.L.; Arce-Johnson, P. Powdery mildew resistance genes in vines: An opportunity to achieve a more sustainable viticulture. MDPI Pathog. 2022, 11, 703. [Google Scholar] [CrossRef]
- Gadoury, D.M.; Cadle-Davidson, L.; Wilcox, W.F.; Dry, I.B.; Seem, R.C.; Milgroom, M.G. Grapevine powdery mildew (Erysiphe necator): A fascinating system for the study of the biology, ecology and epidemiology of an obligate biotroph. Mol. Plant Pathol. 2012, 13, 1–16. [Google Scholar] [CrossRef]
- Malviya, D.; Thosar, R.; Kokare, N.; Pawar, S.; Singh, U.B.; Saha, S.; Rai, J.P.; Singh, H.V.; Somkuwar, R.G.; Saxena, A.K. A comparative analysis of microbe-based technologies developed at ICAR-NBAIM against Erysiphe necator causing powdery mildew disease in grapes (Vitis vinifera L.). Front Microbiol. 2022, 13, 871901. [Google Scholar] [CrossRef]
- Nail, W.R.; Howell, G.S. Effects of powdery mildew of grape on carbon assimilation mechanisms of potted ‘Chardonnay’ Grapevines. HortScience 2004, 39, 1670–1673. [Google Scholar] [CrossRef]
- Moriondo, M.; Orlandini, S.; Giuntoli, A.; Bindi, M. The effect of downy and powdery mildew on grapevine (Vitis vinifera L.) leaf gas exchange. J. Phytopathol. 2005, 153, 350–357. [Google Scholar] [CrossRef]
- Liu, E.; Gold, K.M.; Combs, D.; Cadle-Davidson, L.; Jiang, Y. Deep semantic segmentation for the quantification of grape foliar diseases in the vineyard. Front. Plant Sci. 2022, 13, 978761. [Google Scholar] [CrossRef]
- Vielba-Fernández, A.; Polonio, Á.; Ruiz-Jiménez, L.; de Vicente, A.; Pérez-García, A.; Fernández-Ortuño, D. Fungicide resistance in powdery mildew fungi. MDPI Microorg. 2020, 8, 1431. [Google Scholar] [CrossRef] [PubMed]
- Massi, F.; Torriani, S.F.F.; Borghi, L.; Toffolatti, S.L. Fungicide resistance evolution and detection in plant pathogens: Plasmopara viticola as a case study. MDPI Microorg. 2021, 9, 119. [Google Scholar] [CrossRef] [PubMed]
- Yildirim, İ.; Dardeniz, A. Effects of alternative spray programs and various combinations of green pruning on powdery mildew [Uncinula necator (Schw.) Burr.] in Karasakız (Kuntra) grape cultivar. Turk. J. Agric. For. 2010, 3, 213–223. [Google Scholar] [CrossRef]
- Romanazzi, G.; Mancini, V.; Feliziani, E.; Servili, A.; Endeshaw, S.; Neri, D. Impact of alternative fungicides on grape downy mildew control and vine growth and development. Plant Dis. 2016, 100, 739–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebbon, G.P. Environmental and health aspects of agricultural spray oils. In Spray Oils Beyond 2000; Beattie, G.A.C., Watson, D.M., Stevens, M.L., Rae, D.J., Spooner-Hart, R.N., Eds.; University of Western Sydney: Penrith, Australia, 2002; pp. 232–246. [Google Scholar]
- Nile, A.S.; Kwon, Y.D.; Nile, S.H. Horticultural oils: Possible alternatives to chemical pesticides and insecticides. Environ. Sci. Pollut. Res. 2019, 26, 21127–21139. [Google Scholar] [CrossRef] [PubMed]
- Eichmann, R.; Hückelhoven, R. Accommodation of powdery mildew fungi in intact plant cells. J. Plant Physiol. 2008, 165, 5–18. [Google Scholar] [CrossRef]
- Dell, K.J.; Gubler, W.D.; Krueger, R.; Sanger, M.; Bettiga, L.J. The efficacy of JMS Stylet-Oil on grape powdery mildew and bunch rot and effects on fermentation. Am. J. Enol. Vitic. 1998, 49, 11–16. [Google Scholar]
- Rae, D.J. Use of spray oils with synthetic insecticides, acaricides and fungicides. In Spray Oils Beyond 2000; Beattie, G.A.C., Watson, D.M., Stevens, M.L., Rae, D.J., Spooner-Hart, R.N., Eds.; University of Western Sydney: Penrith, Australia, 2002; pp. 248–266. [Google Scholar]
- Grove, G.G.; Lunden, J.; Spayd, S. Use of Petroleum Delivered Spray Oils in Washington Grapevine Powdery Mildew Management Programs; Online: Plant Health Progress; WSU, Department of Plant Pathology: Pullman, WA, USA, 2005. [Google Scholar] [CrossRef] [Green Version]
- Janousek, C.N.; Bay, I.S.; Gubler, W.D. Control of Grape Powdery Mildew with Synthetic, Biological, and Organic Fungicides: 2009 Filed Trials; UC Davis, Department of Plant Pathology: Davis, CA, USA; Available online: https://escholarship.org/uc/item/8fz3p4vc (accessed on 13 September 2022).
- Northover, J.; Schneider, K.E. Physical modes of action of petroleum and plant oils on powdery and downy mildews of grapevines. Plant Dis. 1996, 77, 152–157. [Google Scholar] [CrossRef]
- Matese, A.; Di Gennaro, S.F. Technology in precision viticulture: A state of the art review. Int. J. Wine Res. 2015, 7, 69–81. [Google Scholar] [CrossRef] [Green Version]
- Doster, M.A.; Schnathorst, W.C. Compare susceptibility of various grapevine cultivars to the powdery mildew fungus Uncinula necator. Am. J. Enol. Vitic. 1985, 36, 101–104. [Google Scholar]
- Gaforio, L.; García-Muñoz, S.; Cabello, F.; Muñoz-Organero, G. Evaluation of susceptibility to powdery mildew (Erysiphe necator) in Vitis vinifera varieties. Vitis 2011, 50, 123–126. [Google Scholar]
- Gadoury, D.M.; Seem, R.C.; Ficke, A.; Wilcox, W.F. Ontogenic resistance to powdery mildew in grape berries. Phytopathology 2003, 93, 547–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, M.C. Phytotoxicity: An overview of the physiological responses of plants exposed to fungicides. J. Bot. 2012, 135479. [Google Scholar] [CrossRef] [Green Version]
- Saladin, G.; Magné, C.; Clément, C. Effects of fludioxonil and pyrimethanil, two fungicides used against Botrytis cinerea, on carbohydrate physiology in Vitis vinifera L. Pest Manag. Sci. 2003, 59, 1083–1092. [Google Scholar] [CrossRef]
- Petit, A.N.; Fontaine, F.; Clément, C.; Vaillant-Gaveau, N. Photosynthesis limitations of grapevine after treatment with the fungicide fludioxonil. J. Agric. Food Chem. 2008, 56, 6761–6767. [Google Scholar] [CrossRef]
- Hodgkinson, M.C.; Johnson, D.; Smith, G. Causes of phytotoxicity induced by petroleum-derived spray oils. In Spray Oils Beyond 2000; Beattie, G.A.C., Watson, D.M., Stevens, M.L., Rae, D.J., Spooner-Hart, R.N., Eds.; University of Western Sydney: Penrith, Australia, 2002; pp. 170–178. [Google Scholar]
- Finger, S.A.; Wolf, T.K.; Baudoin, A.B. Effects of horticultural oils on the photosynthesis, fruit maturity, and crop yield of winegrapes. Am. J. Enol. Vitic. 2002, 53, 116–124. [Google Scholar]
- Ferrini, F.; Mattii, G.B.; Nicese, F.P. The effect of temperature on key physiological responses of grapevine leaf. Am. J. Enol. Vitic. 1995, 46, 375–379. [Google Scholar]
- Nazari, M.; Dada, A.; Asgharzadeh, A. Effects of spray volck oil in different times on the cluster characters of grape (Kolahdari var) in North-Khorasan Condition. JLS 2014, 4, 2231–2345. [Google Scholar]
- Wicks, T.J.; Hitch, C.; Campbell, K.; Hall, B. Control of grapevine powdery mildew with mineral oil: An assessment of oil concentration and spray volume. Aust. J. Grape Wine Res. 1999, 5, 61–65. [Google Scholar] [CrossRef]
- Martín, B.; Hernández, S.; Silvarrey, C.; Jacay, J.A.; Cabaleiro, C. Vegetable, fish and mineral oils control grapevine powdery mildew. Phytopathol. Mediterr. 2005, 44, 169–179. [Google Scholar]
- Viala, P.; Vermorel, V. Tome IV (Chardonnay). In Traité Général de Viticulture, Ampélographie (1901–1909); Masson et cie: Paris, France, 1903; pp. 5–23. [Google Scholar]
- Bényei, F. Államilag elismert fehérborszőlő-fajták. In Borszőlőfajták, Csemegeszőlő-Fajták és Alanyok; Bényei, F., Lőrincz, A., Eds.; Mezőgazda Kiadó: Budapest, Hungary, 2005; pp. 107–108,165. [Google Scholar]
- Viala, P.; Vermorel, V. Tome III (Limberger). In Traité Général de Viticulture, Ampélographie (1901–1909); Masson et cie: Paris, France, 1902; pp. 203–209. [Google Scholar]
- Hajdu, E. Magyar Szőlőfajták/Varieties of Hungarian Grapes; Mezőgazda Kiadó: Budapest, Hungary, 2003; p. 80. [Google Scholar]
- Lőrincz, A. Államilag elismert vörösborszőlő-fajták (Kékfrankos). In Borszőlőfajták, Csemegeszőlő-Fajták és Alanyok; Bényei, F., Lőrincz, A., Eds.; Mezőgazda Kiadó: Budapest, Hungary, 2005; pp. 165–166. [Google Scholar]
- Candolfi-Vasconcelos, M.C.; Castagnoli, S. Plant Growth, Leaf Gas-Exchange, Fruit Yield and Quality as Affected by Green Covers; Technical Report; OSU, Oregon Wine Research Institute: Corvallis, OR, USA, 1996; Available online: https://ir.library.oregonstate.edu/concern/technical_reports/n009w345b (accessed on 13 September 2022).
- Baudoin, A.; Finger, S.M.; Wolf, T.K. Factors affecting reductions in photosynthesis caused by applying horticultural oil to grapevine leaves. HortScience 2006, 41, 346–351. [Google Scholar] [CrossRef] [Green Version]
- Shtienberg, D. Effects of foliar diseases to gas exchange processes: A comparative study. Phytopath 1992, 82, 760–765. [Google Scholar] [CrossRef]
- Medrano, H.; Tomás, M.; Martorell, S.; Flexas, J.; Hernández, E.; Rosselló, J.; Pou, A.; Escalona, J.M.; Bota, J. From leaf to whole-plant water use efficiency (WUE) in complex canopies: Limitations of leaf WUE as a selection target. Crop J. 2015, 3, 220–228. [Google Scholar] [CrossRef] [Green Version]
- Wicks, T.J.; Hitch, C.J. Integration of strobilurins and other fungicides for the control of powdery mildew on grapes. Aust. J. Grape Wine Res. 2002, 8, 132–139. [Google Scholar] [CrossRef]
- Pálfi, X.; Bisztray, G.D.; Villangó, S.; Pálfi, Z.; Deák, T.; Karácsony, Z.; Cseke, G.; Nagy, P.T.; Zsófi, Z. Paraffinolaj hatékonyságának tesztelése szőlőlisztharmat ellen az Egri Borvidéken. Acta Agrar. Debr. 2016, 68, 73–80. [Google Scholar] [CrossRef]
- Keller, M. The Science of Grapevines: Anatomy and Physiology, 1st ed.; Chapter 2.3: Reproductive Cycle; Academic Press: Cambridge, MA, USA; Elsevier Inc.: Amsterdam, The Netherlands, 2010; pp. 69–77. ISBN 978-0-12-374881-2. [Google Scholar]
- Winkel, T.; Rambal, S. Stomatal conductance of some grapevines growing in the field under a Mediterranean environment. Agric. For. Metemol. 1990, 51, 107–121. [Google Scholar] [CrossRef]
- Escalona, J.M.; Flexas, J.; Medrano, H. Stomatal and non-stomatal limitations of photosynthesis under water stress in field-grown grapevines. Aust. J. Plant Physiol. 1999, 26, 421–433. [Google Scholar] [CrossRef]
- Mullins, M.G.; Bauquet, A.; Williams, L.E. Biology of the Grapevine; Chapter 4: Developmental Physiology: The Vegetative Grapevine; Cambridge University Press: Cambridge, UK, 1995; p. 83. [Google Scholar]
- Pálfi, X.; Lovas, M.; Zsófi, Z.; Kátai, J.; Karácsony, Z.; Váczy, K.Z. Paraffin oil induces resistance against powdery mildew in grapevine through salicylic acid signaling. Pest Manag. Sci. 2021, 77, 6492. [Google Scholar] [CrossRef]
- Gudin, C.; Syratt, W.J.; Boize, L. The mechanism of photosynthetic inhibition and the development of scorch in tomato plants treated with spray oils. Ann. App. Biol. 1976, 84, 213–219. [Google Scholar] [CrossRef]
- Guidi, L.; Nali, C.; Ciompi, S.; Lorenzini, G.; Soldatini, G.F. Effects of two wilt fungi on leaf gas exchange of asymptomatic leaves of tomato plants. Adv. Hortic. Sci. 1997, 11, 123–127. [Google Scholar]
- Lakso, A.N.; Pratt, C.; Pearson, R.C.; Pool, R.M.; Seem, R.C.; Welser, M.J. Photosynthesis, transpiration, and water-use efficiency of mature grape leaves infected with Uncinula necator (powdery mildew). Phytopathology 1982, 72, 232–236. [Google Scholar] [CrossRef]








| Chardonnay 2013 | gs vs. A | gs vs. E | ||||||||
| C | CT | D1 | D2 | D3 | C | CT | D1 | D2 | D3 | |
| Pearson r | 0.921 | −0.114 | 0.787 | 0.940 | 0.821 | 0.730 | 0.833 | 0.870 | 0.930 | 0.806 |
| p value | 0.026 | 0.854 | 0.114 | 0.017 | 0.088 | 0.161 | 0.079 | 0.055 | 0.020 | 0.099 |
| Significance | * | ns | ns | * | ns | ns | ns | ns | * | ns |
| Kékfrankos 2013 | gs vs. A | gs vs. E | ||||||||
| C | CT | D1 | D2 | D3 | C | CT | D1 | D2 | D3 | |
| Pearson r | 0.793 | 0.208 | 0.834 | −0.651 | 0.467 | 0.775 | 0.677 | 0.970 | 0.761 | 0.807 |
| p value | 0.109 | 0.737 | 0.079 | 0.234 | 0.428 | 0.123 | 0.209 | 0.006 | 0.135 | 0.090 |
| Significance | ns | ns | ns | ns | ns | ns | ns | ** | ns | ns |
| Chardonnay 2014 | gs vs. A | gs vs. E | ||||||||
| C | CT | D1 | D2 | D3 | C | CT | D1 | D2 | D3 | |
| Pearson r | 0.987 | 0.606 | 0.955 | 0.815 | 0.909 | 0.899 | −0.210 | 0.969 | 0.985 | 0.865 |
| p value | 0.001 | 0.278 | 0.011 | 0.092 | 0.032 | 0.038 | 0.734 | 0.006 | 0.002 | 0.058 |
| Significance | ** | ns | * | ns | * | * | ns | ** | ** | ns |
| Kékfrankos 2014 | gs vs. A | gs vs. E | ||||||||
| C | CT | D1 | D2 | D3 | C | CT | D1 | D2 | D3 | |
| Pearson r | 0.859 | 0.864 | 0.833 | 0.498 | 0.442 | 0.979 | 0.760 | 0.929 | 0.957 | 0.948 |
| p value | 0.062 | 0.058 | 0.080 | 0.393 | 0.455 | 0.003 | 0.135 | 0.022 | 0.010 | 0.014 |
| Significance | ns | ns | ns | ns | ns | ** | ns | * | * | * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pálfi, X.; Villangó, S.; Karácsony, Z.; Kátai, J.; Zsófi, Z. The Effect of Paraffin Oil Spraying and Powdery Mildew Infection on Leaf Gas Exchange and Yield of Chardonnay and Kékfrankos (Vitis vinifera L.) in Hungary. Agronomy 2022, 12, 2684. https://doi.org/10.3390/agronomy12112684
Pálfi X, Villangó S, Karácsony Z, Kátai J, Zsófi Z. The Effect of Paraffin Oil Spraying and Powdery Mildew Infection on Leaf Gas Exchange and Yield of Chardonnay and Kékfrankos (Vitis vinifera L.) in Hungary. Agronomy. 2022; 12(11):2684. https://doi.org/10.3390/agronomy12112684
Chicago/Turabian StylePálfi, Xénia, Szabolcs Villangó, Zoltán Karácsony, János Kátai, and Zsolt Zsófi. 2022. "The Effect of Paraffin Oil Spraying and Powdery Mildew Infection on Leaf Gas Exchange and Yield of Chardonnay and Kékfrankos (Vitis vinifera L.) in Hungary" Agronomy 12, no. 11: 2684. https://doi.org/10.3390/agronomy12112684
APA StylePálfi, X., Villangó, S., Karácsony, Z., Kátai, J., & Zsófi, Z. (2022). The Effect of Paraffin Oil Spraying and Powdery Mildew Infection on Leaf Gas Exchange and Yield of Chardonnay and Kékfrankos (Vitis vinifera L.) in Hungary. Agronomy, 12(11), 2684. https://doi.org/10.3390/agronomy12112684