
Assessing the Response of Vitis vinifera L. cv. Xynisteri to Different Irrigation Regimes and Its Comparison to cvs. Maratheftiko, Shiraz and Sauvignon Blanc

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, Experimental Design and Treatments
2.2. Measurements
2.2.1. Climate
2.2.2. Vine Performance Measures
2.2.3. Physiology Measures
2.2.4. Stomatal Density
2.2.5. Statistical Analysis
3. Results
3.1. Climate
3.2. Vine Growth and Physiology Measurements
3.2.1. Cyprus Xynisteri Vineyard Trial
3.2.2. Potted Vine Trials
3.2.3. Stomatal Density
4. Discussion
4.1. Vine Growth Measurements
4.2. Water Potential
4.3. Stomatal Conductance
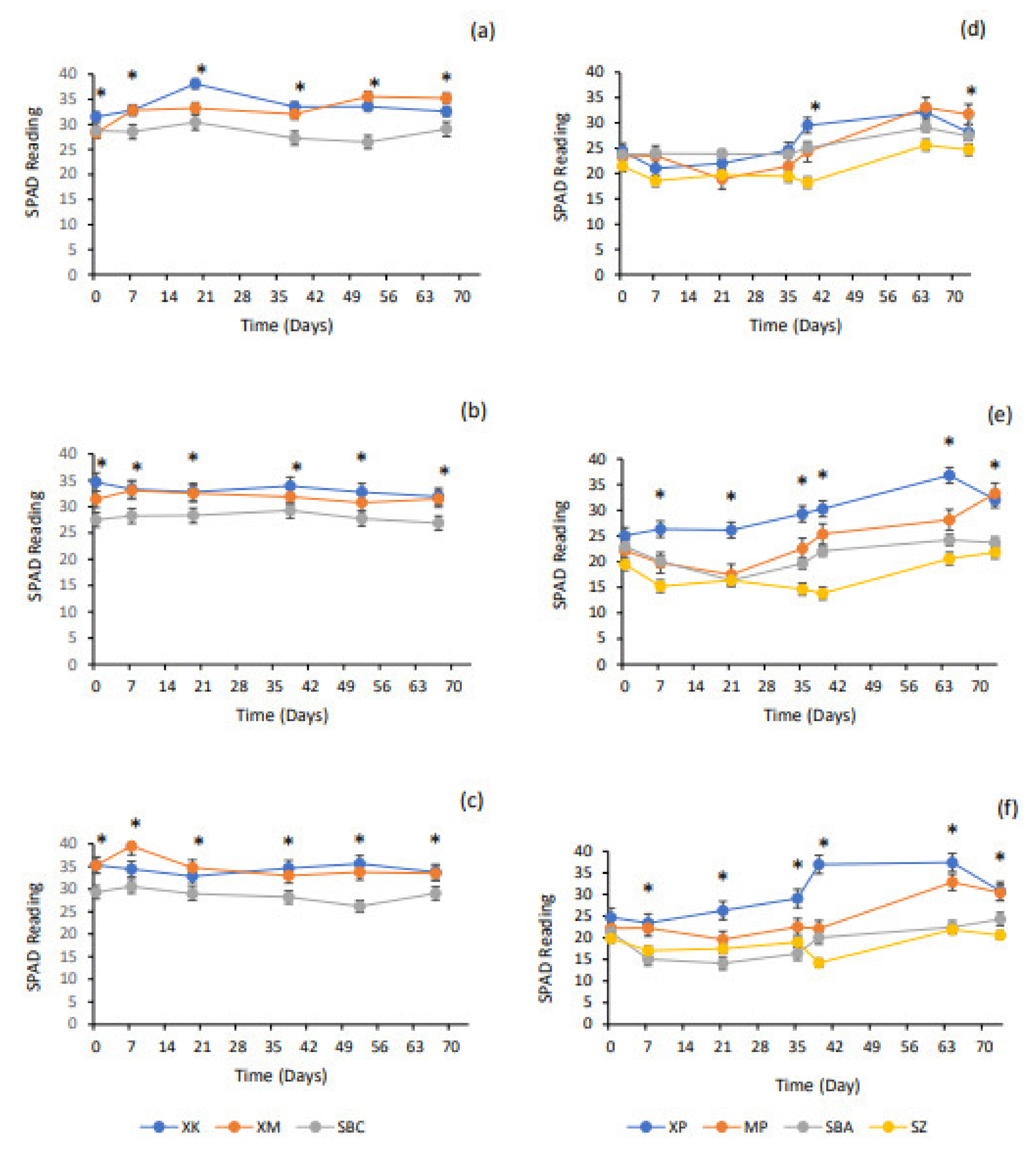
4.4. SPAD Readings (Indicative Chlorophyll Content)
4.5. Biomass
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jones, G.V.; White, M.A.; Cooper, O.R.; Storchmann, K. Climate change and global wine quality. Clim. Chang. 2005, 73, 319–343. [Google Scholar] [CrossRef]
- Schultz, H.R.; Jones, G.V. Climate induced historic and future changes in viticulture. J. Wine Res. 2010, 21, 137–145. [Google Scholar] [CrossRef]
- Camps, J.O.; Ramos, M.C. Grape harvest and yield responses to inter-annual changes in temperature and precipitation in an area of north-east Spain with a Mediterranean climate. Int. J. Biometeorol. 2012, 56, 853–864. [Google Scholar] [CrossRef] [PubMed]
- Webb, L.B. Adaptation of the Australian Wine Industry to Climate Change—Opportunities and Vulnerabilities. University of Melbourne and CSIRO, Australia. 2011. Available online: https://www.wineaustralia.com/research/projects/adaptation-of-the-australian-wine-indust (accessed on 1 July 2021).
- Keller, M. Managing grapevines to optimise fruit development in a challenging environment: A climate change primer for viticulturists. Aust. J. Grape Wine Res. 2010, 16, 56–69. [Google Scholar] [CrossRef]
- Webb, L.B.; Watterson, I.; Bhend, J.; Whetton, P.H.; Barlow, E.W.R. Global climate analogues for winegrowing regions in future periods: Projections of temperature and precipitation. Aust. J. Grape Wine Res. 2013, 19, 331–341. [Google Scholar] [CrossRef]
- Jarvis, C.; Darbyshire, R.; Goodwin, I.; Barlow, E.W.R.; Eckard, R. Advancement of winegrape maturity continuing for winegrowing regions in Australia with variable evidence of compression of the harvest period. Aust. J. Grape Wine Res. 2019, 25, 101–108. [Google Scholar] [CrossRef] [Green Version]
- Cook, B.; Wolkovich, E. Climate change decouples drought from early wine grape harvests in France. Nat. Clim. Chang. 2016, 6, 715–719. [Google Scholar] [CrossRef]
- Krieger, M.; Lohmann, G.; Laepple, T. Seasonal climate impacts on the grape harvest date in Burgundy (France). Clim. Past 2011, 7, 425–435. [Google Scholar] [CrossRef] [Green Version]
- Jones, G.V.; Duff, A.A.; Hall, A.; Myers, J.W. Spatial analysis of climate in winegrape growing regions in the Western United States. Am. J. Enol. Vitic. 2010, 61, 313–326. [Google Scholar]
- Van Leeuwen, C.; Schultz, H.R.; Garcia de Cortazar-Atauri, I.; Duchêne, E.; Ollat, B.; Pieri, P.; Bois, B.; Goutouly, J.-P.; Quénol, H.; Touzard, J.M.; et al. Why climate change will not dramatically decrease viticultural suitability in main wine-producing areas by 2050. Proc. Natl. Acad. Sci. USA 2013, 110, E3051–E3052. [Google Scholar] [CrossRef] [Green Version]
- Van Leeuwen, C.; Destrac-Irvine, A.; Dubernet, M.; Duchêne, E.; Gowdy, M.; Marguerit, E.; Pieri, P.; Parker, A.; de Rességuier, L.; Ollat, N. An update on the impact of climate change in viticulture and potential adaptations. Agronomy 2019, 9, 514. [Google Scholar] [CrossRef] [Green Version]
- Evans, J.P. 21st century climate change in the Middle East. Clim. Chang. 2009, 92, 417–432. [Google Scholar] [CrossRef]
- Hannah, L.; Roehrdanz, P.R.; Ikegami, M.; Shepard, A.V.; Shaw, M.R.; Tabor, G.; Zhi, L.; Marquet, P.A.; Hijmans, R.J. Climate change, wine, and conservation. Proc. Natl. Acad. Sci. USA 2013, 110, 6907–6912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Remenyi, T.A.; Rollins, D.A.; Love, P.T.; Earl, N.O.; Bindoff, N.L.; Harris, R.M.B. Australia’s Wine Future: A Climate Atlas; University of Tasmania: Hobart, TAS, Australia, 2019; Available online: http://ecite.utas.edu.au/137759 (accessed on 1 July 2021).
- Adamides, G. A review of climate-smart agriculture applications in Cyprus. Atmosphere 2020, 11, 898. [Google Scholar] [CrossRef]
- Grigoriou, A.; Tsaniklidis, G.; Hagidimitriou, M.; Nikoloudakis, N. The Cypriot indigenous grapevine germplasm is a multi-clonal varietal mixture. Plants 2020, 9, 1034. [Google Scholar] [CrossRef] [PubMed]
- Litskas, V.D.; Tzortzakis, N.; Stavrinides, M.C. Determining the carbon footprint and emission hotspots for the wine produced in Cyprus. Atmosphere 2020, 11, 463. [Google Scholar] [CrossRef]
- Vink, S.N.; Chrysargyris, A.; Tzortzakis, N.; Salles, J.F. Bacterial community dynamics varies with soil management and irrigation practices in grapevines (Vitis vinifera L.). Appl. Soil Ecol. 2021, 158, 103–107. [Google Scholar] [CrossRef]
- Heyman, L.; Chrysargyris, A.; Demeestere, K.; Tzortzakis, N.; Höfte, M. Responses to drought stress modulate the susceptibility to Plasmopara viticola in Vitis vinifera self-rooted cuttings. Plants 2021, 10, 273. [Google Scholar] [CrossRef] [PubMed]
- Chrysargyris, A.; Xylia, P.; Antoniou, O.; Tzortzakis, N. Climate change due to heat and drought stress can alter the physiology of Maratheftiko local Cyprian grapevine variety. J. Water Clim. Chang. 2018, 9, 715–727. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Xylia, P.; Litskas, V.; Mandoulaki, A.; Antoniou, D.; Boyias, T.; Stavrinides, M.; Tzortzakis, N. Drought stress and soil management practices in grapevines in Cyprus under the threat of climate change. J. Water Clim. Chang. 2018, 9, 703–714. [Google Scholar] [CrossRef] [Green Version]
- Chrysargyris, A.; Xylia, P.; Litskas, V.; Stavrinides, M.; Heyman, L.; Demeestere, K.; Höfte, M.; Tzortzakis, N. Assessing the impact of drought stress and soil cultivation in Chardonnay and Xynisteri grape cultivars. Agronomy 2020, 10, 670. [Google Scholar] [CrossRef]
- Tzortzakis, N.; Chrysargyris, A.; Aziz, A. Adaptive response of a native Mediterranean grapevine cultivar upon short-term exposure to drought and heat stress in the context of climate change. Agronomy 2020, 10, 249. [Google Scholar] [CrossRef] [Green Version]
- Koufos, G.; Mavromatis, T.; Koundouras, S.; GJones, G. Adaptive capacity of winegrape varieties cultivated in Greece to climate change: Current trends and future projections. OENO One 2020, 54, 1201–1219. [Google Scholar] [CrossRef]
- Phogat, V.; Cox, J.W.; Mallants, D.; Petrie, P.R.; Oliver, D.P.; Pitt, T.R. Historical and future trends in evapotranspiration components and irrigation requirement of winegrapes. Aust. J. Grape Wine Res. 2020, 26, 312–324. [Google Scholar] [CrossRef]
- Nambuthiri, S.; Hagen, E.; Fulcher, A.; Geneve, R. Evaluating a physiological-based, on-demand irrigation system for container-grown woody plants with different water requirements. Hortic. Sci. 2017, 52, 251–257. [Google Scholar] [CrossRef]
- Incrocci, L.; Marzialetti, P.; Incrocci, G.; Di Vita, A.; Balendonck, J.; Bibbiani, C.; Spagnol, S.; Pardossi, A. Substrate water status and evapotranspiration irrigation scheduling in heterogenous container nursery crops. Agric. Water Manag. 2014, 131, 30–40. [Google Scholar] [CrossRef]
- Girona, J.; Mata, M.; del Campo, J.; Arbonés, A.; Bartra, E.; Marsal, J. The use of midday leaf water potential for scheduling deficit irrigation in vineyards. Irrig. Sci. 2006, 24, 115–127. [Google Scholar] [CrossRef]
- Rhie, Y.H.; Kim, J. Changes in physical properties of various coir dust and perlite mixes and their capacitance sensor volumetric water content calibrations. Hortic. Sci. 2017, 52, 162. [Google Scholar] [CrossRef]
- Begg, J.E.; Turner, N.C. Water potential gradients in field tobacco. Plant Physiol. 1970, 46, 343–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meron, M.; Grimes, D.W.; Phene, C.J.; Davis, K.R. Pressure chamber procedures for leaf water potential measurements of cotton. Irrig. Sci. 1987, 8, 215–222. [Google Scholar] [CrossRef]
- Marquard, R.D.; Tipton, J.L. Relationship between extractable chlorophyll and an in situ method to estimate leaf greenness. HortScience 1987, 22, 1327. [Google Scholar]
- Hilu, K.W.; Randall, J.L. Convenient method for studying grass leaf epidermis. Taxon 1984, 33, 413–415. [Google Scholar] [CrossRef]
- Lee, C.J.; O’Donnell, M.; O’Neill, M. Statistical analysis of field trials with changing treatment variance. Agron. J. 2008, 100, 484–489. [Google Scholar] [CrossRef]
- Bureau of Meteorology. Record-Breaking La Niña Events. 2012. Available online: http://www.bom.gov.au/climate/enso/history/ln-2010-12/three-phases-of-ENSO.shtml (accessed on 1 March 2021).
- Bureau of Meteorology. Climate Statistics for Australian Locations. 2021. Available online: http://www.bom.gov.au/climate/averages/tables/cw_023000_All.shtml (accessed on 1 March 2021).
- Liles, C.; Verdon-Kidd, D.C. Refining the growing season temperature parameter for use in winegrape suitability analysis. Aust. J. Grape Wine Res. 2020, 26, 343–357. [Google Scholar] [CrossRef]
- Copper, A.W.; Collins, C.; Bastian, S.; Johnson, T.; Koundouras, S.; Karaolis, C.; Savvides, S. Vine performance benchmarking of indigenous Cypriot grape varieties Xynisteri and Maratheftiko. OENO One 2020, 54, 935–954. [Google Scholar] [CrossRef]
- García-Tejera, O.; López-Bernal, A.; Orgaz, F.; Testi, L.; Villalobos, F. The pitfalls of water potential for irrigation scheduling. Agric. Water Manag. 2021, 243, 1–8. [Google Scholar] [CrossRef]
- Theodorou, N.; Nikolaou, N.; Zioziou, E.; Kyraleou, M.; Kallithraka, S.; Kotseridis, Y.; Koundouras, S. Anthocyanin content and composition in four red winegrape cultivars (Vitis vinifera L.) under variable irrigation. OENO One 2019, 53, 39–51. [Google Scholar] [CrossRef]
- Smart, R.E. Principles of grapevine canopy microclimate manipulation with implications for yield and quality. A review. Am. J. Enol. Vitic. 1985, 36, 230. [Google Scholar]
- Cifre, J.; Bota, J.; Escalona, J.M.; Medrano, H.; Flexas, J. Physiological tools for irrigation scheduling in grapevine (Vitis vinifera L.): An open gate to improve water-use efficiency. Agric. Ecosyst. Environ. 2005, 106, 159–170. [Google Scholar] [CrossRef]
- Steele, M.R.; Gitelson, A.A.; Rundquist, D.C. A comparison of two techniques for nondestructive measurement of chlorophyll content in grapevine leaves. Agron. J. 2008, 100, 779–782. [Google Scholar] [CrossRef] [Green Version]
- Ling, Q.; Huang, W.; Jarvis, P. Use of a SPAD-502 meter to measure leaf chlorophyll concentration in Arabidopsis thaliana. Photosynth. Res. 2011, 107, 209–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogiers, S.; Hardie, W.; Smith, J. Stomatal density of grapevine leaves (Vitis vinifera L.) responds to soil temperature and atmospheric carbon dioxide. Aust. J. Grape Wine Res. 2011, 17, 147–152. [Google Scholar] [CrossRef]
- Boso, S.; Gago, P.; Alonso-Villaverde, V.; Santiago, J.L.; Mendez, J.; Pazos, I.; Martínez, M.C. Variability at the electron microscopic level in leaves of members of the genus Vitis. Sci. Hortic. 2011, 128, 228–238. [Google Scholar] [CrossRef]
- Galat-Giorgi, E.; Keller, M.; Sadras, V.; Roig, F.A.; Perez Peña, J. High temperature during the budswell phase of grapevines increases shoot water transport capacity. Agric. For. Meteorol. 2020, 295, 108–173. [Google Scholar] [CrossRef]
- Allen, L.H.; Zhang, L.; Boote, K.J.; Hauser, B.A. Elevated temperature intensity, timing, and duration of exposure affect soybean internode elongation, mainstem node number, and pod number per plant. Crop J. 2018, 6, 148–161. [Google Scholar] [CrossRef]
- Keller, M.; Tarara, J.M. Warm spring temperatures induce persistent season-long changes in shoot development in grapevines. Ann. Bot. 2010, 106, 131–141. [Google Scholar] [CrossRef] [Green Version]
- Alatzas, A.; Theocharis, S.; Miliordos, D.-E.; Leontaridou, K.; Kanellis, A.K.; Kotseridis, Y.; Hatzopoulos, P.; Koundouras, S. The effect of water deficit on two Greek Vitis vinifera L. cultivars: Physiology, grape composition and gene expression during berry development. Plants 2021, 10, 1947. [Google Scholar] [CrossRef]
- Bauerle, T.L.; Smart, D.R.; Bauerle, W.L.; Stockert, C.; Eissenstat, D.M. Root foraging in response to heterogeneous soil moisture in two grapevines that differ in potential growth rate. New Phytol. 2008, 179, 857–866. [Google Scholar] [CrossRef]
- Gerzon, E.; Biton, I.; Yaniv, Y.; Zemach, H.; Netzer, Y.; Schwartz, A.; Fait, A.; Ben-Ari, G. Grapevine anatomy as a possible determinant of isohydric or anisohydric behavior. Am. J. Enol. Vitic. 2015, 66, 340–347. [Google Scholar] [CrossRef]
- Chaves, M.M.; Zarrouk, O.; Francisco, R.; Costa, J.M.; Santos, T.; Regalado, A.P.; Rodrigues, M.L.; Lopes, C.M. Grapevine under deficit irrigation: Hints from physiological and molecular data. Ann. Bot. 2010, 105, 661–676. [Google Scholar] [CrossRef] [Green Version]
- Levin, A.D.; Williams, L.E.; Matthews, M.A. A continuum of stomatal responses to water deficits among 17 wine grape cultivars. Funct. Plant Biol. 2020, 47, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Hochberg, U.; Rockwell, F.E.; Holbrook, N.M.; Cochard, H. Iso/anisohydry: A plant-environment interaction rather than a simple hydraulic trait. Trends Plant Sci. 2018, 23, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Villalobos-González, L.; Muñoz-Araya, M.; Franck, N.; Pastenes, C. Controversies in midday water potential regulation and stomatal behavior might result from the environment, genotype, and/or rootstock: Evidence from Carménère and Syrah grapevine varieties. Front. Plant Sci. 2019, 10, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dayer, S.; Scharwies, J.D.; Ramesh, S.A.; Sullivan, W.; Doerflinger, F.C.; Pagay, V.; Tyerman, S.D. Comparing hydraulics between two grapevine cultivars reveals differences in stomatal regulation under water stress and exogenous ABA applications. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Liu, Y.; Lu, Y.; Liao, Y.; Nie, J.; Yuan, X.; Chen, F. Use of a leaf chlorophyll content index to improve the prediction of above-ground biomass and productivity. PeerJ-Life Environ. 2019, 6, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Metay, A.; Magnier, J.; Guilpart, N.; Christophe, A. Nitrogen supply controls vegetative growth, biomass and nitrogen allocation for grapevine (cv. Shiraz) grown in pots. Funct. Plant Biol. 2014, 42, 105–114. [Google Scholar] [CrossRef]
- Jumrani, K.; Bhatia, V.S.; Pandey, G.P. Impact of elevated temperatures on specific leaf weight, stomatal density, photosynthesis and chlorophyll fluorescence in soybean. Photosynth. Res. 2017, 131, 333–350. [Google Scholar] [CrossRef]
- Ferlito, F.; Distefano, G.; Gentile, A.; Allegra, M.; Lakso, A.; Nicolosi, E. Scion–rootstock interactions influence the growth and behaviour of the grapevine root system in a heavy clay soil. Aust. J. Grape Wine Res. 2020, 26, 68–78. [Google Scholar] [CrossRef]
- Yıldırım, K.; Yağcı, A.; Sucu, S.; Tunç, S. Responses of grapevine rootstocks to drought through altered root system architecture and root transcriptomic regulations. Plant Physiol. Biochem. 2018, 127, 256–268. [Google Scholar] [CrossRef]
- Gambetta, G.A.; Herrera, J.C.; Dayer, S.; Feng, Q.; Hochberg, U.; Castellarin, S.D. The physiology of drought stress in grapevine: Towards an integrative definition of drought tolerance. J. Exp. Bot. 2020, 71, 4658–4676. [Google Scholar] [CrossRef]
- Alsina, M.M.; Smart, D.R.; Bauerle, T.; de Herralde, F.; Biel, C.; Stockert, C.; Negron, C.; Save, R. Seasonal changes of whole root system conductance by a drought-tolerant grape root system. J. Exp. Bot. 2011, 62, 99–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prinsi, B.; Negri, A.S.; Failla, O.; Scienza, A.; Espen, L. Root proteomic and metabolic analyses reveal specific responses to drought stress in differently tolerant grapevine rootstocks. BMC Plant Biol. 2018, 18, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Prinsi, B.; Simeoni, F.; Galbiati, M.; Meggio, F.; Tonelli, C.; Scienza, A.; Espen, L. Grapevine rootstocks differently affect physiological and molecular responses of the scion under water deficit condition. Agronomy 2021, 11, 289. [Google Scholar] [CrossRef]
- Sharp, R.E.; LeNoble, M.E. ABA, ethylene and the control of shoot and root growth under water stress. J. Exp. Bot. 2002, 53, 33–37. [Google Scholar] [CrossRef]
- Hopper, D.W.; Ghan, R.; Schlauch, K.A.; Cramer, G.A. Transcriptomic network analyses of leaf dehydration responses identify highly connected ABA and ethylene signalling hubs in three grapevine species differing in drought tolerance. BMC Plant Biol. 2016, 16, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.T.; Chen, Z.Y.; Jiang, Y.; Duan, B.B.; Xi, Z.M. Involvement of ABA and antioxidant system in brassinosteroid-induced water stress tolerance of grapevine (Vitis vinifera L.). Sci. Hortic. 2019, 256, 108–596. [Google Scholar] [CrossRef]
- Liu, J.; Zhao, F.L.; Guo, Y.; Fan, X.C.; Wang, Y.J.; Wen, Y.Q. The ABA receptor-like gene VyPYL9 from drought-resistance wild grapevine confers drought tolerance and ABA hypersensitivity in Arabidopsis. Plant Cell Tissue Organ Cult. 2019, 138, 543–558. [Google Scholar] [CrossRef]
- Li, H.; Gao, Z.; Chen, Q.; Li, M.; Luo, J.; Wang, L.; Hu, M.S.; Zahid, L.; Wang, L.; Zhao, S.; et al. Grapevine ABA receptor VvPYL1 regulates root hair development in transgenic Arabidopsis. Plant Physiol. Biochem. 2020, 149, 190–200. [Google Scholar] [CrossRef]
- Vinehealth Australia. Government of South Australia. 2021. Available online: https://vinehealth.com.au/2021/11/rootstock-deep-dive/ (accessed on 14 December 2021).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Start Date | End Date | Total Days | Measurements |
|---|---|---|---|---|
| Cyprus vineyard trial 2019 | 11 June (EL–21) | 26 September (EL–38) | 107 | 5 |
| Cyprus potted trial 2019 | 19 July (EL–33) | 24 September (EL–38) | 67 | 6 |
| Australian potted trial 2020/2021 | 16 December (EL–33) | 27 February (EL–38) | 74 | 7 |
| Treatment | Shoot Number | Shoot Length (cm) | Leaf Number | Shoot Diameter (cm) | Internode Length (cm) | Bunch Length Flower (cm) | Bunch Width Flower (cm) | Bunch Number | Average Bunch Weight (gm) | Yield per Vine (kg) |
|---|---|---|---|---|---|---|---|---|---|---|
| Nil | 28.2 | 163 | 46 | 1.0 | 9.1 | 17.9 | 8.9 | 25.9 | 209 | 5.4 |
| 50% | 25.3 | 146 | 43 | 0.96 | 9.5 | 15.9 | 8.5 | 25.9 | 251 | 6.5 |
| Full | 27.3 | 141 | 42 | 0.96 | 9.5 | 17.6 | 9.4 | 24.1 | 257 | 6.2 |
| p < 0.05 | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| Treatment | ETH | pH | TA | VA | Malic | Fruct | Gluc | Red | FolinC |
|---|---|---|---|---|---|---|---|---|---|
| (g/L) | (g/L) | (g/L) | Acid | (g/L) | (g/L) | Sug | (mg/L) | ||
| (g/L) | (g/L) | ||||||||
| Nil | 0.3 | 3.9 | 1.71 | 0.31 | 1.53 | 109 a | 114 | 206 | 104 |
| 50% | 0.25 | 3.96 | 1.66 | 0.29 | 1.57 | 109 a | 112 | 204 | 108 |
| Full | 0.31 | 3.85 | 1.85 | 0.27 | 1.6 | 103 b | 107 | 194 | 103 |
| Pr > F | ns | ns | ns | ns | ns | 0.04 * | ns | ns | ns |
| Treatment | Shoot Length (cm) | Leaves per Shoot | Shoot Diameter (cm) | Internode Length (cm) |
|---|---|---|---|---|
| 2019 | ||||
| XM | 196 b | 87 a | 0.97 a | 7.2 a |
| XK | 2358 a | 105 a | 1.03 a | 7.5 a |
| SBC | 118 c | 98 a | 0.45 b | 5.9 b |
| Pr > F | <0.0001 | 0.079 | <0.0001 | <0.0001 |
| 2020/2021 | ||||
| XP | 152 a | 44 ab | 1.03 a | 10.4 b |
| MP | 166 a | 36 b | 1.09 a | 12.6 a |
| SZ | 171 a | 47 a | 0.75 b | 11.9 ab |
| SBA | 101 b | 42 ab | 0.74 b | 8.01 c |
| Pr > F | <0.0001 | 0.004 | <0.0001 | <0.0001 |
| Factor | F Value | p-Value |
|---|---|---|
| 2019 | ||
| Stem water potential | ||
| Time | 104.376 | <0.0001 * |
| Irrigation | 51.823 | 0.0002 * |
| Time × Irrigation | 4.484 | 0.001 * |
| Stomatal conductance | ||
| Time | 118.548 | <0.0001 * |
| Irrigation | 27.634 | 0.001 * |
| Time × Irrigation | 5.131 | 0.0002 * |
| SPAD reading | ||
| Time | 1.286 | 0.296 |
| Irrigation | 0.234 | 0.799 |
| Time × Irrigation | 1.976 | 0.073 |
| 2020/2021 | ||
| Stem water potential | ||
| Time | 119.446 | <0.0001 * |
| Irrigation | 15.992 | 0.001 * |
| Time × Irrigation | 3.731 | 0.0004 * |
| Stomatal conductance | ||
| Time | 90.098 | <0.0001 * |
| Irrigation | 2.065 | 0.183 |
| Time × Irrigation | 1.367 | 0.210 |
| SPAD reading | ||
| Time | 16.054 | <0.0001 * |
| Irrigation | 0.133 | 0.877 |
| Time × Irrigation | 0.127 | 1.000 |
| Mass (gm) | Root | Shoot | Leaf | ||||||
|---|---|---|---|---|---|---|---|---|---|
| IR | Full | 50% | 25% | Full | 50% | 25% | Full | 50% | 25% |
| Cyprus 2019 | |||||||||
| XM | 693 b | 582 ab | 387 ab | 264 b | 204 a | 112 b | 243 b | 184 a | 102 ab |
| XK | 939 a | 643 a | 486 a | 377 a | 234 a | 180 a | 359 a | 208 a | 156 a |
| SBC | 493 c | 352 b | 182 b | 109 c | 93 b | 63 c | 129 c | 93 b | 48 b |
| Pr > F | <0.0001 | 0.01 | <0.0001 | <0.0001 | 0.0001 | 0.0002 | <0.0001 | 0.0017 | 0.0001 |
| Adelaide 20/21 | |||||||||
| XP | 1233 a | 1135 a | 892 a | 458 a | 411 a | 342 a | 357 a | 291 a | 259 a |
| MP | 620 b | 567 b | 539 b | 425 ab | 366 ab | 296 ab | 252 ab | 240 ab | 201 ab |
| SZ | 592 b | 445 b | 320 c | 299 b | 286 b | 238 ab | 215 b | 154 c | 137 b |
| SBA | 610 b | 494 b | 443 b | 307 b | 274 b | 206 b | 236 b | 205 bc | 140 b |
| Pr > F | 0.0004 | <0.0001 | <0.0001 | 0.009 | 0.035 | 0.029 | 0.011 | 0.0003 | 0.001 |
| Season | Variety | Stomatal Density |
|---|---|---|
| 2019 | XCV | 238.6 a |
| XK | 227.5 a | |
| XM | 233.2 a | |
| SBC | 139.8 b | |
| Pr > F | <0.0001 | |
| 2020/2021 | XP | 206.1 a |
| MP | 189.0 b | |
| SZ | 170.5 c | |
| SBA | 151.4 d | |
| Pr > F | <0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Copper, A.W.; Koundouras, S.; Bastian, S.E.P.; Johnson, T.E.; Collins, C. Assessing the Response of Vitis vinifera L. cv. Xynisteri to Different Irrigation Regimes and Its Comparison to cvs. Maratheftiko, Shiraz and Sauvignon Blanc. Agronomy 2022, 12, 634. https://doi.org/10.3390/agronomy12030634
Copper AW, Koundouras S, Bastian SEP, Johnson TE, Collins C. Assessing the Response of Vitis vinifera L. cv. Xynisteri to Different Irrigation Regimes and Its Comparison to cvs. Maratheftiko, Shiraz and Sauvignon Blanc. Agronomy. 2022; 12(3):634. https://doi.org/10.3390/agronomy12030634
Chicago/Turabian StyleCopper, Alexander W., Stefanos Koundouras, Susan E. P. Bastian, Trent E. Johnson, and Cassandra Collins. 2022. "Assessing the Response of Vitis vinifera L. cv. Xynisteri to Different Irrigation Regimes and Its Comparison to cvs. Maratheftiko, Shiraz and Sauvignon Blanc" Agronomy 12, no. 3: 634. https://doi.org/10.3390/agronomy12030634
APA StyleCopper, A. W., Koundouras, S., Bastian, S. E. P., Johnson, T. E., & Collins, C. (2022). Assessing the Response of Vitis vinifera L. cv. Xynisteri to Different Irrigation Regimes and Its Comparison to cvs. Maratheftiko, Shiraz and Sauvignon Blanc. Agronomy, 12(3), 634. https://doi.org/10.3390/agronomy12030634