
The Effect of Glucosinolates on the Growth and Development of Helicoverpa armigera Larvae and the Expression of Midgut Sulfatase Genes



Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Bioassay
2.3. Identification and Phylogenetic Tree Construction of HaSulf
2.4. Expression of HaSulfs and HaSumf1
2.5. Determination of Protein Production in the Midgut
2.6. Statistical Analysis
3. Results
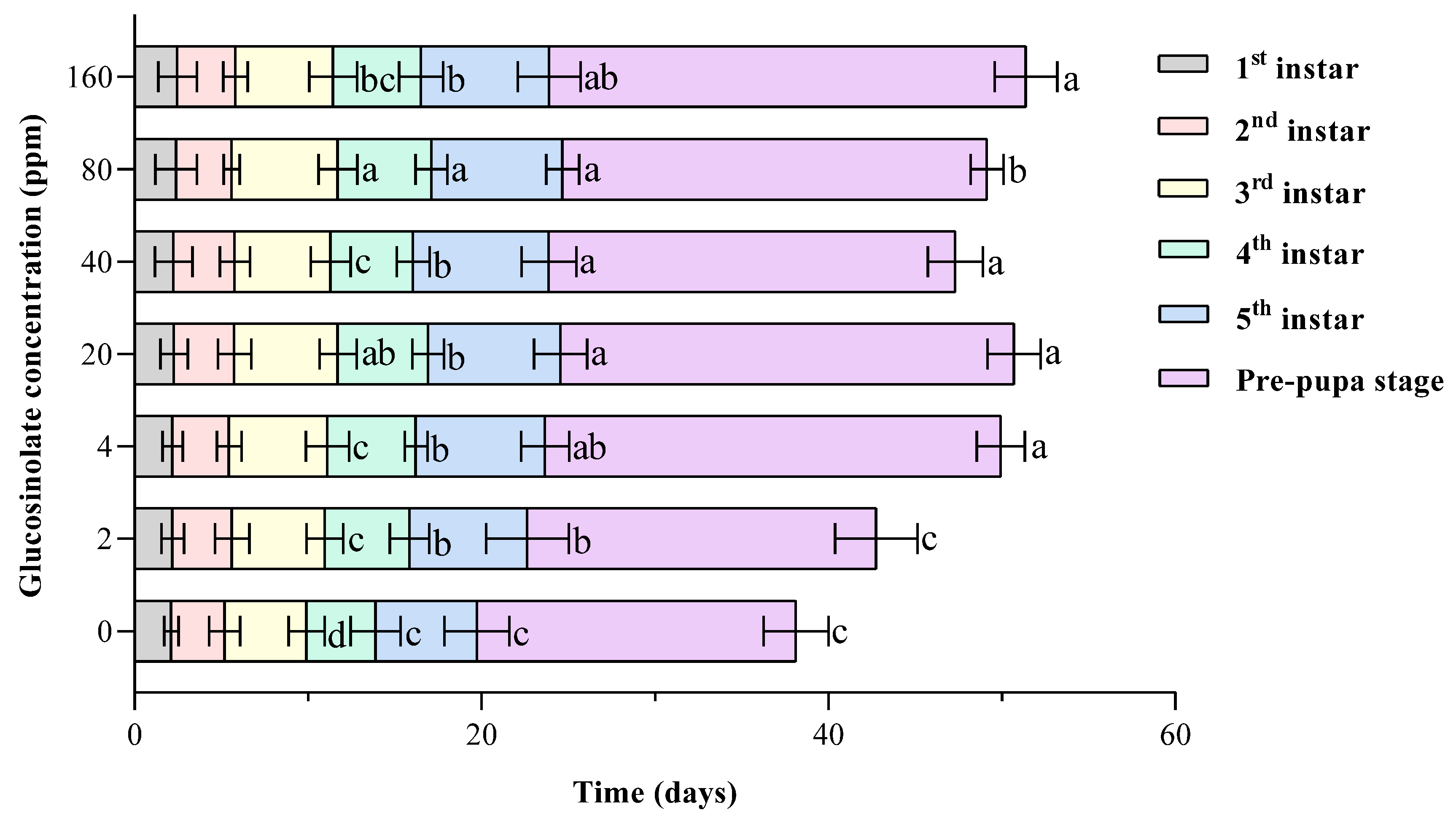
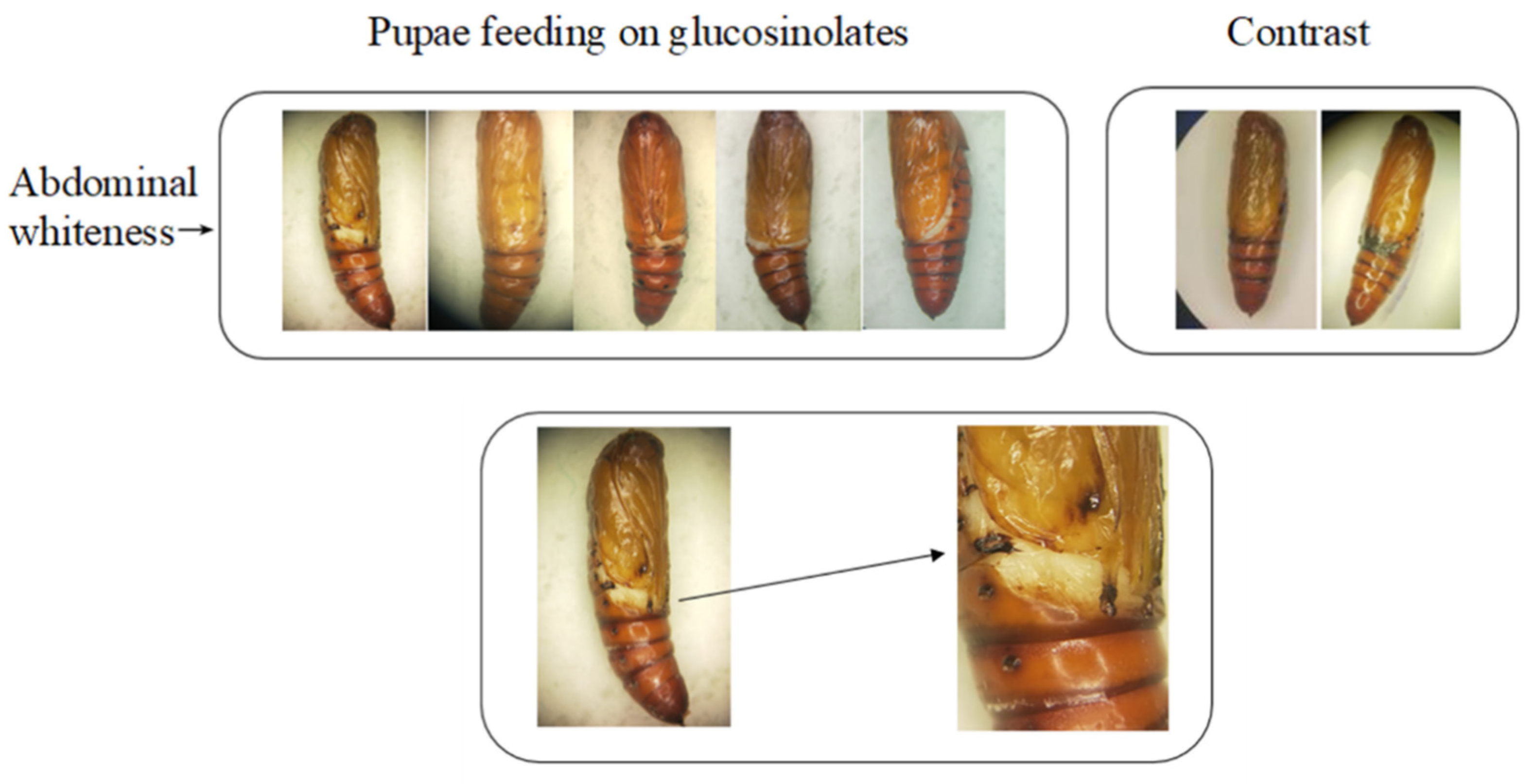
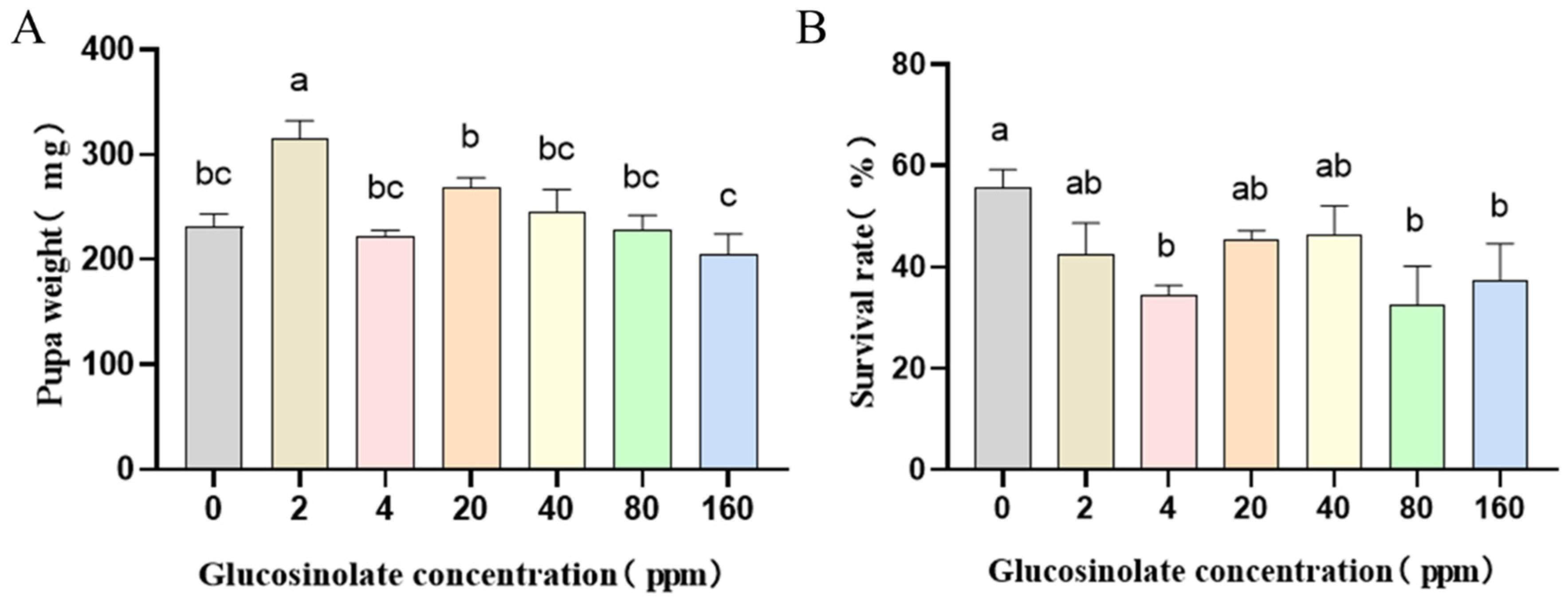
3.1. Effects of Different Concentrations of Glucosinolate on the Growth and Development of H. armigera
3.2. Identification of HaSulf Gene and Protein Prediction
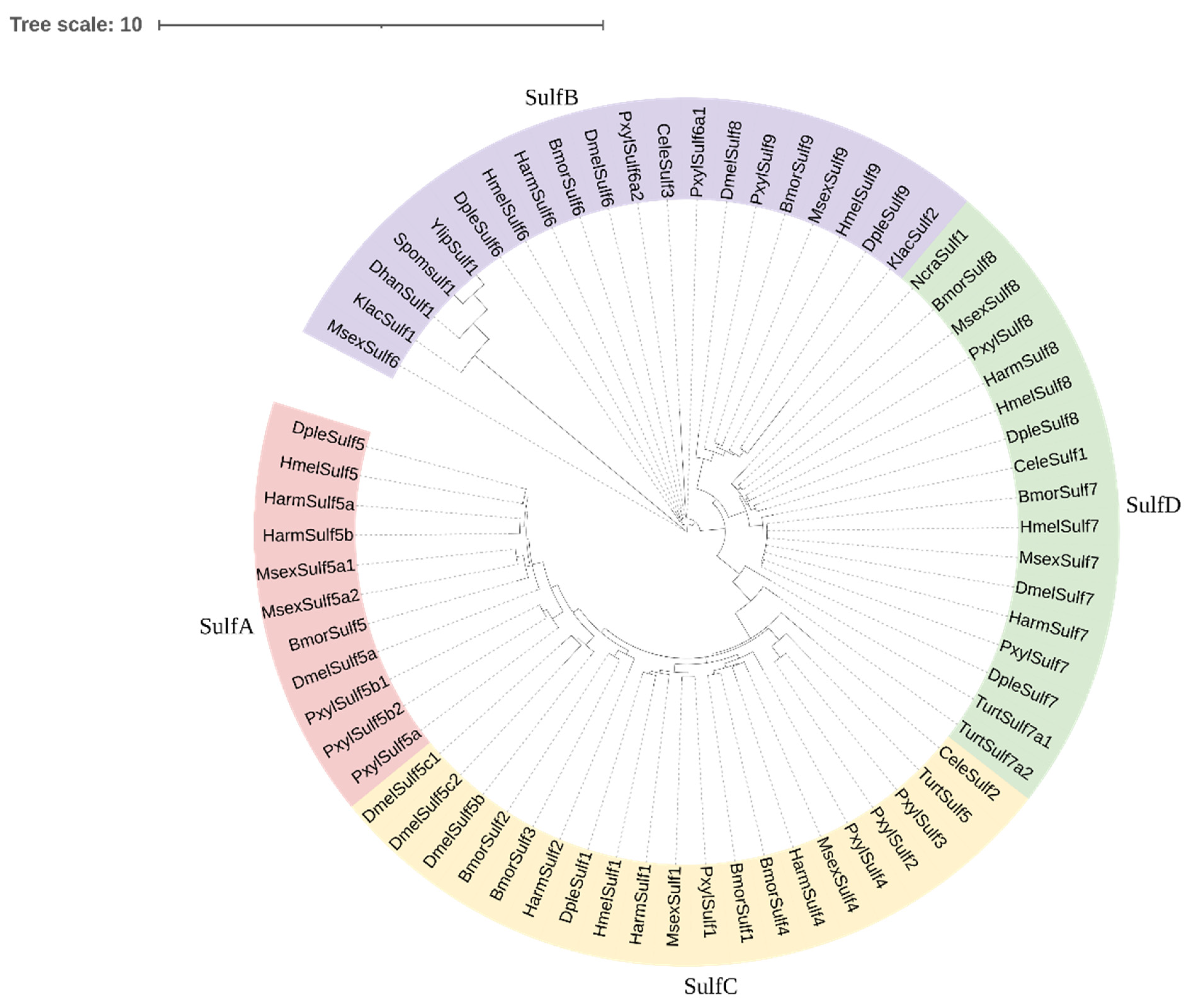
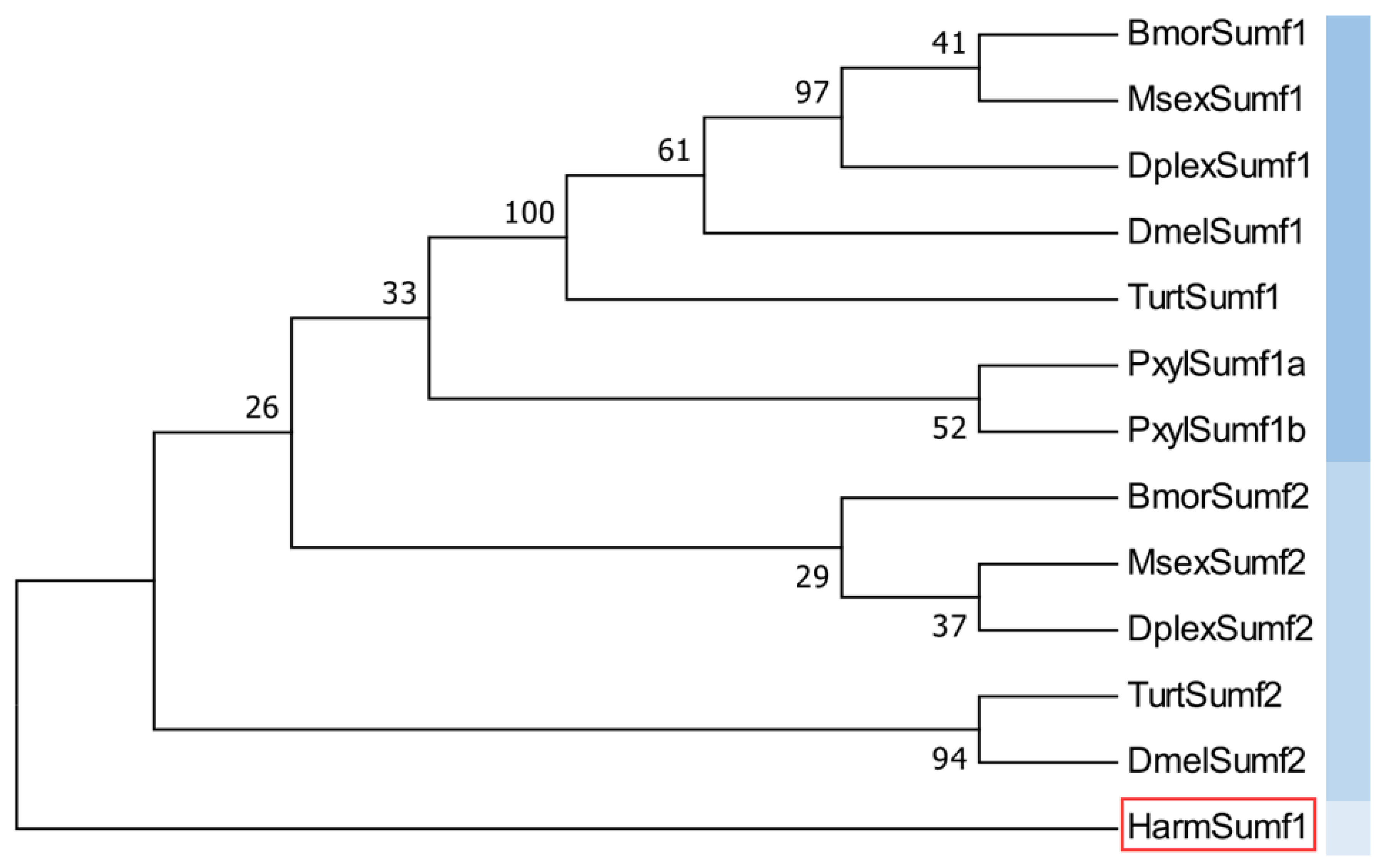
3.3. Prediction of Conserved Domains of HaSulfs and Construction of Phylogenetic Tree
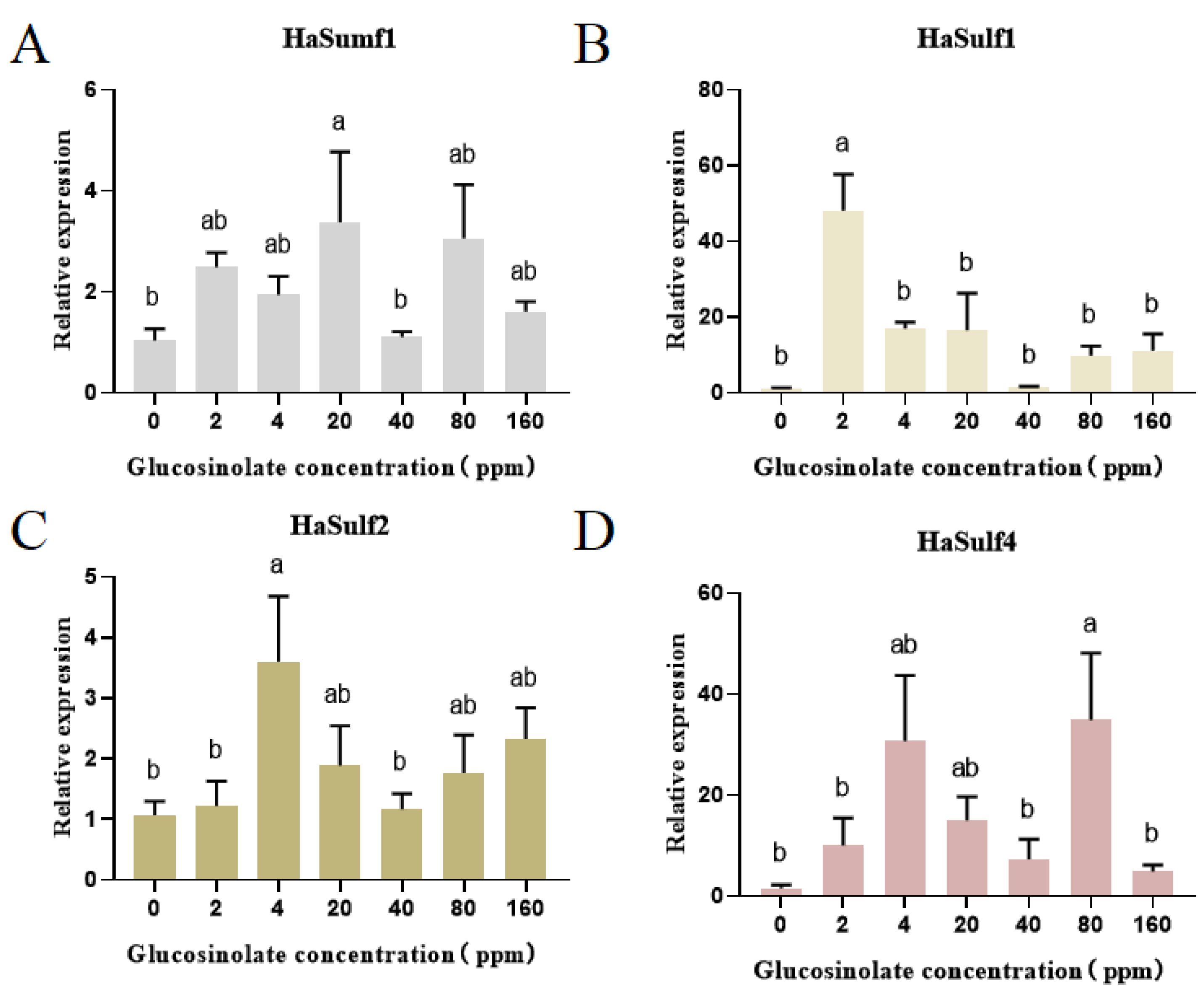
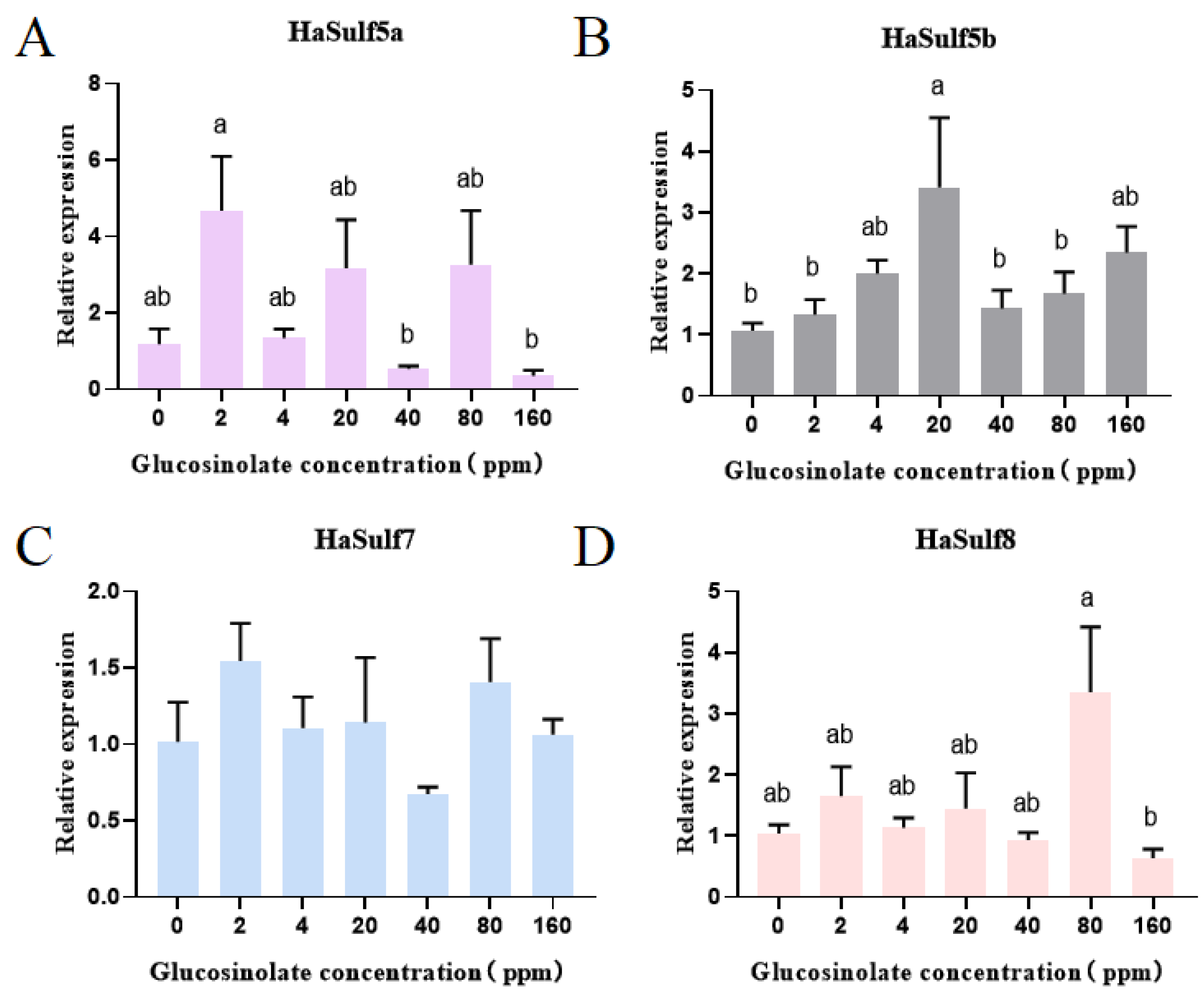
3.4. RT-qPCR Analysis of the Effects of Different Concentrations of Glucosinolate on the HaSulf Genes of H. armigera
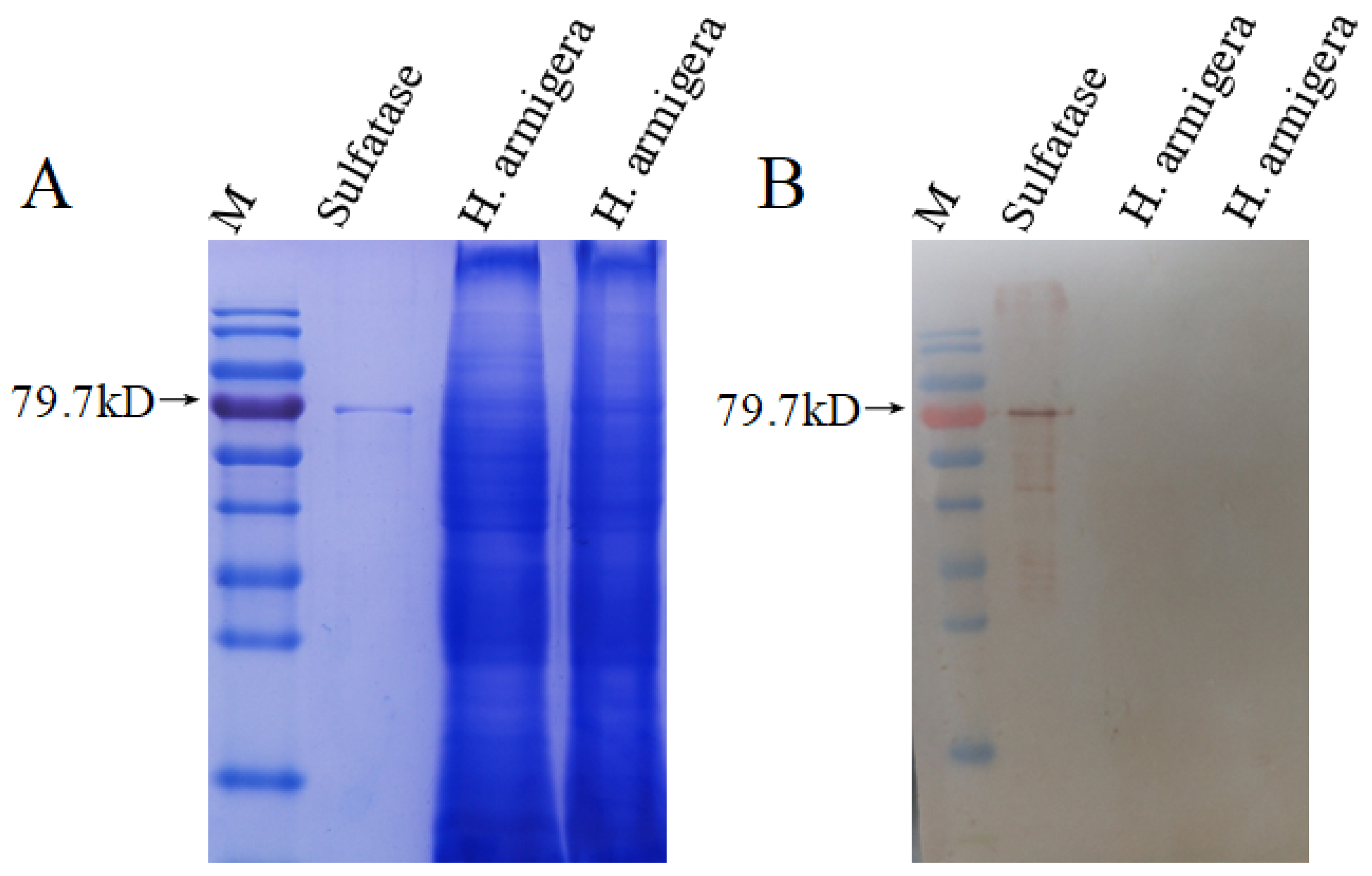
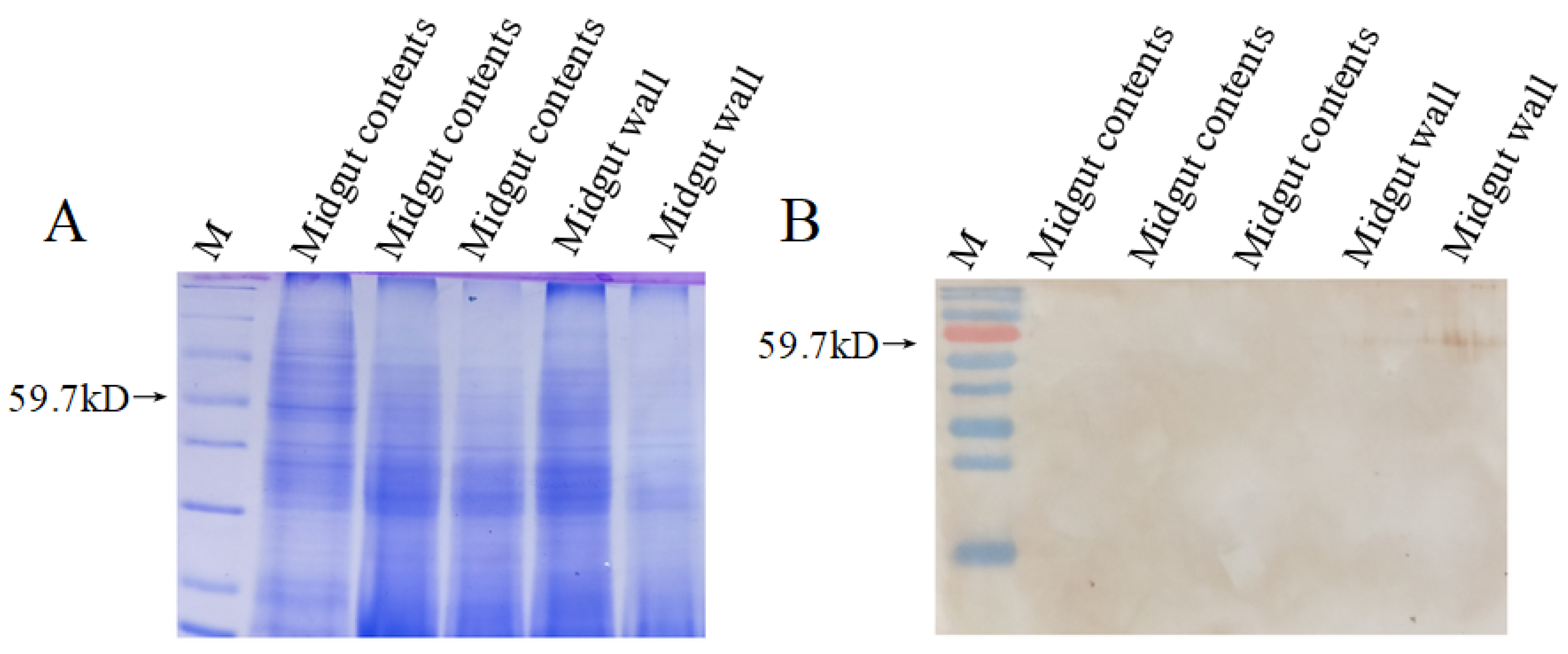
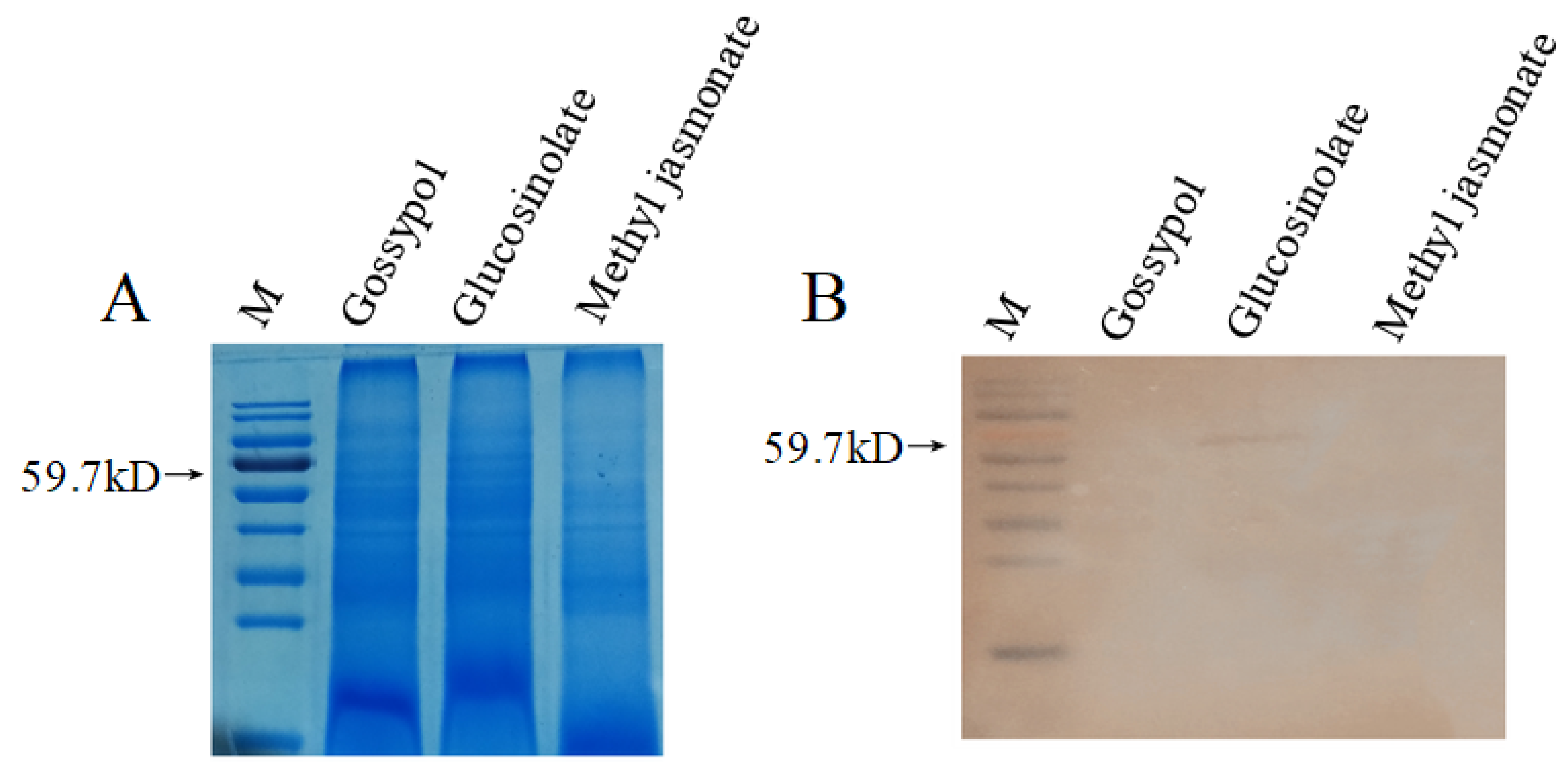
3.5. Western Blot Analysis of HaSulf Production at the Protein Level
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Tay, W.T.; Soria, M.F.; Walsh, T.; Thomazoni, D.; Silvie, P.; Behere, G.; Anderson, C.; Downes, S. A Brave New World for an Old World Pest: Helicoverpa armigera (Lepidoptera: Noctuidae) in Brazil. PLoS ONE 2013, 8, e80134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, P.; Li, X.; Wang, C.; Zhu, S.; Li, Q.; Zhang, Y.; Li, X.; Li, G.; Liu, E.; Gao, H.; et al. The Sensitivity of Field Populations of Metopolophium dirhodum (Walker) (Hemiptera: Aphididae) to Seven Insecticides in Northern China. Agronomy 2021, 11, 1556. [Google Scholar] [CrossRef]
- Falk, K.L.; Kastner, J.; Bodenhausen, N.; Schramm, K.; Paetz, C.; Vassão, D.G.; Reichelt, M.; Von Knorre, D.; Bergelson, J.; Erb, M.; et al. The role of glucosinolates and the jasmonic acid pathway in resistance of Arabidopsis thaliana against molluscan herbivores. Mol. Ecol. 2014, 23, 1188–1203. [Google Scholar] [CrossRef] [Green Version]
- Lü, L.; Luo, J.; Zhang, S.; Yu, Q.; Ma, L.; Liu, X.; Wang, C.; Ma, X.; Ma, Y.; Cui, J. Efficiency of cotton bollworm (Helicoverpa armigera Hübner) control of different Bt cotton varieties in North China. J. Cotton Res. 2018, 1, 4. [Google Scholar] [CrossRef]
- Wei, J.; Liang, G.; Wang, B.; Zhong, F.; Chen, L.; Khaing, M.M.; Zhang, J.; Guo, Y.; Wu, K.; Tabashnik, B.E. Activation of Bt Protoxin Cry1Ac in Resistant and Susceptible Cotton Bollworm. PLoS ONE 2016, 11, e0156560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hopkins, R.J.; van Dam, N.M.; van Loon, J.J. Role of Glucosinolates in Insect-Plant Relationships and Multitrophic Interactions. Annu. Rev. Èntomol. 2009, 54, 57–83. [Google Scholar] [CrossRef] [PubMed]
- Wittstock, U.; Gershenzon, J. Constitutive plant toxins and their role in defense against herbivores and pathogens. Curr. Opin. Plant Biol. 2002, 5, 300–307. [Google Scholar] [CrossRef]
- Liu, J.; Abbas, I.; Noor, R. Development of Deep Learning-Based Variable Rate Agrochemical Spraying System for Targeted Weeds Control in Strawberry Crop. Agronomy 2021, 11, 1480. [Google Scholar] [CrossRef]
- Ahuja, I.; Rohloff, J.; Bones, A.M. Defence mechanisms of Brassicaceae: Implications for plant-insect interactions and potential for integrated pest management. A review. Sustain. Agric. Vol. 2010, 30, 311–348. [Google Scholar] [CrossRef] [Green Version]
- Giamoustaris, A.; Mithen, R. The effect of modifying the glucosinolate content of leaves of oilseed rape (Brassica napus ssp. oleifera) on its interaction with specialist and generalist pests. Ann. Appl. Biol. 1995, 126, 347–363. [Google Scholar] [CrossRef]
- Grbić, M.; Van Leeuwen, T.; Clark, R.M.; Rombauts, S.; Rouzé, P.; Grbić, V.; Osborne, E.J.; Dermauw, W.; Ngoc, P.C.T.; Ortego, F.; et al. The genome of Tetranychus urticae reveals herbivorous pest adaptations. Nature 2011, 479, 487–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarfraz, M.; Dosdall, L.M.; Keddie, B.A. Diamondback moth–host plant interactions: Implications for pest management. Crop Prot. 2005, 25, 625–639. [Google Scholar] [CrossRef]
- Grubb, C.D.; Abel, S. Glucosinolate metabolism and its control. Trends Plant Sci. 2006, 11, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Wittstock, U.; Halkier, B.A. Glucosinolate research in the Arabidopsis era. Trends Plant Sci. 2002, 7, 263–270. [Google Scholar] [CrossRef]
- Kliebenstein, D.; Kroymann, J.; Mitchell-Olds, T. The glucosinolate–myrosinase system in an ecological and evolutionary context. Curr. Opin. Plant Biol. 2005, 8, 264–271. [Google Scholar] [CrossRef]
- Fahey, J.W.; Zalcmann, A.T.; Talalay, P. The chemical diversity and distribution of glucosinolates and isothiocyanates among plants. Phytochemistry 2001, 56, 5–51. [Google Scholar] [CrossRef]
- Badenes-Pérez, F.R.; Reichelt, M.; Gershenzon, J.; Heckel, D.G. Phylloplane location of glucosinolates in Barbarea spp. (Brassicaceae) and misleading assessment of host suitability by a specialist herbivore. New Phytol. 2010, 189, 549–556. [Google Scholar] [CrossRef]
- Poelman, E.H.; Dam, N.M.; Loon, J.J.A.; Vet, L.E.M.; Dicke, M. Chemical diversity in Brassica oleracea affects biodiversity of insect herbivores. Ecology 2009, 90, 1863–1877. [Google Scholar] [CrossRef] [Green Version]
- Ahuja, I.; De Vos, R.C.H.; Rohloff, J.; Stoopen, G.M.; Halle, K.K.; Ahmad, S.J.N.; Hoang, L.; Hall, R.; Bones, A.M. Arabidopsis myrosinases link the glucosinolate-myrosinase system and the cuticle. Sci. Rep. 2016, 6, 38990. [Google Scholar] [CrossRef] [Green Version]
- Ali, J.; Agrawal, A.A. Specialist versus generalist insect herbivores and plant defense. Trends Plant Sci. 2012, 17, 293–302. [Google Scholar] [CrossRef]
- de Vos, M.; Kim, J.H.; Jander, G. Biochemistry and molecular biology of Arabidopsis–aphid interactions. BioEssays 2007, 29, 871–883. [Google Scholar] [CrossRef] [PubMed]
- Heckel, D.G. Insect detoxification and sequestration strategies. Annu. Plant Rev. 2014, 47, 77–114. [Google Scholar] [CrossRef]
- Müller, C. Interactions between glucosinolate- and myrosinase-containing plants and the sawfly Athalia rosae. Phytochem. Rev. 2008, 8, 121–134. [Google Scholar] [CrossRef]
- Winde, I.; Wittstock, U. Insect herbivore counteradaptations to the plant glucosinolate–myrosinase system. Phytochemistry 2011, 72, 1566–1575. [Google Scholar] [CrossRef]
- Beran, F.; Pauchet, Y.; Kunert, G.; Reichelt, M.; Wielsch, N.; Vogel, H.; Reinecke, A.; Svatos, A.; Mewis, I.; Schmid, D.; et al. Phyllotreta striolata flea beetles use host plant defense compounds to create their own glucosinolate-myrosinase system. Proc. Natl. Acad. Sci. USA 2014, 111, 7349–7354. [Google Scholar] [CrossRef] [Green Version]
- Ahn, S.-J.; Betzin, F.; Gikonyo, M.W.; Yang, Z.-L.; Köllner, T.G.; Beran, F. Identification and evolution of glucosinolate sulfatases in a specialist flea beetle. Sci. Rep. 2019, 9, 15725. [Google Scholar] [CrossRef]
- Chen, G.; Liu, H.; Mo, B.-C.; Hu, J.; Liu, S.-Q.; Bustos-Segura, C.; Xue, J.; Wang, X. Growth and Development of Helicoverpa armigera (Lepidoptera: Noctuidae) Larvae Infected by Heliothis virescens ascovirus 3i (HvAV-3i). Front. Physiol. 2020, 11, 93. [Google Scholar] [CrossRef]
- Kumar, S. Plant secondary metabolites (PSMs) of Brassicaceae and their role in plant defense against insect herbivores—A review. J. Appl. Nat. Sci. 2017, 9, 508–519. [Google Scholar] [CrossRef] [Green Version]
- Schramm, K.; Vassão, D.G.; Reichelt, M.; Gershenzon, J.; Wittstock, U. Metabolism of glucosinolate-derived isothiocyanates to glutathione conjugates in generalist lepidopteran herbivores. Insect Biochem. Mol. Biol. 2012, 42, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Schweizer, F.; Heidel-Fischer, H.; Vogel, H.; Reymond, P. Arabidopsis glucosinolates trigger a contrasting transcriptomic response in a generalist and a specialist herbivore. Insect Biochem. Mol. Biol. 2017, 85, 21–31. [Google Scholar] [CrossRef] [Green Version]
- Cosma, M.P.; Pepe, S.; Annunziata, I.; Newbold, R.F.; Grompe, M.; Parenti, G.; Ballabio, A. The Multiple Sulfatase Deficiency Gene Encodes an Essential and Limiting Factor for the Activity of Sulfatases. Cell 2003, 113, 445–456. [Google Scholar] [CrossRef]
- Fraldi, A.; Biffi, A.; Lombardi, A.; Visigalli, I.; Pepe, S.; Settembre, C.; Nusco, E.; Auricchio, A.; Naldini, L.; Ballabio, A.; et al. SUMF1 enhances sulfatase activities in vivo in five sulfatase deficiencies. Biochem. J. 2007, 403, 305–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, X.-L.; He, W.-Y.; Chen, W.; Xu, X.-J.; Qi, W.-P.; Zou, M.-M.; You, Y.-C.; Baxter, S.W.; Wang, P.; You, M.-S. Structure and expression of sulfatase and sulfatase modifying factor genes in the diamondback moth, Plutella xylostella. (L.). Insect Sci. 2017, 25, 946–958. [Google Scholar] [CrossRef] [PubMed]
- Claudianos, C.; Ranson, H.; Johnson, R.M.; Biswas, S.; Schuler, M.A.; Berenbaum, M.R.; Feyereisen, R.; Oakeshott, J.G. A deficit of detoxification enzymes: Pesticide sensitivity and environmental response in the honeybee. Insect Mol. Biol. 2006, 15, 615–636. [Google Scholar] [CrossRef] [Green Version]
- Fornes, O.; Castro-Mondragon, J.A.; Khan, A.; Van Der Lee, R.; Zhang, X.; Richmond, P.A.; Modi, B.; Correard, S.; Gheorghe, M.; Baranašić, D.; et al. JASPAR 2020: Update of the open-access database of transcription factor binding profiles. Nucleic Acids Res. 2019, 48, D87–D92. [Google Scholar] [CrossRef]
- Heidel-Fischer, H.M.; Kirsch, R.; Reichelt, M.; Ahn, S.-J.; Wielsch, N.; Baxter, S.W.; Heckel, D.; Vogel, H.; Kroymann, J. An Insect Counteradaptation against Host Plant Defenses Evolved through Concerted Neofunctionalization. Mol. Biol. Evol. 2019, 36, 930–941. [Google Scholar] [CrossRef]
- Sardiello, M.; Annunziata, I.; Roma, G.; Ballabio, A. Sulfatases and sulfatase modifying factors: An exclusive and promiscuous relationship. Hum. Mol. Genet. 2005, 14, 3203–3217. [Google Scholar] [CrossRef] [Green Version]
- Schlotawa, L.; Steinfeld, R.; von Figura, K.; Dierks, T.; Gärtner, J. Molecular analysis of SUMF1 mutations: Stability and residual activity of mutant formylglycine-generating enzyme determine disease severity in multiple sulfatase deficiency. Hum. Mutat. 2007, 29, 205. [Google Scholar] [CrossRef]
- Parenti, G.; Meroni, G.; Ballabio, A. The sulfatase gene family. Curr. Opin. Genet. Dev. 1997, 7, 386–391. [Google Scholar] [CrossRef]
- Chen, W.; Dong, Y.; Saqib, H.S.A.; Vasseur, L.; Zhou, W.; Zheng, L.; Lai, Y.; Ma, X.; Lin, L.; Xu, X.; et al. Functions of duplicated glucosinolate sulfatases in the development and host adaptation of Plutella xylostella. Insect Biochem. Mol. Biol. 2020, 119, 103316. [Google Scholar] [CrossRef]
- Manivannan, A.; Israni, B.; Luck, K.; Götz, M.; Seibel, E.; Easson, M.; Kirsch, R.; Reichelt, M.; Stein, B.; Winter, S.; et al. Identification of a sulfatase that detoxifies glucosinolates in the phloem-feeding insect Bemisia tabaci and prefers indolic glucosinolates. Front. Plant Sci. 2021, 12, 671286. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Peng, Z.; Yang, Z.; Tian, L.; Liu, S.; Wang, S.; Wu, Q.; Xie, W.; Zhang, Y. Genome-wide identification and analysis of sulfatase and sulfatase modifying factor genes in Bemisia tabaci (Hemiptera: Aleyrodidae). Insect Sci. 2021, 28, 1541–1552. [Google Scholar] [CrossRef] [PubMed]
- Agnihotri, A.R.; Hulagabali, C.V.; Adhav, A.S.; Joshi, R.S. Mechanistic insight in potential dual role of sinigrin against Helicoverpa armigera. Phytochemistry 2017, 145, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Beran, F.; Sporer, T.; Paetz, C.; Ahn, S.-J.; Betzin, F.; Kunert, G.; Shekhov, A.; Vassão, D.G.; Bartram, S.; Lorenz, S.; et al. One Pathway Is Not Enough: The Cabbage Stem Flea Beetle Psylliodes chrysocephala Uses Multiple Strategies to Overcome the Glucosinolate-Myrosinase Defense in Its Host Plants. Front. Plant Sci. 2018, 9, 1754. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer (5′-3′) |
|---|---|
| HaRPS15 qPCR F | CTGAGGTCGATGAAACTCTC |
| HaRPS15 qPCR R | CTCCATGAGTTGCTCATTG |
| HaSumf1 qPCR F | GCCAAAGATGGTTATGAAGG |
| HaSumf1 qPCR R | ATTCCCACACATTGCCAG |
| HaSulf1 qPCR F | GTTCGCTTCCGACAATGGAG |
| HaSulf1 qPCR R | GCATCAACCCTTGCCAAACT |
| HaSulf2 qPCR F | CGGTGATATAGTTGCAGCTTT |
| HaSulf2 qPCR R | CTCCACAATAACCCGACAACC |
| HaSulf5a qPCR F | ATGGAGTCCGCTACTGAAG |
| HaSulf5a qPCR R | ACACTAACATCGCCACCAG |
| HaSulf5b qPCR F | GTTGCTTCCACAGTTCCTG |
| HaSulf5b qPCR R | CGAATCCTCGGTTCATAGG |
| Protein Name | Gene ID | Gene Size (bp) | ORF (bp) | Protein (AA) | Location | Assembly |
|---|---|---|---|---|---|---|
| HaSumf1 | 110375229 | 2246 | 1020 | 339 | NW_018395484.1 (276539..279127) | Harm_1.0 (GCF_002156985.1) |
| HaSulf1 | 110375086 | 3156 | 1848 | 615 | NW_018395392.1 (3407398..3426461) | Harm_1.0 (GCF_002156985.1) |
| HaSulf2 | 110374783 | 2411 | 1608 | 535 | NW_018395392.1 (3399889..3404850) | Harm_1.0 (GCF_002156985.1) |
| HaSulf4 | 110375159 | 2877 | 1659 | 552 | NW_018395392.1 (3370118..3375945) | Harm_1.0 (GCF_002156985.1) |
| HaSulf5a | 110381566 | 1327 | 873 | 290 | NW_018395780.1 (177570..180958) | Harm_1.0 (GCF_002156985.1) |
| HaSulf5b | 110374404 | 1644 | 1617 | 538 | NW_018395467.1 (310504..316591) | Harm_1.0 (GCF_002156985.1) |
| HaSulf6 | 110370991 | 1806 | 1524 | 507 | NW_018395415.1 (788596..790955) | Harm_1.0 (GCF_002156985.1) |
| HaSulf7 | 110373702 | 5627 | 4212 | 1403 | NW_018395390.1 (3168620..3241586) | Harm_1.0 (GCF_002156985.1) |
| HaSulf8 | 110373657 | 1869 | 1491 | 496 | NW_018395449.1 (312589..321088) | Harm_1.0 (GCF_002156985.1) |
| Protein Name | Molecular Weight | Theoretical pI | Extinction Coefficients (M−1 cm−1) * | Estimated Half-Life (hours) ** | Instability Index | Aliphatic Index | Grand Average of Hydropathicity (GRAVY) |
|---|---|---|---|---|---|---|---|
| HaSumf1 | 38,590.95 | 5.64 | 82,110 | 30 | 38.17(stable) | 55.28 | −0.690 |
| HaSulf1 | 68,881.90 | 8.86 | 135,595 | 30 | 38.58(stable) | 83.85 | −0.338 |
| HaSulf2 | 59,732.77 | 5.19 | 104,670 | 30 | 36.61(stable) | 89.29 | −0.163 |
| HaSulf4 | 67,365.48 | 6.07 | 104,990 | 30 | 35.13(stable) | 90.15 | −0.204 |
| HaSulf5a | 32,028.26 | 6.45 | 54,680 | 30 | 41.02(unstable) | 87.83 | −0.358 |
| HaSulf5b | 59,969.67 | 6.63 | 99,280 | 30 | 33.03(stable) | 91.04 | −0.214 |
| HaSulf6 | 58,420.09 | 9.38 | 89,075 | 30 | 41.02(unstable) | 87.28 | −0.360 |
| HaSulf7 | 162,832.01 | 9.13 | 180,710 | 30 | 52.71(unstable) | 62.37 | −0.985 |
| HaSulf8 | 55,756.01 | 5.74 | 85,440 | 30 | 45.91(unstable) | 76.85 | −0.347 |
| Matrix ID | Name | Score | Relative Score | Sequence ID | Start | End | Strand | Predicted Sequence |
|---|---|---|---|---|---|---|---|---|
| MA0049.1 | MA0049.1.hb | 11.975525 | 0.9738739070180799 | HaSumf1 | 574 | 583 | + | GCAAAAAAAA |
| MA0015.1 | MA0015.1.Cf2 | 10.631421 | 0.9114732446482743 | HaSulf1 | 346 | 355 | + | GTATATGTTT |
| MA0049.1 | MA0049.1.hb | 10.612954 | 0.9318941952719153 | HaSulf2 | 78 | 87 | + | GAGAAAAAA |
| MA0049.1 | MA0049.1.hb | 11.291026 | 0.9527850486969089 | HaSulf4 | 510 | 519 | + | GAACAAAAA |
| MA0049.1 | MA0049.1.hb | 11.139023 | 0.9481019493665909 | HaSulf5a | 1072 | 1081 | + | GAATAAAAAT |
| MA0015.1 | MA0015.1.Cf2 | 11.832657 | 0.9412119301944221 | HaSulf5b | 63 | 72 | + | ATATATGTGT |
| MA0010.1 | MA0010.1.br | 12.489317 | 0.9206608060709957 | HaSulf6 | 399 | 412 | + | AAAATAAACAAAAG |
| MA0012.1 | MA0012.1.br | 8.726485 | 0.8802668436514 | HaSulf7 | 978 | 988 | + | TCAACTATAAC |
| MA0015.1 | MA0015.1.Cf2 | 8.444539 | 0.857333168027844 | HaSulf8 | 33 | 42 | + | ATAAATGTAC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Cai, F.; Kuerban, G.; Zhang, S.; Li, C.; Zhao, Y.; Jin, L.; Ma, X. The Effect of Glucosinolates on the Growth and Development of Helicoverpa armigera Larvae and the Expression of Midgut Sulfatase Genes. Agronomy 2022, 12, 306. https://doi.org/10.3390/agronomy12020306
Li X, Cai F, Kuerban G, Zhang S, Li C, Zhao Y, Jin L, Ma X. The Effect of Glucosinolates on the Growth and Development of Helicoverpa armigera Larvae and the Expression of Midgut Sulfatase Genes. Agronomy. 2022; 12(2):306. https://doi.org/10.3390/agronomy12020306
Chicago/Turabian StyleLi, Xue, Fan Cai, Guli Kuerban, Shaohui Zhang, Chaoyong Li, Ying Zhao, Lin Jin, and Xiaoli Ma. 2022. "The Effect of Glucosinolates on the Growth and Development of Helicoverpa armigera Larvae and the Expression of Midgut Sulfatase Genes" Agronomy 12, no. 2: 306. https://doi.org/10.3390/agronomy12020306
APA StyleLi, X., Cai, F., Kuerban, G., Zhang, S., Li, C., Zhao, Y., Jin, L., & Ma, X. (2022). The Effect of Glucosinolates on the Growth and Development of Helicoverpa armigera Larvae and the Expression of Midgut Sulfatase Genes. Agronomy, 12(2), 306. https://doi.org/10.3390/agronomy12020306