
Application of PGPB Combined with Variable N Doses Affects Growth, Yield-Related Traits, N-Fertilizer Efficiency and Nutritional Status of Lettuce Grown under Controlled Condition

 ,
, 
 ,
,  ,
,  ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site and Material
2.2. Design and Procedure
2.3. Measurements
2.4. Statistical Analyses
3. Results
3.1. Implications of PGPB and Nitrogen Dose on Yield and Yield-Related Features
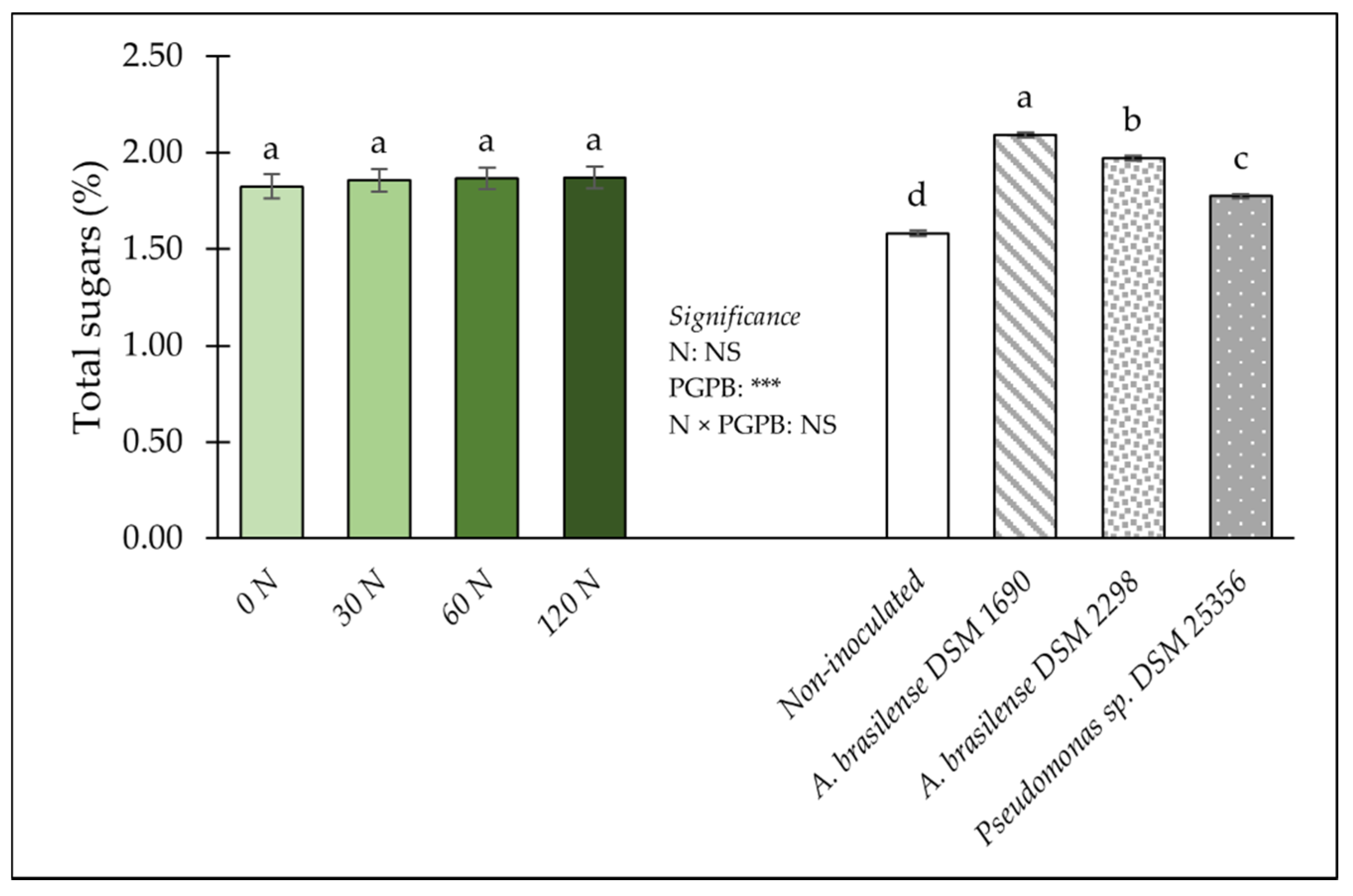
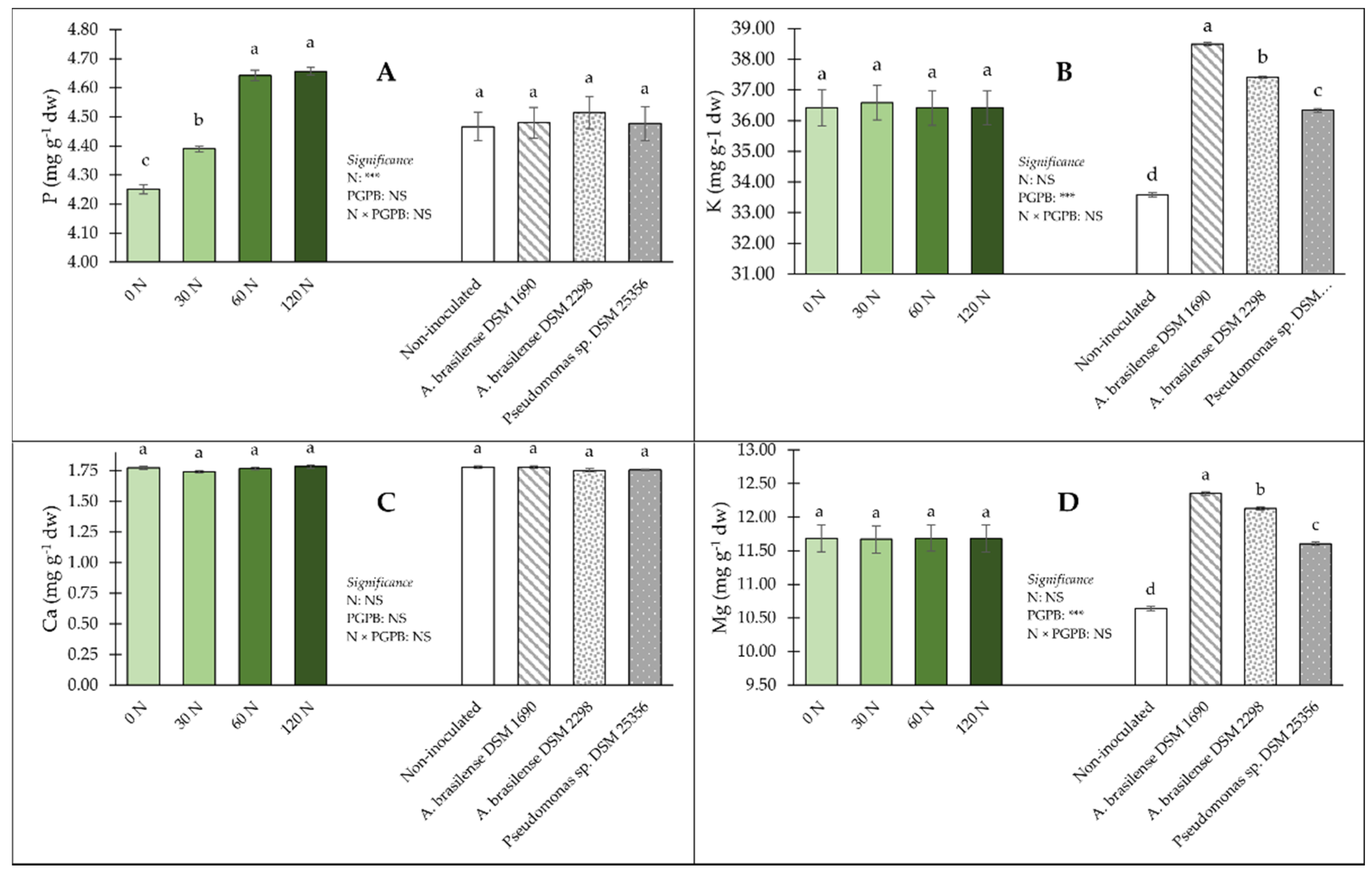
3.2. Implications of PGPB and Nitrogen Dose on Leaf Colour, Nutritional and Functional Components and Mineral Concentrations
3.3. Nitrogen Indices
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mou, B. Lettuce. In Vegetables I; Prohens, J., Nuez, F., Eds.; Springer: New York, NY, USA, 2008; pp. 75–116. [Google Scholar]
- Sabatino, L.; Consentino, B.B.; Rouphael, Y.; De Pasquale, C.; Iapichino, G.; D’Anna, F.; La Bella, S. Protein Hydrolysates and Mo-Biofortification Interactively Modulate Plant Performance and Quality of ‘Canasta’ Lettuce Grown in a Protected Environment. Agronomy 2021, 11, 1023. [Google Scholar] [CrossRef]
- FAOSTAT. Available online: https://www.fao.org/faostat/en/#data (accessed on 17 November 2021).
- Kim, M.J.; Moon, Y.; Tou, J.C.; Mou, B.; Waterland, N.L. Nutritional value, bioactive compounds and health benefits of lettuce (Lactuca sativa L.). J. Food Compos. Anal. 2016, 49, 19–34. [Google Scholar] [CrossRef]
- Ciriello, M.; Formisano, L.; Pannico, A.; El-Nakhel, C.; Fascella, G.; Duri, L.G.; Cristofano, F.; Gentile, B.R.; Giordano, M.; Rouphael, Y.; et al. Nutrient Solution Deprivation as a Tool to Improve Hydroponics Sustainability: Yield, Physiological, and Qualitative Response of Lettuce. Agronomy 2021, 11, 1469. [Google Scholar] [CrossRef]
- El-Nakhel, C.; Pannico, A.; Kyriacou, M.C.; Giordano, M.; De Pascale, S.; Rouphael, Y. Macronutrient deprivation eustress elicits differential secondary metabolites in red and green-pigmented butterhead lettuce grown in a closed soilless system. J. Sci. Food Agric. 2019, 99, 6962–6972. [Google Scholar] [CrossRef]
- Li, Z.; Zhao, X.; Sandhu, A.K.; Gu, L. Effects of exogenous abscisic acid on yield, antioxidant capacities, and phytochemical contents of greenhouse grown lettuces. J. Agric. Food Chem. 2010, 58, 6503–6509. [Google Scholar] [CrossRef]
- Luz, G.L.; Medeiros, S.L.P.; Manfron, P.A.; Amaral, A.D.; Muller, L.; Torrez, M.G.; Mentges, L.A. The nitrate issue in hydroponic lettuce and the human health. Ciência Rural 2008, 38, 2388–2394. [Google Scholar] [CrossRef] [Green Version]
- Romani, A.; Pinelli, P.; Galardi, C.; Sani, G.; Cimato, A.; Heimler, D. Polyphenols in greenhouse and open-air-grown lettuce. Food Chem. 2002, 79, 337–342. [Google Scholar] [CrossRef] [Green Version]
- Miceli, A.; Vetrano, F.; Sabatino, L.; D’Anna, F.; Moncada, A. Influence of Preharvest Gibberellic Acid Treatments on Postharvest Quality of Minimally Processed Leaf Lettuce and Rocket. Horticulturae 2019, 5, 63. [Google Scholar] [CrossRef] [Green Version]
- Sabatino, L.; La Bella, S.; Ntatsi, G.; Iapichino, G.; D’Anna, F.; De Pasquale, C.; Consentino, B.B.; Rouphael, Y. Selenium biofortification and grafting modulate plant performance and functional features of cherry tomato grown in a soilless system. Sci. Hortic. 2021, 285, 110095. [Google Scholar] [CrossRef]
- Mauro, R.P.; Agnello, M.; Distefano, M.; Sabatino, L.; San Bautista Primo, A.; Leonardi, C.; Giuffrida, F. Chlorophyll Fluorescence, Photosynthesis and Growth of Tomato Plants as Affected by Long-Term Oxygen Root Zone Deprivation and Grafting. Agronomy 2020, 10, 137. [Google Scholar] [CrossRef] [Green Version]
- Solaiman, A.R.M.; Rahbbani, M.G. Effect of NPKS and cow dung on growth and yield of tomato. Bull. Inst. Trop. Agric. Kyushu Univ. 2006, 29, 31–37. [Google Scholar]
- Zaidi, A.; Ahmad, E.; Khan, M.S.; Saif, S.; Rizvi, A. Role of Plant Growth Promoting Rhizobacteria in Sustainable Production of Vegetables: Current Perspective. Sci. Hortic. 2015, 193, 231–239. [Google Scholar] [CrossRef]
- Sabatino, L.; Iapichino, G.; La Bella, S.; Tuttolomondo, T.; D’Anna, F.; Cardarelli, M.; Consentino, B.B.; Rouphael, Y. An Appraisal of Calcium Cyanamide as Alternative N Source for Spring-Summer and Fall Season Curly Endive Crops: Effects on Crop Performance, NUE and Functional Quality Components. Agronomy 2020, 10, 1357. [Google Scholar] [CrossRef]
- Sabatino, L.; Iapichino, G.; Consentino, B.B.; D’Anna, F.; Rouphael, Y. Rootstock and Arbuscular Mycorrhiza Combinatorial Effects on Eggplant Crop Performance and Fruit Quality under Greenhouse Conditions. Agronomy 2020, 10, 693. [Google Scholar] [CrossRef]
- Consentino, B.B.; Virga, G.; La Placa, G.G.; Sabatino, L.; Rouphael, Y.; Ntatsi, G.; Iapichino, G.; La Bella, S.; Mauro, R.P.; D’Anna, F.; et al. Celery (Apium graveolens L.) Performances as Subjected to Different Sources of Protein Hydrolysates. Plants 2020, 9, 1633. [Google Scholar] [CrossRef]
- La Bella, S.; Consentino, B.B.; Rouphael, Y.; Ntatsi, G.; De Pasquale, C.; Iapichino, G.; Sabatino, L. Impact of Ecklonia Maxima Seaweed Extract and Mo Foliar Treatments on Biofortification, Spinach Yield, Quality and NUE. Plants 2021, 10, 1139. [Google Scholar] [CrossRef] [PubMed]
- Davison, J. Plant Beneficial Bacteria. Nat. Biotechnol. 1988, 6, 282–286. [Google Scholar] [CrossRef]
- Kloepper, J.W.; Lifshitz, R.; Zablotowicz, R.M. Free-Living Bacterial Inocula for Enhancing Crop Productivity. Trends Biotechnol. 1989, 7, 39–44. [Google Scholar] [CrossRef]
- Cassan, F.; García de Salamone, I. Azospirillum: Cell Physiology, Plant Response, Agronomic and Environmental Research in Argentina; Asociación Argentina de Microbiologia: Dean Funes, Argentina, 2008; ISBN 978-987-98475-8-9. [Google Scholar]
- Tien, T.M.; Gaskins, M.H.; Hubbell, D.H. Plant growth substances produced by Azospirillum brasilense and their effect on the growth of pearl millet (Pennisetum Americanum L.). Appl. Environ. Microbiol. 1979, 37, 1016–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bottini, R.; Fulchieri, M.; Pearce, D.; Pharis, R. Identification of gibberelins A1, A3, and iso-A3 in cultures of A. lipoferum. Plant Physiol. 1989, 90, 45–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strzelczyk, E.; Kamper, M.; Li, C. Cytocinin-like-substances and ethylene production by Azospirillum in media with different carbon sources. Microbiol. Res. 1994, 149, 55–60. [Google Scholar] [CrossRef]
- Rodriguez, H.; Gonzalez, T.; Goire, I.; Bashan, Y. Gluconic Acid Production and Phosphate Solubilization by the Plant Growth-Promoting Bacterium Azospirillum spp. Naturwissenschaften 2004, 91, 552–555. [Google Scholar] [CrossRef] [PubMed]
- Correa, O.S.; Romero, A.M.; Soria, M.A.; de Estrada, M. Azospirillum brasilense-plant genotype interactions modify tomato response to bacterial diseases, and root and foliar microbial communities. In Azospirillum sp.: Cell Physiology, Plant Interactions and Agronomic Research in Argentina; Cassán, F.D., Garcia de Salamone, I., Eds.; Asociación Argentina de Microbiologia: Dean Funes, Argentina, 2008; Volume 266, pp. 85–94. [Google Scholar]
- Weller, D.M.; Thomashow, L.S. Current Challenges in Introducing Beneficial Microorganisms into the Rhizosphere. In Molecular Ecology of Rhizosphere Microorganisms; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 1994; pp. 1–18. ISBN 978-3-527-61581-0. [Google Scholar]
- Glick, B.R. The enhancement of plant growth by freeliving bacteria. Can. J. Microbiol. 1995, 41, 109–117. [Google Scholar] [CrossRef]
- Probanza, A.; Lucas, J.A.; Acero, N.; Gutierrez Mañero, F.J. The Influence of Native Rhizobacteria on European Alder (Alnus Glutinosa L. Gaertn.) Growth. Plant Soil 1996, 182, 59–66. [Google Scholar] [CrossRef]
- Dobereiner, J.; Pedrosa, F.O. Nitrogen-Fixing Bacteria in Nonleguminous Crop Plants; Brock/Springer Series in Contemporary Bioscience; Science Tech Publishers: Hampshire, UK, 1987; pp. 1–155. [Google Scholar]
- Broadley, M.R.; Escobar-Gutièrrez, A.J.; Burns, A.; Burns, G. What are the effects of nitrogen deficiency on growth components of lettuce? New Phytol. 2000, 147, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Tesi, R. Orticoltura Mediterranea Sostenibile; Pàtron Editore: Bologna, Italy, 2010. [Google Scholar]
- Meda, A.; Lamien, C.E.; Romito, M.; Millogo, J.; Nacoulma, O.G. Determination of the total phenolic, flavonoid and proline contents in Burkina Fasan honey, as well as their radical scavenging activity. Food Chem. 2005, 91, 571–577. [Google Scholar] [CrossRef]
- Serna, M.; Hernández, F.; Coll, F.; Coll, Y.; Amorós, A. Effects of brassinosteroid analogues on total phenols, antioxidant activity, sugars, organic acids and yield of field grown endive (Cichorium endivia L.). J. Sci. Food Agric. 2013, 93, 1765–1771. [Google Scholar] [CrossRef]
- Costache, M.A.; Campeanu, G.; Neata, G. Studies concerning the extraction of chlorophyll and total carotenoids from vegetables. Rom. Biotechnol. Lett. 2012, 17, 7702–7708. [Google Scholar]
- Fogg, D.N.; Wilkinson, A.N. The colorimetric determination of phosphorus. Analyst 1958, 83, 406–414. [Google Scholar] [CrossRef]
- Morand, P.; Gullo, J.L. Mineralisation des tissus vegetaux en vue du dosage de P, Ca, Mg, Na, K. Ann. Agron. 1970, 21, 229–236. [Google Scholar]
- Colla, G.; Cardarelli, M.; Bonini, P.; Rouphael, Y. Foliar Applications of Protein Hydrolysate, Plant and Seaweed Extracts Increase Yield but Differentially Modulate Fruit Quality of Greenhouse Tomato. Horts 2017, 52, 1214–1220. [Google Scholar] [CrossRef]
- Searchinger, T.; Hanson, C.; Ranganathan, J.; Lipinski, B.; Waite, R.; Winterbottom, R.; Dinshaw, A.; Heimlich, R. The Great Balancing Act; World Resources Institite: Washington, DC, USA, 2013. [Google Scholar]
- Zaidi, A.; Khan, M.S. Microbial Strategies for Vegetable Production; Springer International Publishing: Cham, Switzerland, 2017; ISBN 978-3-319-54400-7. [Google Scholar]
- Gravel, V.; Antoun, H.; Tweddell, R.J. Growth Stimulation and Fruit Yield Improvement of Greenhouse Tomato Plants by Inoculation with Pseudomonas Putida or Trichoderma Atroviride: Possible Role of Indole Acetic Acid (IAA). Soil Biol. Biochem. 2007, 39, 1968–1977. [Google Scholar] [CrossRef]
- Bhattacharyya, P.N.; Jha, D.K. Plant Growth-Promoting Rhizobacteria (PGPR): Emergence in Agriculture. World J. Microbiol. Biotechnol. 2012, 28, 1327–1350. [Google Scholar] [CrossRef]
- Singh, A.; Maji, S.; Kumar, S. Effect of biofertilizers on yield and biomolecules of anti-cancerous vegetable broccoli. Int. J. Bio-Resour. Stress Manag. 2014, 5, 262–268. [Google Scholar] [CrossRef]
- Mantelin, S.; Touraine, B. Plant Growth-promoting Bacteria and Nitrate Availability: Impacts on Root Development and Nitrate Uptake. J. Exp. Bot. 2004, 55, 27–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parewa, H.P.; Yadav, J.; Rakshit, A.; Meena, V.S.; Karthikeyan, N. Plant growth promoting rhizobacteria enhance growth and nutrient uptake of crops. Agric. Sustain. Dev. 2014, 2, 101–116. [Google Scholar]
- Ordookhani, K.; Zare, M. Effect of Pseudomonas, Azotobacter and arbuscular mycorrhiza fungi on lycopene, antioxidant activity and total soluble solid in tomato (Lycopersicon esculentum F1 Hybrid, Delba). Adv. Environ. Biol. 2011, 5, 1290–1294. [Google Scholar]
- Katsenios, N.; Andreou, V.; Sparangis, P.; Djordjevic, N.; Giannoglou, M.; Chanioti, S.; Stergiou, P.; Xanthou, M.-Z.; Kakabouki, I.; Vlachakis, D.; et al. Evaluation of Plant Growth Promoting Bacteria Strains on Growth, Yield and Quality of Industrial Tomato. Microorganisms 2021, 9, 2099. [Google Scholar] [CrossRef]
- Fernández-Vázquez, R.; Stinco, C.M.; Meléndez-Martínez, A.J.; Heredia, F.J.; Vicario, I.M. Visual and Instrumental Evaluation of Orange Juice Color: A Consumers’ Preference Study. J. Sens. Stud. 2011, 26, 436–444. [Google Scholar] [CrossRef]
- Bojović, B.; Marković, A. Correlation between nitrogen and chlorophyll content in wheat (Triticum aestivum L.). Kragujev. J. Sci. 2009, 31, 69–74. [Google Scholar]
- Cappellari, L.d.R.; Chiappero, J.; Santoro, M.V.; Giordano, W.; Banchio, E. Inducing Phenolic Production and Volatile Organic Compounds Emission by Inoculating Mentha Piperita with Plant Growth-Promoting Rhizobacteria. Sci. Hortic. 2017, 220, 193–198. [Google Scholar] [CrossRef]
- Kloepper, J.W. Plant-growth-promoting rhizobacteria as biological control agents. In Metting Soil Microbial Ecology: Applications in Agricultural and Environmental Management; Marcel Dekker Inc.: New York, NY, USA, 1993; pp. 255–273. [Google Scholar]
- Van Loon, L.C. Plant response to plant growth-promoting rhizobacteria. Eur. J. Plant Pathol. 2007, 119, 243–254. [Google Scholar] [CrossRef] [Green Version]
- Babalola, O.O. Beneficial bacteria of agricultural importance. Biotechnol. Lett. 2010, 32, 1559–1570. [Google Scholar] [CrossRef]
- Ottaiano, L.; Di Mola, I.; Cozzolino, E.; El-Nakhel, C.; Rouphael, Y.; Mori, M. Biostimulant Application under Different Nitrogen Fertilization Levels: Assessment of Yield, Leaf Quality, and Nitrogen Metabolism of Tunnel-Grown Lettuce. Agronomy 2021, 11, 1613. [Google Scholar] [CrossRef]
- Di Mola, I.; Cozzolino, E.; Ottaiano, L.; Giordano, M.; Rouphael, Y.; Colla, G.; Mori, M. Effect of vegetal- and seaweed extractbased biostimulants on agronomical and leaf quality traits of plastic tunnel-grown baby lettuce under four regimes of nitrogen fertilization. Agronomy 2019, 9, 571. [Google Scholar] [CrossRef] [Green Version]
- Radhakrishnan, R.; Lee, I.J. Gibberellins producing Bacillus methylotrophicus KE2 supports plant growth and enhances nutritional metabolites and food values of lettuce. Plant Physiol. Biochem. 2016, 109, 181–189. [Google Scholar] [CrossRef]
- Pinto, E.; Almeida, A.A.; Aguiar, A.A.R.M.; Ferreira, I.M.P.L.V.O. Changes in Macrominerals, Trace Elements and Pigments Content during Lettuce (Lactuca Sativa L.) Growth: Influence of Soil Composition. Food Chem. 2014, 152, 603–611. [Google Scholar] [CrossRef] [PubMed]
- de Santi Ferrara, F.I.; Oliveira, Z.M.; Gonzales, H.H.S.; Floh, E.I.S.; Barbosa, H.R. Endophytic and Rhizospheric Enterobacteria Isolated from Sugar Cane Have Different Potentials for Producing Plant Growth-Promoting Substances. Plant Soil 2012, 353, 409–417. [Google Scholar] [CrossRef]
- Hungria, M.; Ribeiro, R.A.; Nogueira, M.A. Draft Genome Sequences of Azospirillum Brasilense Strains Ab-V5 and Ab-V6, Commercially Used in Inoculants for Grasses and Legumes in Brazil. Genome Announc. 2018, 6, e00393-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hungria, M.; Campo, R.J.; Souza, E.M.; Pedrosa, F.O. Inoculation with Selected Strains of Azospirillum Brasilense and A. Lipoferum Improves Yields of Maize and Wheat in Brazil. Plant Soil 2010, 331, 413–425. [Google Scholar] [CrossRef]
- Rouphael, Y.; Carillo, P.; Colla, G.; Fiorentino, N.; Sabatino, L.; El-Nakhel, C.; Giordano, M.; Pannico, A.; Cirillo, V.; Shabani, E.; et al. Appraisal of Combined Applications of Trichoderma virens and a Biopolymer-Based Biostimulant on Lettuce Agronomical, Physiological, and Qualitative Properties under Variable N Regimes. Agronomy 2020, 10, 196. [Google Scholar] [CrossRef] [Green Version]
- Sandhya, V.; Ali, S.Z.; Grover, M.; Reddy, G.; Venkateswarlu, B. Effect of Plant Growth Promoting Pseudomonas spp. on Compatible Solutes, Antioxidant Status and Plant Growth of Maize under Drought Stress. Plant Growth Regul. 2010, 62, 21–30. [Google Scholar] [CrossRef]
- Pırlak, L.; Köse, M. Effects of Plant Growth Promoting Rhizobacteria on Yield and Some Fruit Properties of Strawberry. J. Plant Nutr. 2009, 32, 1173–1184. [Google Scholar] [CrossRef]
- Bénard, C.; Gautier, H.; Bourgaud, F.; Grasselly, D.; Navez, B.; Caris-Veyrat, C.; Weiss, M.; Génard, M. Effects of Low Nitrogen Supply on Tomato (Solanum Lycopersicum) Fruit Yield and Quality with Special Emphasis on Sugars, Acids, Ascorbate, Carotenoids, and Phenolic Compounds. J. Agric. Food Chem. 2009, 57, 4112–4123. [Google Scholar] [CrossRef] [PubMed]
- Zeffa, D.M.; Perini, L.J.; Silva, M.B.; de Sousa, N.V.; Scapim, C.A.; de Oliveira, A.L.M.; Júnior, A.T.D.; Gonçalves, L.S.A. Azospirillum brasilense Promotes Increases in Growth and Nitrogen Use Efficiency of Maize Genotypes. PLoS ONE 2019, 14, e0215332. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Treatments | Plant Height (cm) | Root Collar Diameter (mm) | Number of Leaves (No. plant−1) | Head Fresh Weight (g plant−1) | Head Dry Matter (%) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| N dose (kg ha−1) | PGPB | ||||||||||
| 0 | Non-inoculated | 24.4 | g | 16.8 | i | 24.0 | d | 292.9 | l | 12.5 | g |
| A. brasilense DSM 1690 | 27.9 | d | 20.1 | h | 28.3 | b | 633.4 | h | 14.2 | c | |
| A. brasilense DSM 2298 | 30.2 | c | 28.5 | a | 25.7 | c | 637.4 | h | 13.8 | d | |
| Pseudomonas sp. DSM 25356 | 25.2 | f | 12.8 | j | 23.7 | e | 457.0 | k | 13.3 | e | |
| 30 | Non-inoculated | 28.4 | d | 28.5 | a | 24.7 | cd | 565.0 | j | 12.7 | g |
| A. brasilense DSM 1690 | 31.5 | b | 22.9 | f | 25.3 | c | 751.3 | c | 14.7 | c | |
| A. brasilense DSM 2298 | 32.8 | a | 24.1 | e | 24.7 | cd | 835.3 | a | 14.0 | c | |
| Pseudomonas sp. DSM 25356 | 28.0 | d | 21.8 | g | 30.0 | a | 653.5 | g | 13.6 | de | |
| 60 | Non-inoculated | 29.0 | c | 28.9 | a | 28.7 | ab | 631.1 | h | 13.1 | f |
| A. brasilense DSM 1690 | 32.0 | a | 23.8 | ef | 30.3 | a | 603.5 | i | 14.9 | b | |
| A. brasilense DSM 2298 | 26.9 | e | 25.3 | d | 26.0 | c | 675.3 | e | 14.3 | c | |
| Pseudomonas sp. DSM 25356 | 30.2 | c | 27.1 | c | 25.7 | c | 725.6 | d | 13.8 | d | |
| 120 | Non-inoculated | 30.0 | c | 26.0 | d | 26.7 | bc | 630.1 | h | 13.5 | e |
| A. brasilense DSM 1690 | 25.8 | f | 28.0 | b | 27.7 | b | 658.1 | f | 15.5 | a | |
| A. brasilense DSM 2298 | 29.8 | cd | 24.2 | e | 26.3 | c | 816.3 | b | 15.1 | b | |
| Pseudomonas sp. DSM 25356 | 30.3 | c | 22.7 | f | 29.7 | a | 829.4 | a | 13.8 | d | |
| Significance | |||||||||||
| Nitrogen (N) | *** | *** | *** | *** | *** | ||||||
| PGPB | *** | *** | *** | *** | *** | ||||||
| N × PGPB | *** | *** | *** | *** | *** | ||||||
| Treatments | SSC (°Brix) | a* | b* | L* | |||||
|---|---|---|---|---|---|---|---|---|---|
| N dose (kg ha−1) | PGPB | ||||||||
| 0 | Non-inoculated | 4.1 | b | −19.32 | c | 26.87 | c | 46.50 | cd |
| A. brasilense DSM 1690 | 4.1 | b | −20.67 | d | 28.30 | c | 41.70 | e | |
| A. brasilense DSM 2298 | 4.0 | c | −17.09 | b | 22.58 | e | 45.39 | d | |
| Pseudomonas sp. DSM 25356 | 4.1 | a | −19.40 | c | 32.46 | a | 53.10 | b | |
| 30 | Non-inoculated | 3.2 | e | −17.26 | b | 24.85 | d | 52.17 | b |
| A. brasilense DSM 1690 | 4.4 | a | −19.34 | c | 32.61 | a | 44.65 | d | |
| A. brasilense DSM 2298 | 4.2 | b | −11.30 | a | 24.39 | d | 44.36 | d | |
| Pseudomonas sp. DSM 25356 | 4.1 | b | −17.96 | b | 33.34 | a | 52.11 | b | |
| 60 | Non-inoculated | 3.7 | d | −17.87 | b | 30.55 | b | 52.96 | b |
| A. brasilense DSM 1690 | 4.2 | b | −20.74 | d | 33.67 | a | 47.36 | c | |
| A. brasilense DSM 2298 | 4.3 | a | −18.82 | c | 25.11 | d | 44.67 | d | |
| Pseudomonas sp. DSM 25356 | 4.0 | c | −19.63 | c | 31.15 | b | 47.60 | c | |
| 120 | Non-inoculated | 4.2 | b | −16.81 | b | 25.74 | d | 45.50 | d |
| A. brasilense DSM 1690 | 4.1 | b | −17.13 | b | 25.49 | d | 48.05 | c | |
| A. brasilense DSM 2298 | 4.1 | b | −20.22 | d | 23.18 | e | 44.18 | d | |
| Pseudomonas sp. DSM 25356 | 4.5 | a | −17.00 | b | 26.17 | c | 54.31 | a | |
| Significance | |||||||||
| Nitrogen (N) | * | *** | *** | *** | |||||
| PGPB | *** | *** | *** | *** | |||||
| N × PGPB | *** | *** | *** | *** | |||||
| Treatments | Ascorbic Acid (mg g−1 fw) | Total Phenolics (µg g−1 fw) | Carotenoids (mg 100 g−1 fw) | Total Chlorophyll (mg 100 g−1 fw) | N (mg g−1 dw) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| N dose (kg ha−1) | PGPB | ||||||||||
| 0 | Non-inoculated | 28.28 | f | 47.90 | d | 14.94 | g | 33.47 | h | 25.66 | i |
| A. brasilense DSM 1690 | 39.06 | a | 55.38 | a | 17.52 | e | 38.32 | c | 30.27 | f | |
| A. brasilense DSM 2298 | 36.84 | b | 51.15 | b | 17.23 | e | 36.74 | f | 28.98 | g | |
| Pseudomonas sp. DSM 25356 | 35.65 | c | 49.76 | c | 16.70 | f | 35.81 | g | 25.56 | i | |
| 30 | Non-inoculated | 28.14 | f | 47.52 | d | 15.59 | g | 35.37 | g | 28.67 | h |
| A. brasilense DSM 1690 | 39.18 | a | 55.65 | a | 20.96 | b | 40.50 | b | 33.46 | d | |
| A. brasilense DSM 2298 | 36.51 | b | 51.51 | b | 18.67 | d | 37.67 | d | 30.18 | f | |
| Pseudomonas sp. DSM 25356 | 34.94 | d | 49.80 | c | 17.20 | ef | 36.96 | e | 29.43 | g | |
| 60 | Non-inoculated | 27.78 | f | 47.52 | d | 15.52 | g | 35.45 | g | 30.46 | f |
| A. brasilense DSM 1690 | 39.35 | a | 55.49 | a | 22.88 | a | 40.43 | b | 35.47 | b | |
| A. brasilense DSM 2298 | 36.46 | bc | 51.55 | b | 19.52 | c | 37.86 | d | 31.55 | e | |
| Pseudomonas sp. DSM 25356 | 34.44 | d | 49.61 | c | 17.23 | e | 36.88 | e | 31.69 | e | |
| 120 | Non-inoculated | 27.22 | g | 45.54 | e | 15.48 | g | 35.41 | g | 34.36 | c |
| A. brasilense DSM 1690 | 37.27 | b | 52.01 | b | 22.79 | a | 42.53 | a | 38.59 | a | |
| A. brasilense DSM 2298 | 34.99 | d | 50.26 | c | 19.48 | c | 38.76 | c | 34.70 | c | |
| Pseudomonas sp. DSM 25356 | 30.98 | e | 48.18 | d | 17.36 | e | 37.28 | e | 35.52 | b | |
| Significance | |||||||||||
| Nitrogen (N) | *** | *** | *** | *** | *** | ||||||
| PGPB | *** | *** | *** | *** | *** | ||||||
| N × PGPB | *** | ** | *** | *** | *** | ||||||
| Treatments | NUE (t kg−1) | UE (kg kg−1) | |||
|---|---|---|---|---|---|
| N dose (kg ha−1) | PGPB | ||||
| 30 | Non-inoculated | 3.01 | d | 11.00 | d |
| A. brasilense DSM 1690 | 4.01 | b | 19.66 | a | |
| A. brasilense DSM 2298 | 4.45 | a | 18.87 | b | |
| Pseudomonas sp. DSM 25356 | 3.49 | c | 13.92 | c | |
| 60 | Non-inoculated | 1.68 | g | 6.70 | f |
| A. brasilense DSM 1690 | 1.61 | h | 8.50 | e | |
| A. brasilense DSM 2298 | 1.80 | f | 8.13 | e | |
| Pseudomonas sp. DSM 25356 | 1.94 | e | 8.44 | e | |
| 120 | Non-inoculated | 0.84 | j | 3.91 | h |
| A. brasilense DSM 1690 | 0.88 | j | 5.26 | g | |
| A. brasilense DSM 2298 | 1.09 | i | 5.69 | g | |
| Pseudomonas sp. DSM 25356 | 1.11 | i | 5.43 | g | |
| Significance | |||||
| Nitrogen (N) | *** | *** | |||
| PGPB | *** | *** | |||
| N × PGPB | *** | *** | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Consentino, B.B.; Aprile, S.; Rouphael, Y.; Ntatsi, G.; De Pasquale, C.; Iapichino, G.; Alibrandi, P.; Sabatino, L. Application of PGPB Combined with Variable N Doses Affects Growth, Yield-Related Traits, N-Fertilizer Efficiency and Nutritional Status of Lettuce Grown under Controlled Condition. Agronomy 2022, 12, 236. https://doi.org/10.3390/agronomy12020236
Consentino BB, Aprile S, Rouphael Y, Ntatsi G, De Pasquale C, Iapichino G, Alibrandi P, Sabatino L. Application of PGPB Combined with Variable N Doses Affects Growth, Yield-Related Traits, N-Fertilizer Efficiency and Nutritional Status of Lettuce Grown under Controlled Condition. Agronomy. 2022; 12(2):236. https://doi.org/10.3390/agronomy12020236
Chicago/Turabian StyleConsentino, Beppe Benedetto, Simona Aprile, Youssef Rouphael, Georgia Ntatsi, Claudio De Pasquale, Giovanni Iapichino, Pasquale Alibrandi, and Leo Sabatino. 2022. "Application of PGPB Combined with Variable N Doses Affects Growth, Yield-Related Traits, N-Fertilizer Efficiency and Nutritional Status of Lettuce Grown under Controlled Condition" Agronomy 12, no. 2: 236. https://doi.org/10.3390/agronomy12020236
APA StyleConsentino, B. B., Aprile, S., Rouphael, Y., Ntatsi, G., De Pasquale, C., Iapichino, G., Alibrandi, P., & Sabatino, L. (2022). Application of PGPB Combined with Variable N Doses Affects Growth, Yield-Related Traits, N-Fertilizer Efficiency and Nutritional Status of Lettuce Grown under Controlled Condition. Agronomy, 12(2), 236. https://doi.org/10.3390/agronomy12020236