
Utilization of Vertical Cordon System to Improve Source-Sink Balance and Wine Aroma under Water Shortage Conditions of Maturana Blanca

 ,
,  , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Irrigation Treatments
2.2. Grapevine Water Status
2.3. Vegetative Growth Determinations
2.4. Yield Components
2.5. Fruit Composition Analysis
2.6. Winemaking
2.7. Analysis of Wine Volatile Composition
2.8. Statistical Analysis
3. Results
3.1. Climate and Irrigation Treatments
3.2. Differences in Vegetative Growth and Leaf Area
3.3. Vine Water Status
3.4. Yield Components
3.5. Must Quality
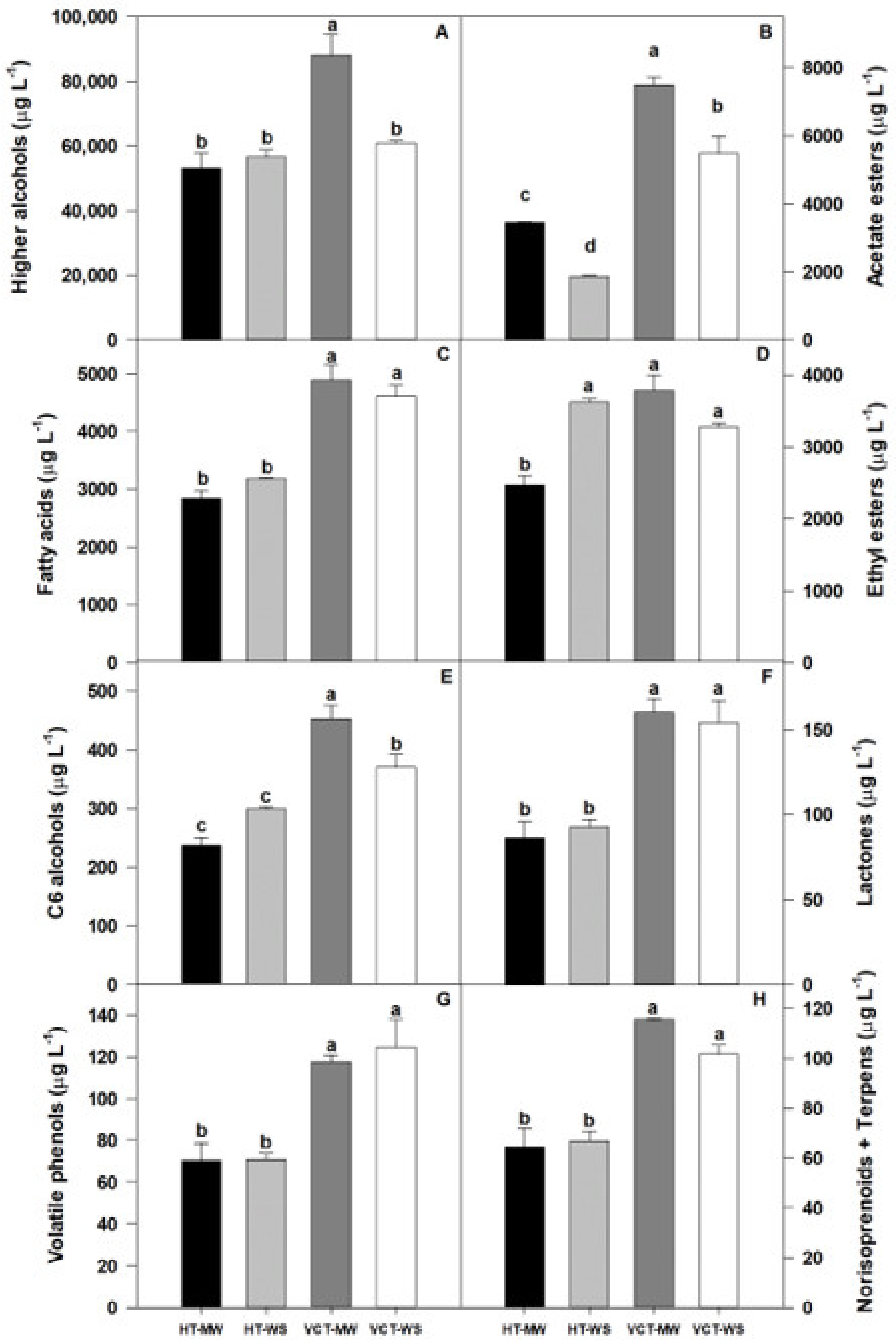
3.6. Wine Volatile Composition
3.7. Principal Component Analysis (PCA)
4. Discussion
4.1. Vegetative Development
4.2. Crop Yield
4.3. Must Composition
4.4. Wine Aroma
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dry, P.R.; Loveys, B.R. Factors influencing grapevine vigour and the potential for control with partial rootzone drying. Aust. J. Grape Wine Res. 1998, 4, 140–148. [Google Scholar] [CrossRef]
- Smart, R.E. Principles of Grapevine Canopy Microclimate Manipulation with Implications for Yield and Quality. A review. Am. J. Enol. Vitic. 1985, 36, 230–239. [Google Scholar]
- Kliewer, W.M.; Dokoozlian, N.K. Leaf area/crop weight ratios of grapevines: Influence on fruit composition and wine quality. Am. J. Enol. Vitic. 2005, 56, 170–181. [Google Scholar]
- Bramley, R.G.V.; Ouzman, J.; Boss, P.K. Variation in vine vigour, grape yield and vineyard soils and topography as indicators of variation in the chemical composition of grapes, wine and wine sensory attributes. Aust. J. Grape Wine Res. 2011, 17, 217–229. [Google Scholar] [CrossRef]
- Chaves, M.M.; Santos, T.P.; Souza, C.R.; Ortuño, M.F.; Rodrigues, M.L.; Lopes, C.M.; Maroco, J.P.; Pereira, J.S. Deficit irrigation in grapevine improves water-use efficiency while controlling vigour and production quality. Ann. Appl. Biol. 2007, 150, 237–252. [Google Scholar] [CrossRef]
- Jackson, D.I.; Lombard, P.B.; Kabinett, L.Q. Environmental and management practices affecting grape composition and wine quality—A review. Am. J. Enol. Vitic. 1993, 44, 409–430. [Google Scholar]
- Chen, K.; Li, J. A glance into the aroma of white wine. In White Wine Technology; Morata, A., Ed.; Academic Press: Cambridge, MA, USA, 2022; pp. 313–326. [Google Scholar] [CrossRef]
- Silva Ferreira, A.C.; Monteiro, J.; Oliveira, C.; Guedes de Pinho, P. Study of major aromatic compounds in port wines from carotenoid degradation. Food Chem. 2008, 110, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Martínez de Toda, F.; Sancha, J.C. Ampelographical characterization of white Vitis vinifera L. cultivars preserved in Rioja. Bull. l’OIV 1997, 70, 688–702. [Google Scholar]
- Cervera, M.T.; Cabezas, J.A.; Sancha, J.C.; Martínez de Toda, F.; Martínez-Zapater, J.M. Application of AFLPS to the characterization of grapevine Vitis vinifera L. genetic resources. A case study with accessions from Rioja (Spain). Theor. Appl. Genet. 1998, 97, 51–59. [Google Scholar] [CrossRef] [Green Version]
- Martínez De Toda, F.; Balda, P.; Sancha, J.C. Preservation of intravarietal diversity in clonal and sanitary pre-selection for a minority variety in danger of extinction: Maturana blanca. J. Int. Sci. Vigne Vin 2012, 46, 123–130. [Google Scholar] [CrossRef]
- Balda, P.; Martínez de Toda, F. Variedades Minoritarias de Vid en la DOCa Rioja; Consejería de Agricultura, Ganadería y Medio Ambiente: Logroño, Spain, 2017; ISBN 978-84-8125-686-4.
- Naranjo, A.; Martínez-Lapuente, L.; Ayestarán, B.; Guadalupe, Z.; Pérez, I.; Canals, C.; Adell, E. Aromatic and Sensory Characterization of Maturana Blanca Wines Made with Different Technologies. Beverages 2021, 7, 10. [Google Scholar] [CrossRef]
- Martínez, J.; López, E.; Baroja, E. Evaluación agronómica y enológica de la variedad Tempranillo blanco (Vitis vinifera L.) y de otras variedades minoritarias blancas. In Jornadas del Grupo Viticultura y Enologia de la SECH-Retos Actuales I+D en Viticultura; García-Escudero, E., Martín, I., Eds.; SECH: Logroño, Spain, 2014; pp. 27–33. ISBN 978-84-8125-675. [Google Scholar]
- Mawdsley, P.F.W.; Dodson Peterson, J.C.; Casassa, L.F. Multi-year study of the effects of cluster thinning on vine performance, fruit and wine composition of Pinot noir (clone 115) in California’s Edna Valley AVA (USA). Sci. Hortic. 2019, 256, 108631. [Google Scholar] [CrossRef]
- Gambetta, J.M.; Bastian, S.E.P.; Cozzolino, D.; Jeffery, D.W. Factors influencing the aroma composition of Chardonnay wines. J. Agric. Food Chem. 2014, 62, 6512–6534. [Google Scholar] [CrossRef] [PubMed]
- Guilpart, N.; Metay, A.; Gary, C. Grapevine bud fertility and number of berries per bunch are determined by water and nitrogen stress around flowering in the previous year. Eur. J. Agron. 2014, 54, 9–20. [Google Scholar] [CrossRef]
- Reynolds, A.G.; Vanden Heuvel, J.E. Influence of grapevine training systems on vine growth and fruit composition: A review. Am. J. Enol. Vitic. 2009, 60, 251–268. [Google Scholar]
- Zurowietz, A.; Lehr, P.P.; Kleb, M.; Merkt, N.; Gödde, V.; Bednarz, H.; Niehaus, K.; Zörb, C. Training grapevines generates a metabolomic signature of wine. Food Chem. 2022, 368, 130665. [Google Scholar] [CrossRef] [PubMed]
- Yuste, J. Podas de formación y producción en distintos sistemas de conducción. Alternativas de poda en cordón vertical, vaso y espaldera. In Proceedings of the Ponencias del II Curso de Viticultura y Enología en la Ribera del Duero, Aranda de Duero, Spain, 9–13 July 2002; pp. 11–21. [Google Scholar]
- Vanden Heuvel, J.E.; Proctor, J.T.A.; Sullivan, J.A.; Fisher, K.H. Influence of training/trellising system and rootstock selection on productivity and fruit composition of Chardonnay and Cabernet franc grapevines in Ontario, Canada. Am. J. Enol. Vitic. 2004, 55, 253–264. [Google Scholar]
- Williams, L.E.; Phene, C.J.; Grimes, D.W.; Trout, T.J. Water use of mature Thompson Seedless grapevines in California. Irrig. Sci. 2003, 22, 11–18. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2022. Impacts, Adaptation and Vulnerability. Summary for Policymakers. Contribution for Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. Available online: https://www.ipcc.ch/ (accessed on 3 March 2022).
- Van Leeuwen, C.; Darriet, P. The Impact of Climate Change on Viticulture and Wine Quality. J. Wine Econ. 2016, 11, 150–167. [Google Scholar] [CrossRef] [Green Version]
- Zheng, W. Viticultural Techniques of Canopy Management to Mitigate the Effects of Global Warming; Universidad de La Rioja: Logrono, Spain, 2017. [Google Scholar]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration: Guidelines for Computing Crop Water Requirements; Food and Agriculture Organization of the United Nations: Rome, Italy, 1998. [Google Scholar]
- Eichhorn, K.W.; Lorenz, D.H. Phänologische Entwicklungsstadien der Rebe. Nachrichtenbl. Dt. Pflanzenschutzd. 1977, 29, 119–120. [Google Scholar]
- Sanchez-de-Miguel, P.; Junquera, P.; De la Fuente, M.; Jimenez, L.; Linares, R.; Baeza, P.; Lissarrague, J.R. Estimation of vineyard leaf area by linear regression. Span. J. Agric. Res. 2011, 9, 202. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-de-Miguel, P.; Baeza, P.; Junquera, P.; Lissarrague, J.R. Vegetative Development: Total Leaf Area and Surface Area Indexes. In Methodologies and Results in Grapevine Research; Delrot, S., Medrano, H., Or, E., Bavaresco, L., Grando, S., Eds.; Springer Netherlands: Dordrecht, The Netherlands, 2010; pp. 31–44. ISBN 978-90-481-9283-0. [Google Scholar]
- OIV. Compendium of International Methods of Wine and Must Analysis; OIV: Paris, France, 2021; ISBN 9782850380167. [Google Scholar]
- Lipka, Z.; Tanner, H. Une nouvelle méthode de dosage du acide tartarique dans les moûts, les vines et autres boissons (selon Rebelein). Rev. Suisse Vitic. Arboric. Hortic. 1974, 6, 5–10. [Google Scholar]
- Aerny, J. Composes azotes des mouts et des vins. Rev. Suisse Vitic. Arboric. Hortic. 1996, 28, 161–165. [Google Scholar]
- Ribereau-Gayon, P.; Glories, Y.; Maujean, A.; Dubourdieu, D. Phenolic compound. In Handbook of Enology. The Chemistry of Wine Stabilization and Treatments; Wiley: West Sessex, UK, 2000; pp. 129–187. [Google Scholar]
- Coelho, E.; Lemos, M.; Genisheva, Z.; Domingues, L.; Vilanova, M.; Oliveira, J.M. Validation of a LLME/GC-MS methodology for quantification of volatile compounds in fermented beverages. Molecules 2020, 25, 621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, X.; Jia, F.; Cai, J.; Shi, Y.; Duan, C.; Lan, Y. Characterization and evolution of volatile compounds of cabernet sauvignon wines from two different clones during oak barrel aging. Foods 2022, 11, 74. [Google Scholar] [CrossRef]
- Cai, J.; Zhu, B.Q.; Wang, Y.H.; Lu, L.; Lan, Y.B.; Reeves, M.J.; Duan, C.Q. Influence of pre-fermentation cold maceration treatment on aroma compounds of Cabernet Sauvignon wines fermented in different industrial scale fermenters. Food Chem. 2014, 154, 217–229. [Google Scholar] [CrossRef] [PubMed]
- Albanese, D.; Attanasio, G.; Cinquanta, L.; Di Matteo, M. Volatile Compounds in Red Wines Processed on an Industrial Scale by Short Pre-fermentative Cold Maceration. Food Bioprocess Technol. 2013, 6, 3266–3272. [Google Scholar] [CrossRef]
- Francis, I.L.; Newton, J.L. Determining wine aroma from compositional data. Aust. J. Grape Wine Res. 2005, 11, 114–126. [Google Scholar] [CrossRef]
- Etiévant, P.X. Wine. In Volatile Compounds in Food and Beverages; CRC Press: Boca Raton, FL, USA, 1991; pp. 483–546. [Google Scholar]
- Ferreira, V.; López, R.; Cacho, J.F. Quantitative determination of the odorants of young red wines from different grape varieties. J. Sci. Food Agric. 2000, 80, 1659–1667. [Google Scholar] [CrossRef]
- Sánchez-Palomo, E.; Gómez García-Carpintero, E.; Gómez Gallego, M.A.; González Viñas, M.A. The aroma of Rojal red wines from La Mancha region-determination of key odorants. In Gas Chromatography in Plant Science, Wine Technology, Toxicology and Some Specific Application; InTech: Rijeka, Croatia, 2012; pp. 147–170. [Google Scholar]
- Wang, X.C.; Li, A.H.; Dizy, M.; Ullah, N.; Sun, W.X.; Tao, Y.S. Evaluation of aroma enhancement for “Ecolly” dry white wines by mixed inoculation of selected Rhodotorula mucilaginosa and Saccharomyces cerevisiae. Food Chem. 2017, 228, 550–559. [Google Scholar] [CrossRef]
- Vilanova, M.; Campo, E.; Escudero, A.; Graña, M.; Masa, A.; Cacho, J. Volatile composition and sensory properties of Vitis vinifera red cultivars from North West Spain: Correlation between sensory and instrumental analysis. Anal. Chim. Acta 2012, 720, 104–111. [Google Scholar] [CrossRef]
- Guth, H. Quantitation and Sensory Studies of Character Impact Odorants of Different White Wine Varieties. J. Agric. Food Chem. 1997, 45, 3027–3032. [Google Scholar] [CrossRef]
- Culleré, L.; Escudero, A.; Cacho, J.; Ferreira, V. Gas Chromatography-Olfactometry and Chemical Quantitative Study of the Aroma of Six Premium Quality Spanish Aged Red Wines. J. Agric. Food Chem. 2004, 52, 1653–1660. [Google Scholar] [CrossRef]
- Peinado, R.A.; Mauricio, J.C.; Moreno, J. Aromatic series in sherry wines with gluconic acid subjected to different biological aging conditions by Saccharomyces cerevisiae var. capensis. Food Chem. 2006, 94, 232–239. [Google Scholar] [CrossRef]
- Niu, Y.; Wang, P.; Xiao, Q.; Xiao, Z.; Mao, H.; Zhang, J. Characterization of odor-active volatiles and odor contribution based on binary interaction effects in mango and vodka cocktail. Molecules 2020, 25, 1083. [Google Scholar] [CrossRef] [Green Version]
- Pineau, B.; Barbe, J.C.; Van Leeuwen, C.; Dubourdieu, D. Which impact for β-damascenone on red wines aroma? J. Agric. Food Chem. 2007, 55, 4103–4108. [Google Scholar] [CrossRef]
- Vilanova, M.; Genisheva, Z.; Bescansa, L.; Masa, A.; Oliveira, J.M. Volatile composition of wines from cvs. Blanco lexítimo, Agudelo and Serradelo (Vitis vinifera) grown in Betanzos (NW Spain). J. Inst. Brew. 2009, 115, 35–40. [Google Scholar] [CrossRef] [Green Version]
- de-la-Fuente-Blanco, A.; Sáenz-Navajas, M.P.; Valentin, D.; Ferreira, V. Fourteen ethyl esters of wine can be replaced by simpler ester vectors without compromising quality but at the expense of increasing aroma concentration. Food Chem. 2020, 307, 125553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, P.; Gao, J.; Qian, M.; Li, H. Characterization of the key aroma compounds in Chinese syrah wine by gas chromatography-Olfactometry-Mass spectrometry and Aroma reconstitution studies. Molecules 2017, 22, 1045. [Google Scholar] [CrossRef]
- Noguerol-Pato, R.; González-Barreiro, C.; Cancho-Grande, B.; Santiago, J.L.; Martínez, M.C.; Simal-Gándara, J. Aroma potential of Brancellao grapes from different cluster positions. Food Chem. 2012, 132, 112–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Navarro, J.; Izquierdo-Cañas, P.M.; Mena-Morales, A.; Chacón-Vozmediano, J.L.; Martínez-Gascueña, J.; García-Romero, E.; Hermosín-Gutiérrez, I.; Gómez-Alonso, S. Comprehensive chemical and sensory assessment of wines made from white grapes of Vitis vinifera cultivars albillo dorado and montonera del casar: A comparative study with airén. Foods 2020, 9, 1282. [Google Scholar] [CrossRef] [PubMed]
- Aznar, M.; López, R.; Cacho, J.; Ferreira, V. Prediction of aged red wine aroma properties from aroma chemical composition. Partial least squares regression models. J. Agric. Food Chem. 2003, 51, 2700–2707. [Google Scholar] [CrossRef]
- Bernardo, S.; Dinis, L.T.; Machado, N.; Moutinho-Pereira, J. Grapevine abiotic stress assessment and search for sustainable adaptation strategies in Mediterranean-like climates. A review. Agron. Sustain. Dev. 2018, 38, 66. [Google Scholar] [CrossRef] [Green Version]
- Favero, A.C.; de Amorim, D.A.; da Mota, R.V.; de Souza, C.R.; Regina, M.D.A. Physiological responses and production of “Syrah” vines as a function of training systems. Sci. Agric. 2010, 67, 267–273. [Google Scholar] [CrossRef] [Green Version]
- Keller, M.; Smithyman, R.P.; Mills, L.J. Interactive effects of deficit irrigation and crop load on cabernet sauvignon in an arid climate. Am. J. Enol. Vitic. 2008, 59, 221–234. [Google Scholar]
- Keller, M.; Mills, L.J.; Wample, R.L.; Spayd, S.E. Crop load management in concord grapes using different pruning techniques. Am. J. Enol. Vitic. 2004, 55, 35–50. [Google Scholar]
- Martinez De Toda, F. Claves de la Viticultura de Calidad; Mundi-Prensa: Madrid, Spain, 2011. [Google Scholar]
- Clingeleffer, P.R. Update: Minimal pruning of cordon trained vines (MPCT). Austr. Grape Grow. Winemak. 1989, 304, 78–83. [Google Scholar]
- Smart, R.E.; Dick, J.K.; Gravett, I.M.; Fisher, B.M. Canopy Management to Improve Grape Yield and Wine Quality-Principles and Practices. S. Afr. J. Enol. Vitic. 1990, 11, 3–17. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, A.G.; Wardle, D.A. Effects of Timing and Severity of Summer Hedging on Growth, Yield, Fruit Composition, and Canopy Characteristics of de Chaunac. I. Canopy Characteristics and Growth Parameters. Am. J. Enol. Vitic. 1989, 40, 109–120. [Google Scholar]
- Del Campo, M.G.; Ruiz, C.; Sotes, V.; Lissarrague, J.R. Water consumption in grapevines: Influence of leaf area and irrigation. Acta Hortic. 1999, 493, 279–286. [Google Scholar] [CrossRef]
- Ojeda, H.; Deloire, A.; Carbonneau, A. Influence of water deficits on grape berry growth. Vitis 2001, 40, 141–145. [Google Scholar]
- Musacchi, S.; Iglesias, I.; Neri, D. Training systems and sustainable orchard management for european pear (Pyrus communis L.) in the mediterranean area: A review. Agronomy 2021, 11, 1765. [Google Scholar] [CrossRef]
- Esteban, M.A.; Villanueva, M.J.; Lissarrague, J.R. Effect of irrigation on changes in berry composition of Tempranillo during maturation. Sugars, organic acids, and mineral elements. Am. J. Enol. Vitic. 1999, 50, 418–434. [Google Scholar]
- Koundouras, S.; Hatzidimitriou, E.; Karamolegkou, M.; Dimopoulou, E.; Kallithraka, S.; Tsialtas, J.T.; Zioziou, E.; Nikolaou, N.; Kotseridis, Y. Irrigation and rootstock effects on the phenolic concentration and aroma potential of Vitis vinifera L. cv. Cabernet Sauvignon grapes. J. Agric. Food Chem. 2009, 57, 7805–7813. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, T.P.; Lopes, C.M.; Lucília Rodrigues, M.; de Souza, C.R.; Ricardo-da-Silva, J.M.; Maroco, J.P.; Pereira, J.S.; Manuela Chaves, M. Effects of deficit irrigation strategies on cluster microclimate for improving fruit composition of Moscatel field-grown grapevines. Sci. Hortic. 2007, 112, 321–330. [Google Scholar] [CrossRef]
- Wang, X.; Lesefko, S.; de Bei, R.; Fuentes, S.; Collins, C. Effects of canopy management practices on grapevine bud fruitfulness. Oeno One 2020, 54, 313–325. [Google Scholar] [CrossRef]
- Keller, M. The Science of Grapevines; Academic Press: Cambridge, MA, USA, 2020. [Google Scholar]
- Martínez De Toda, F.; Balda, P. Delaying berry ripening through manipulating leaf area to fruit ratio. Vitis-J. Grapevine Res. 2013, 52, 171–176. [Google Scholar]
- Trought, M.C.T.; Parker, A.K.; Van Leeuwen, C. Can a change in vineyard practice mitigate warming due to climate change? Acta Hortic. 2015, 1082, 397–402. [Google Scholar] [CrossRef]
- Valentini, G.; Allegro, G.; Pastore, C.; Colucci, E.; Filippetti, I. Post-veraison trimming slow down sugar accumulation without modifying phenolic ripening in Sangiovese vines. J. Sci. Food Agric. 2019, 99, 1358–1365. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Wu, X.; Needs, S.; Liu, D.; Fuentes, S.; Howell, K. The influence of apical and basal defoliation on the canopy structure and biochemical composition of Vitis vinifera cv. shiraz grapes and wine. Front. Chem. 2017, 5, 1–9. [Google Scholar] [CrossRef] [Green Version]
- O’brien, P.; Collins, C.; De Bei, R. Leaf removal applied to a sprawling canopy to regulate fruit ripening in cabernet sauvignon. Plants 2021, 10, 1017. [Google Scholar] [CrossRef]
- Bravdo, B.; Hepner, Y.; Loinger, C.; Cohen, S.; Tabacman, H. Effect of crop level on growth, yield and wine quality of a high yielding carignane vineyard. Am. J. Enol. Vitic. 1984, 35, 247–252. [Google Scholar]
- Bravdo, B.; Hepner, Y.; Loinger, C.; Cohen, S.; Tabacman, H. Effect of irrigation and crop level on growth, yield and wine quality of Cabernet Sauvignon. Am. J. Enol. Vitic. 1985, 36, 132–139. [Google Scholar]
- Smart, R.E.; Robinson, M. Sunlight into Wine: A Handbook for Winegrape Canopy Management; Winetitles: Underdale, Australia, 1991. [Google Scholar]
- Sweetman, C.; Sadras, V.O.; Hancock, R.D.; Soole, K.L.; Ford, C.M. Metabolic effects of elevated temperature on organic acid degradation in ripening Vitis vinifera fruit. J. Exp. Bot. 2014, 65, 5975–5988. [Google Scholar] [CrossRef] [PubMed]
- Intrigliolo, D.S.; Castel, J.R. Effects of irrigation on the performance of grapevine cv. Tempranillo in Requena, Spain. Am. J. Enol. Vitic. 2008, 59, 30–38. [Google Scholar]
- Martínez de Toda Fernández, F.; García Martín, J.; Balda Manzanos, P. Adaptación al calentamiento climático de veinte variedades de vid, minoritarias de la DOCa Rioja, por su potencial de acidez. Zubía 2017, 29, 83–94. [Google Scholar]
- Hale, C.R. Relation between potassium and the malate and tartrate contents of grape berries. Vitis 1977, 16, 9–19. [Google Scholar]
- Mpelasoka, B.S.; Schachtman, D.P.; Treeby, M.T.; Thomas, M.R. A review of potassium nutrition in grapevines with special emphasis on berry accumulation. Aust. J. Grape Wine Res. 2003, 9, 154–168. [Google Scholar] [CrossRef]
- Kodur, S. Effects of juice pH and potassium on juice and wine quality, and regulation of potassium in grapevines through rootstocks (Vitis): A short review. Vitis-J. Grapevine Res. 2011, 50, 1–6. [Google Scholar]
- Boulton, R. The general relationship between potassium, sodium and pH in grape juice and wine. Am. J. Enol. Vitic. 1980, 31, 182–186. [Google Scholar]
- Rojas-Lara, B.A.; Morrison, J.C. Differential effects of shading fruit or foliage on the development and composition of grape berries. Vitis 1989, 28, 199–208. [Google Scholar]
- Dokoozlian, N.K.; Kliewer, W.M. Influence of Light on Grape Berry Growth andComposition Varies during Fruit Development. J. Am. Soc. Hortic. Sci. 1996, 121, 869–874. [Google Scholar] [CrossRef]
- Kyraleou, M.; Kallithraka, S.; Koundouras, S.; Chira, K.; Haroutounian, S.; Spinthiropoulou, H.; Kotseridis, Y. Effect of vine training system on the phenolic composition of red grapes (Vitis vinifera L. cv. Xinomavro). J. Int. Sci. Vigne Vin 2015, 49, 71–84. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Smart, R.; Wang, H.; Dambergs, B.; Sparrow, A.; Qian, M.C. Effect of grape bunch sunlight exposure and UV radiation on phenolics and volatile composition of Vitis vinifera L. cv. Pinot noir wine. Food Chem. 2015, 173, 424–431. [Google Scholar] [CrossRef] [PubMed]
- Salón, J.L.; Chirivella, C.; Castel, J.R. Response of cv. Bobal to timing of deficit irrigation in Requena, Spain: Water relations, yield, and wine quality. Am. J. Enol. Vitic. 2005, 56, 1–8. [Google Scholar]
- Hardie, W.J.; O’Brien, T.P.; Jaudzems, V.G. Cell biology of grape secondary metabolism: A viticultural perspective. In Proceedings of the Ninth Australian Wine Industry Technical Conference, Adelaide, Australia, 16–19 July 1995; Australian Wine Industry Technical Conference Inc.: Adelaide, Australia, 1997; pp. 78–82. [Google Scholar]
- Swiegers, J.H.; Bartowsky, E.J.; Henschke, P.A.; Pretorius, I.S. Yeast and bacterial modulation of wine aroma and flavour. Aust. J. Grape Wine Res. 2005, 11, 139–173. [Google Scholar] [CrossRef]
- Ayestarán, B.; Martínez-Lapuente, L.; Guadalupe, Z.; Canals, C.; Adell, E.; Vilanova, M. Effect of the winemaking process on the volatile composition and aromatic profile of Tempranillo Blanco wines. Food Chem. 2019, 276, 187–194. [Google Scholar] [CrossRef]
- Oliveira, J.M.; Faria, M.; Sá, F.; Barros, F.; Araújo, I.M. C6-alcohols as varietal markers for assessment of wine origin. Anal. Chim. Acta 2006, 563, 300–309. [Google Scholar] [CrossRef] [Green Version]
- Buesa, I.; Intrigliolo, D.S.; Castel, J.R.; Vilanova, M. Influence of water regime on grape aromatic composition of Muscat of Alexandria in a semiarid climate. Sci. Hortic. 2021, 290, 110525. [Google Scholar] [CrossRef]
- Belancic, A.; Agosin, E.; Ibacache, A.; Bordeu, E.; Baumes, R.; Razungles, A.; Bayonove, C. Influence of sun exposure on the aromatic composition of chilean Muscat grape cultivars Moscatel de Alejandria and Moscatel rosada. Am. J. Enol. Vitic. 1997, 48, 181–186. [Google Scholar]
- Wang, J.; Abbey, T.; Kozak, B.; Madilao, L.L.; Tindjau, R.; Del Nin, J.; Castellarin, S.D. Evolution over the growing season of volatile organic compounds in Viognier (Vitis vinifera L.) grapes under three irrigation regimes. Food Res. Int. 2019, 125, 108512. [Google Scholar] [CrossRef]
- Talaverano, I.; Valdés, E.; Moreno, D.; Gamero, E.; Mancha, L.; Vilanova, M. The combined effect of water status and crop level on Tempranillo wine volatiles. J. Sci. Food Agric. 2017, 97, 1533–1542. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Shellie, K.C.; Wang, H.; Qian, M.C. Influence of deficit irrigation and kaolin particle film on grape composition and volatile compounds in Merlot grape (Vitis vinifera L.). Food Chem. 2012, 134, 841–850. [Google Scholar] [CrossRef]
- Mendez-Costabel, M.P.; Wilkinson, K.L.; Bastian, S.E.P.; Jordans, C.; Mccarthy, M.; Ford, C.M.; Dokoozlian, N.K. Effect of increased irrigation and additional nitrogen fertilisation on the concentration of green aroma compounds in Vitis vinifera L. Merlot fruit and wine. Aust. J. Grape Wine Res. 2014, 20, 80–90. [Google Scholar] [CrossRef]
- López, E.F.; Gómez, E.F. Comparison of solvents for determination of monoterpenes in wine using liquid-liquid extraction. Chromatographia 2000, 52, 798–802. [Google Scholar] [CrossRef]
- Reynolds, A.G.; Wardle, D. Influence of fruit microclimate on monoterpene levels of Gewurztraminer. Am. J. Enol. Vitic. 1989, 40, 149–154. [Google Scholar]
- Friedel, M.; Frotscher, J.; Nitsch, M.; Hofmann, M.; Bogs, J.; Stoll, M.; Dietrich, H. Light promotes expression of monoterpene and flavonol metabolic genes and enhances flavour of winegrape berries (Vitis vinifera L. cv. Riesling). Aust. J. Grape Wine Res. 2016, 22, 409–421. [Google Scholar] [CrossRef]
- Feng, H.; Skinkis, P.A.; Qian, M.C. Pinot noir wine volatile and anthocyanin composition under different levels of vine fruit zone leaf removal. Food Chem. 2017, 214, 736–744. [Google Scholar] [CrossRef] [PubMed]
- Young, P.R.; Eyeghe-Bickong, H.A.; du Plessis, K.; Alexandersson, E.; Jacobson, D.A.; Coetzee, Z.; Deloire, A.; Vivier, M.A. Grapevine plasticity in response to an altered microclimate: Sauvignon Blanc modulates specific metabolites in response to increased berry exposure. Plant Physiol. 2016, 170, 1235–1254. [Google Scholar] [CrossRef] [Green Version]
- Bowen, A.J.; Reynolds, A.G. Aroma compounds in Ontario Vidal and Riesling icewines. II. Effects of crop level. Food Res. Int. 2015, 76, 550–560. [Google Scholar] [CrossRef] [PubMed]
- Rutan, T.E.; Herbst-Johnstone, M.; Kilmartin, P.A. Effect of Cluster Thinning Vitis vinifera cv. Pinot Noir on Wine Volatile and Phenolic Composition. J. Agric. Food Chem. 2018, 66, 10053–10066. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, A.G.; Parchomchuk, P.; Berard, R.; Naylor, A.P.; Hogue, E. Gewurztraminer grapevines respond to length of water stress duration. Int. J. Fruit Sci. 2005, 5, 75–94. [Google Scholar] [CrossRef]
- Giordano, M.; Zecca, O.; Belviso, S.; Reinotti, M.; Gerbi, V.; Rolle, L. Volatile fingerprint and physico-mechanical properties of ‘Muscat blanc’ grapes grown in mountain area: A first evidence of the influence of water regimes. Ital. J. Food Sci. 2013, 25, 329–338. [Google Scholar]
- Des Gachons, C.P.; Van Leeuwen, C.; Tominaga, T.; Soyer, J.P.; Gaudillère, J.P.; Dubourdieu, D. Influence of water and nitrogen deficit on fruit ripening and aroma potential of Vitis vinifera L. cv Sauvignon blanc in field conditions. J. Sci. Food Agric. 2005, 85, 73–85. [Google Scholar] [CrossRef]
- Sumby, K.M.; Grbin, P.R.; Jiranek, V. Microbial modulation of aromatic esters in wine: Current knowledge and future prospects. Food Chem. 2010, 121, 1–16. [Google Scholar] [CrossRef]
- Perestrelo, R.; Fernandes, A.; Albuquerque, F.F.; Marques, J.C.; Câmara, J.S. Analytical characterization of the aroma of Tinta Negra Mole red wine: Identification of the main odorants compounds. Anal. Chim. Acta 2006, 563, 154–164. [Google Scholar] [CrossRef] [Green Version]
- Moreno, D.; Valdés, E.; Uriarte, D.; Gamero, E.; Talaverano, I.; Vilanova, M. Early leaf removal applied in warm climatic conditions: Impact on Tempranillo wine volatiles. Food Res. Int. 2017, 98, 50–58. [Google Scholar] [CrossRef]
- Šuklje, K.; Antalick, G.; Coetzee, Z.; Schmidtke, L.M.; Baša Česnik, H.; Brandt, J.; du Toit, W.J.; Lisjak, K.; Deloire, A. Effect of leaf removal and ultraviolet radiation on the composition and sensory perception of Vitis vinifera L. cv. Sauvignon Blanc wine. Aust. J. Grape Wine Res. 2014, 20, 223–233. [Google Scholar] [CrossRef]
- Vilanova, M.; Diago, M.P.; Genisheva, Z.; Oliveira, J.M.; Tardaguila, J. Early leaf removal impact on volatile composition of Tempranillo wines. J. Sci. Food Agric. 2012, 92, 935–942. [Google Scholar] [CrossRef]
- Song, J.; Smart, R.E.; Dambergs, R.G.; Sparrow, A.M.; Wells, R.B.; Wang, H.; Qian, M.C. Pinot Noir wine composition from different vine vigour zones classified by remote imaging technology. Food Chem. 2014, 153, 52–59. [Google Scholar] [CrossRef]
- Bubola, M.; Rusjan, D.; Lukić, I. Crop level vs. leaf removal: Effects on Istrian Malvasia wine aroma and phenolic acids composition. Food Chem. 2020, 312, 126046. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Palomo, E.; Gómez García-Carpintero, E.; Alonso-Villegas, R.; González-Viñas, M.A. Characterization of aroma compounds of Verdejo white wines from the La Mancha region by odour activity values. Flavour Fragr. J. 2010, 25, 456–462. [Google Scholar] [CrossRef]
- Sun, Q.; Sacks, G.; Lerch, S.; Vanden Heuvel, J.E. Impact of shoot thinning and harvest date on yield components, fruit composition, and wine quality of Marechal Foch. Am. J. Enol. Vitic. 2011, 62, 32–41. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.Q.; Cheng, G.; Duan, L.L.; Jiang, R.; Pan, Q.H.; Duan, C.Q.; Wang, J. Effect of training systems on fatty acids and their derived volatiles in Cabernet Sauvignon grapes and wines of the north foot of Mt. Tianshan. Food Chem. 2015, 181, 198–206. [Google Scholar] [CrossRef]
- Escudero, A.; Campo, E.; Fariña, L.; Cacho, J.; Ferreira, V. Analytical characterization of the aroma of five premium red wines. Insights into the role of odor families and the concept of fruitiness of wines. J. Agric. Food Chem. 2007, 55, 4501–4510. [Google Scholar] [CrossRef] [PubMed]
- Boidron, J.-N.; Chatonnet, P.; Pons, M. Influence du bois sur certaines substances odorantes des vins. OENO One 1988, 22, 275. [Google Scholar] [CrossRef]
- Castro Mejías, R.; Natera Marín, R.; García Moreno, M.D.V.; García Barroso, C. Optimisation of headspace solid-phase microextraction for the analysis of volatile phenols in wine. J. Chromatogr. A 2003, 995, 11–20. [Google Scholar] [CrossRef]
- Cheynier, V.; Schneider, R.; Salmon, J.-M.; Fulcrand, H. 3.26-Chemistry of Wine. In Comprehensive Natural Products II; Liu, H.-W., Mander, L., Eds.; Elsevier: Oxford, UK, 2010; pp. 1119–1172. ISBN 978-0-08-045382-8. [Google Scholar]
- Cataldo, E.; Salvi, L.; Paoli, F.; Fucile, M.; Mattii, G.B. Effect of agronomic techniques on aroma composition of white grapevines: A review. Agronomy 2021, 11, 2027. [Google Scholar] [CrossRef]
- Alem, H.; Rigou, P.; Schneider, R.; Ojeda, H.; Torregrosa, L. Impact of agronomic practices on grape aroma composition: A review. J. Sci. Food Agric. 2019, 99, 975–985. [Google Scholar] [CrossRef]
- Alem, H.; Ojeda, H.; Rigou, P.; Schneider, R.; Torregrosa, L. The reduction of plant sink/source does not systematically improve the metabolic composition of Vitis vinifera white fruit. Food Chem. 2021, 345, 128825. [Google Scholar] [CrossRef]
- Škrab, D.; Sivilotti, P.; Comuzzo, P.; Voce, S.; Degano, F.; Carlin, S.; Arapitsas, P.; Masuero, D.; Vrhovšek, U. Cluster thinning and vineyard site modulate the metabolomic profile of ribolla gialla base and sparkling wines. Metabolites 2021, 11, 331. [Google Scholar] [CrossRef]
- Reynolds, A.G.; Wardle, D.A.; Naylor, A.P. Impact of training system, vine spacing, and basal leaf removal on riesling. Vine performance, berry composition, canopy microclimate, and vineyard labor requirements. Am. J. Enol. Vitic. 1996, 47, 63–76. [Google Scholar]
- Zoecklein, B.W.; Wolf, T.K.; Pélanne, L.; Miller, M.K.; Birkenmaier, S.S. Effect of vertical shoot-positioned, Smart-Dyson, and Geneva double-curtain training systems on viognier grape and wine composition. Am. J. Enol. Vitic. 2008, 59, 11–21. [Google Scholar]
- Iorio, D.; Gambacorta, G.; Tarricone, L.; Vilanova, M.; Paradiso, V.M. Effect of Early Basal Leaf Removal on Phenolic and Volatile Composition and Sensory Properties of Aglianico Red Wines. Plants 2022, 11, 591. [Google Scholar] [CrossRef]
- Giovanelli, G.; Brenna, O.V. Evolution of some phenolic components, carotenoids and chlorophylls during ripening of three Italian grape varieties. Eur. Food Res. Technol. 2007, 225, 145–150. [Google Scholar] [CrossRef]
- Etchebarne, F.; Terblanche, E.; Iacono, M.B.; Leclercq, L.; Angenieux, M.; Saurin, N.; Ojeda, H. Minimal pruning increases the concentration of aromatic precursors in viognier grapes. In Proceedings of the 19èmes Journées Internationalesde Viticulture GiESCO, Gruissan, France, 31 May–5 June 2015; p. 810. [Google Scholar]
- Romero, P.; Botía, P.; del Amor, F.M.; Gil-Muñoz, R.; Flores, P.; Navarro, J.M. Interactive effects of the rootstock and the deficit irrigation technique on wine composition, nutraceutical potential, aromatic profile, and sensory attributes under semiarid and water limiting conditions. Agric. Water Manag. 2019, 225, 105733. [Google Scholar] [CrossRef]
- Vilanova, M.; Fandiño, M.; Frutos-Puerto, S.; Cancela, J.J. Assessment fertigation effects on chemical composition of Vitis vinifera L. cv. Albariño. Food Chem. 2019, 278, 636–643. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| T° mean | T° max | T° min | ET0 (mm) | Rainfall (mm) | |
|---|---|---|---|---|---|
| April | 10.3 | 23.2 | −1 | 87.6 | 44 |
| May | 14.3 | 30.1 | 2 | 130.1 | 30.7 |
| June | 18.1 | 33.4 | 7.5 | 142.2 | 87.9 |
| July | 20.1 | 37.3 | 9.3 | 162.8 | 0.7 |
| August | 20.4 | 40.4 | 9.2 | 149.5 | 4.6 |
| September | 18.6 | 31.1 | 7.9 | 93.7 | 30.3 |
| 2021 | 12.8 | 40.4 | −2.2 | 1033.7 | 467.7 |
| Average (2005–2021) | 12.5 | 40.8 | −9.2 | 1097.1 | 562.6 |
| LA | PSL | LT | PW | SP | NMS | |
|---|---|---|---|---|---|---|
| HT-MW | 3.99 ± 1.02 | 139.3 ± 34.3 | 9.78 ± 3.10 | 0.85 ± 0.01 | 5.75 ± 0.35 | 12.2 ± 0.31 |
| HT-WS | 3.95 ± 0.51 | 161.5 ± 28.8 | 9.39 ± 3.09 | 0.75 ± 0.05 | 5.50 ± 0.00 | 11.3 ± 0.35 |
| VCT-MW | 5.99 ± 0.94 | 107.6 ± 23.9 | 7.37 ± 2.21 | 0.80 ± 0.07 | 10.8 ± 0.47 | 21.8 ± 0.47 |
| VCT-WS | 5.93 ± 0.88 | 121.7 ± 31.2 | 7.06 ± 2.82 | 0.89 ± 0.07 | 10.2 ± 0.12 | 20.8 ± 0.82 |
| TS | *** | *** | * | ns | *** | *** |
| I | ns | * | ns | ns | ns | ns |
| TS × I | ns | ns | ns | ns | ns | ns |
| Midday Leaf Water Potential (MPa) | |||
|---|---|---|---|
| Flowering | Veraison | Ripening | |
| HT-MW | −1.03 ± 0.04 | −1.41 ± 0.08 | −1.39 ± 0.03 |
| HT-WS | −1.06 ± 0.05 | −1.60 ± 0.08 | −1.56 ± 0.11 |
| VCT-MW | −0.98 ± 0.07 | −1.34 ± 0.03 | −1.51 ± 0.03 |
| VCT-WS | −1.06 ± 0.05 | −1.60 ± 0.10 | −1.64 ± 0.07 |
| TS | ns | ns | ** |
| I | ns | *** | *** |
| TS × I | ns | ns | ns |
| Yield | Clusters Per Shoot | Cluster Weight | Berry Weight | Berries Per Cluster | Leaf Area: Yield | Yield: Pruning Weight | |
|---|---|---|---|---|---|---|---|
| kg Vine−1 | Clusters Shoot−1 | g Cluster−1 | g | Berries Cluster−1 | m2 kg−1 | kg kg−1 | |
| HT-MW | 2.19 ± 0.39 | 1.52 ± 0.25 | 117.7 ± 4.1 | 1.48 ± 0.03 | 79.88 ± 4.23 | 1.82 ± 0.35 | 2.57 ± 0.46 |
| HT-WS | 1.58 ± 0.25 | 1.36 ± 0.16 | 102.7 ± 6.5 | 1.22 ± 0.09 | 84.14 ± 1.21 | 2.72 ± 0.54 | 2.15 ± 0.50 |
| VCT-MW | 3.95 ± 0.27 | 1.70 ± 0.10 | 108.9 ± 3.9 | 1.20 ± 0.04 | 90.52 ± 3.74 | 1.55 ± 0.14 | 4.95 ± 0.35 |
| VCT-WS | 2.97 ± 0.26 | 1.40 ± 0.09 | 102.6 ± 15 | 1.06 ± 0.11 | 96.30 ± 4.95 | 2.02 ± 0.26 | 3.38 ± 0.48 |
| TS | *** | ns | ns | ** | ** | * | *** |
| I | ** | ns | ns | ** | ns | * | * |
| TS × I | ns | ns | ns | ns | ns | ns | ns |
| TSS | PA | pH | TA | Tart. Acid | MA | K | YAN | TPI | |
|---|---|---|---|---|---|---|---|---|---|
| HT-MW | 23.23 ± 0.1 | 13.61 ± 0.1 | 3.24 ± 0.05 | 4.22 ± 0.04 | 6.89 ± 0.21 | 0.40 ± 0.03 | 828.31 ± 67.17 | 246.33 ± 8.50 | 47.00 ± 4.54 |
| HT-WS | 23.33 ± 0.5 | 13.68 ± 0.4 | 3.26 ± 0.01 | 4.32 ± 0.16 | 7.27 ± 0.41 | 0.28 ± 0.05 | 810.31 ± 36.34 | 292.67 ± 14.38 | 63.03 ± 2.11 |
| VCT-MW | 22.73 ± 0.7 | 13.27 ± 0.5 | 3.18 ± 0.03 | 4.33 ± 0.07 | 6.59 ± 0.21 | 0.24 ± 0.00 | 700.68 ± 15.88 | 276.33 ± 22.22 | 51.27 ± 5.65 |
| VCT-WS | 23.23 ± 0.3 | 13.62 ± 0.2 | 3.14 ± 0.04 | 4.81 ± 0.28 | 7.17 ± 0.16 | 0.25 ± 0.05 | 756.35 ± 15.84 | 258.00 ± 15.94 | 65.83 ± 1.59 |
| TS | ns | ns | ** | ** | ns | ** | * | ns | ns |
| I | ns | ns | ns | * | * | ns | ns | ns | ** |
| TS × I | ns | ns | ns | ns | ns | * | ns | * | ns |
| Compounds | HT-MW | HT-WS | VCT-MW | VCT-WS | TS | I | TS × I |
|---|---|---|---|---|---|---|---|
| Higher alcohols | |||||||
| 1-Propanol | 103.79 ± 14.23 | 180.82 ± 17.18 | 399.55 ± 37.17 | 320.42 ± 66.52 | ** | ns | ns |
| 1-Butanol | 24.97 ± 2.80 | 38.80 ± 1.56 | 41.60 ± 4.26 | 38.85 ± 7.05 | ns | ns | ns |
| Isobutanol | 777.13 ± 98.78 | 852.32 ± 40.02 | 1309.44 ± 121.38 | 1348.60 ± 243.62 | * | ns | ns |
| Isopentanol | 16,164.55 ± 1734.46 | 18,261.18 ± 584.11 | 28,444.31 ± 1684.55 | 26,369.24 ± 2238.56 | ** | ns | ns |
| 3-Methyl-1-pentanol | 13.62 ± 1.19 | 19.82 ± 0.53 | 16.05 ± 0.82 | 13.11 ± 0.92 | ns | ns | ** |
| Benzyl alcohol | 23.10 ± 2.37 | 31.35 ± 1.43 | 39.09 ± 2.97 | 21.95 ± 0.75 | ns | ns | ** |
| 2-Phenylethanol | 35,867.42 ± 2769.13 | 37,085.85 ± 1553.82 | 57,775.58 ± 4485.54 | 32,759.28 ± 1856.92 | * | * | * |
| Methionol | 96.50 ± 14.54 | 69.25 ± 4.26 | 76.64 ± 6.36 | 41.45 ± 3.59 | * | * | ns |
| C6 compounds | |||||||
| 1-hexanol | 194.88 ± 15.27 | 248.59 ± 2.65 | 376.01 ± 17.37 | 300.72 ± 30.43 | *** | ns | * |
| trans-3-Hexen-1-ol | 36.16 ± 3.02 | 42.57 ± 2.31 | 62.97 ± 3.15 | 60.53 ± 2.20 | *** | ns | ns |
| cis-3-Hexen-1-ol | 5.89 ± 0.26 | 6.91 ± 0.06 | 12.74 ± 1.66 | 8.80 ± 1.67 | * | ns | ns |
| trans-2-Hexen-1-ol | 0.16 ± 0.07 | 0.38 ± 0.42 | 0.59 ± 0.46 | 0.41 ± 0.39 | ns | ns | ns |
| Terpenes + C13 norisoprenoids | |||||||
| β-Citronellol | 0.50 ± 0.45 | 1.61 ± 2.10 | 1.67 ± 2.10 | 1.74 ± 1.63 | ns | ns | ns |
| Nerol | 49.11 ± 10.04 | 46.76 ± 7.13 | 88.89 ± 5.56 | 81.51 ± 9.71 | ** | ns | ns |
| Geraniol | 5.65 ± 0.29 | 6.56 ± 0.02 | 5.68 ± 0.10 | 4.84 ± 0.30 | ** | ns | ** |
| β-pinene | 0.19 ± 0.09 | 0.29 ± 0.20 | 0.38 ± 0.30 | 0.39 ± 0.43 | ns | ns | ns |
| Linalool | 2.78 ± 0.44 | 4.58 ± 0.07 | 5.87 ± 1.49 | 4.42 ± 0.66 | ns | ns | ns |
| Terpinolene | nd | nd | 0.35 ± 0.49 | nd | ns | ns | ns |
| α-Ionone | 0.21 ± 0.30 | 0.49 ± 0.70 | 1.81 ± 0.89 | 0.80 ± 0.85 | ns | ns | ns |
| β-Ionone | 0.08 ± 0.06 | 0.11 ± 0.16 | 0.24 ± 0.23 | 0.24 ± 0.24 | ns | ns | ns |
| Damascenone | 6.29 ± 1.52 | 6.51 ± 0.81 | 8.70 ± 1.40 | 6.66 ± 1.66 | * | ns | ns |
| Ethyl esters | |||||||
| Ethyl butanoate | 122.96 ± 4.23 | 119.07 ± 1.88 | 250.52 ± 16.07 | 222.53 ± 27.21 | *** | ns | ns |
| Ethyl 2-methylbutyrate | 5.47 ± 0.18 | 6.56 ± 0.18 | 7.82 ± 0.92 | 6.04 ± 0.04 | ns | ns | * |
| Ethyl hexanoate | 539.12 ± 9.38 | 589.15 ± 2.99 | 856.60 ± 80.77 | 840.66 ± 131.10 | ** | ns | ns |
| Ethyl lactate | 683.50 ± 118.27 | 1119.24 ± 112.04 | 1456.27 ± 118.74 | 1221.16 ± 215.75 | * | ns | * |
| Ethyl isovalerate | 8.61 ± 1.61 | 10.13 ± 0.69 | 15.21 ± 1.73 | 9.44 ± 1.21 | * | ns | * |
| Ethyl octanoate | 528.96 ± 51.86 | 553.05 ± 1.95 | 768.21 ± 63.73 | 688.15 ± 94.30 | * | ns | ns |
| Ethyl decanoate | 25.87 ± 2.14 | 25.46 ± 0.44 | 37.72 ± 3.78 | 40.46 ± 2.84 | ** | ns | ns |
| Diethyl succinate | 541.87 ± 1.48 | 1190.07 ± 44.87 | 370.87 ± 16.47 | 228.50 ± 37.08 | *** | *** | *** |
| Acetate esters | |||||||
| Isoamyl acetate | 2270.49 ± 43.67 | 1116.69 ± 25.99 | 5436.94 ± 281.29 | 4284.80 ± 518.20 | *** | ** | ns |
| Pheniethyl acetate | 1148.52 ± 25.03 | 733.56 ± 8.71 | 1947.42 ± 49.11 | 1116.84 ± 191.42 | *** | *** | * |
| Hexyl acetate | 33.88 ± 5.09 | 12.69 ± 0.18 | 97.84 ± 4.87 | 77.48 ± 12.19 | *** | * | ns |
| Fatty acids | |||||||
| Butanoic acid | 61.01 ± 9.30 | 82.92 ± 8.15 | 122.04 ± 12.27 | 128.05 ± 31.42 | * | ns | ns |
| Propanoic acid | 7.37 ± 1.49 | 10.25 ± 1.47 | 19.28 ± 2.56 | 17.12 ± 4.89 | * | ns | ns |
| Isobutyric acid | 60.69 ± 11.50 | 76.82 ± 4.68 | 105.62 ± 4.73 | 108.98 ± 25.79 | * | ns | ns |
| Hexanoic acid | 1219.41 ± 115.14 | 1371.61 ± 30.23 | 2166.27 ± 162.22 | 2164.70 ± 79.68 | *** | ns | ns |
| Octanoic acid | 1081.82 ± 7.56 | 1156.53 ± 23.26 | 1844.08 ± 124.45 | 1628.31 ± 306.78 | ** | ns | ns |
| cis-Geranic acid | 32.30 ± 5.06 | 40.07 ± 3.33 | 42.39 ± 3.44 | 28.08 ± 14.45 | ns | ns | ns |
| trans-Geranic acid | 0.17 ± 0.23 | 0.57 ± 0.22 | 0.29 ± 0.14 | 0.26 ± 0.17 | ns | ns | ns |
| Lactones | |||||||
| γ-Butirolactone | 77.91 ± 14.43 | 87.21 ± 6.46 | 154.55 ± 12.51 | 149.56 ± 18.97 | ** | ns | ns |
| Volatile phenols | |||||||
| 4-Vinylguaiacol | 50.58 ± 7.57 | 52.58 ± 0.10 | 82.12 ± 1.30 | 73.49 ± 9.32 | ** | ns | ns |
| 4-Vinylphenol | 19.59 ± 3.66 | 17.63 ± 3.98 | 34.74 ± 2.70 | 50.71 ± 9.40 | ** | ns | ns |
| Carbonyl compounds | |||||||
| Acetoin | 80.09 ± 20.04 | 242.31 ± 35.88 | 69.04 ± 6.56 | 59.45 ± 13.21 | ** | ** | ** |
| Aldehydes | |||||||
| Vanillin | 0.72 ± 0.37 | 0.84 ±.0.01 | 2.17 ± 0.18 | 1.58 ± 0.50 | ** | ns | ns |
| Compounds | Odor Descriptor | Odor Threshold (µg L−1) | Ref. | OAV | TS | I | TS × I | |||
|---|---|---|---|---|---|---|---|---|---|---|
| HT-MW | HT-WS | VCT-MW | VCT-WS | |||||||
| Higher alcohols | ||||||||||
| 1-Propanol | Alcohol, ripe fruit | 306,000 | [37] | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | * | ns | ns |
| 1-Butanol | Medicinal, phenolic | 150,000 | [37] | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | ns | ns | ns |
| Isobutanol | Fusel | 40,000 | [38] | 0.02 ± 0.00 | 0.02 ± 0.00 | 0.03 ± 0.00 | 0.03 ± 0.01 | * | ns | ns |
| Isopentanol | Alcohol, banana | 7000 | [39] | 2.31 ± 0.25 | 2.61 ± 0.08 | 4.06 ± 0.24 | 3.77 ± 0.32 | ** | ns | ns |
| 3-Methyl-1-pentanol | Herbaceous, cocoa | 50,000 | [40] | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | ns | ns | ** |
| Benzyl alcohol | Caramel, fruity | 200,000 | [41] | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | ns | ns | ** |
| 2-Phenylethanol | Floral, roses, lilac | 10,000 | [42] | 3.59 ± 0.28 | 3.71 ± 0.16 | 5.78 ± 0.45 | 3.28 ± 0.19 | * | * | * |
| Methionol | Baked vegetables | 1000 | [43] | 0.10 ± 0.01 | 0.07 ± 0.00 | 0.08 ± 0.01 | 0.04 ± 0.00 | * | * | ns |
| C6 compounds | ||||||||||
| 1-hexanol | Green, cut grass | 8000 | [40] | 0.02 ± 0.00 | 0.03 ± 0.00 | 0.05 ± 0.00 | 0.04 ± 0.00 | *** | ns | * |
| trans-3-Hexen-1-ol | Green, floral | 400 | [44] | 0.09 ± 0.01 | 0.11 ± 0.00 | 0.16 ± 0.01 | 0.15 ± 0.00 | *** | ns | ns |
| cis-3-Hexen-1-ol | Green, cut grass | 400 | [40] | 0.01 ± 0.00 | 0.02 ± 0.00 | 0.03 ± 0.00 | 0.02 ± 0.00 | * | ns | ns |
| trans-2-Hexen-1-ol | Herbaceous, green | 15,000 | [45] | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | ns | ns | ns |
| Terpenes + C13 norisoprenoids | ||||||||||
| β-Citronellol | Rose | 100 | [46] | 0.00 ± 0.00 | 0.02 ± 0.10 | 0.02 ± 0.01 | 0.02 ± 0.01 | ns | ns | ns |
| Nerol | Fresh, sweet, rose-like | 500 | [47] | 0.10 ± 0.01 | 0.09 ± 0.01 | 0.18 ± 0.01 | 0.16 ± 0.01 | ** | ns | ns |
| Geraniol | Roses, geranium | 20 | [36] | 0.28 ± 0.01 | 0.33 ± 0.00 | 0.28 ± 0.03 | 0.24 ± 0.01 | ** | ns | ** |
| β-pinene | Woody | 1500 | [47] | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | ns | ns | ns |
| Linalool | Green, floral, sweet | 15 | [47] | 0.19 ± 0.02 | 0.31 ± 0.00 | 0.39 ± 0.07 | 0.29 ± 0.03 | ns | ns | ns |
| Terpinolene | Woody, sweet, citrus | 41 | [47] | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.01 ± 0.01 | 0.00 ± 0.00 | ns | ns | ns |
| α-Ionone | Sweet fruit | 2.6 | [40] | 0.08 ± 0.08 | 0.19 ± 0.19 | 0.69 ± 0.24 | 0.31 ± 0.23 | ns | ns | ns |
| β-Ionone | Violets, rose | 0.09 | [40] | 0.89 ± 0.44 | 1.25 ± 1.25 | 2.67 ± 1.79 | 2.94 ± 1.90 | ns | ns | ns |
| Damascenone | Sweet, exotic flowers, stewed | 0.14 | [48] | 38.80 ± 2.24 | 43.25 ± 1.40 | 67.66 ± 2.82 | 52.40 ± 8.40 | * | ns | ns |
| Ethyl esters | ||||||||||
| Ethyl butanoate | Papaya, apple, sweet | 20 | [49] | 6.15 ± 0.15 | 5.95 ± 0.07 | 12.53 ± 0.57 | 11.13 ± 0.96 | *** | ns | ns |
| Ethyl 2-methylbutyrate | Fruity, strawberry, apple, blackberry | 2 | [50] | 2.73 ± 0.06 | 3.28 ± 0.06 | 3.91 ± 0.33 | 3.02 ± 0.01 | ns | ns | * |
| Ethyl hexanoate | Apple, fruity, sweet | 14 | [49] | 38.51 ± 0.47 | 42.08 ± 0.15 | 61.19 ± 4.08 | 60.05 ± 6.62 | ** | ns | ns |
| Ethyl lactate | Strawberry, raspberry | 154,000 | [42] | 0.00 ± 0.00 | 0.01 ± 0.00 | 0.01 ± 0.00 | 0.01 ± 0.00 | *** | *** | *** |
| Ethyl isovalerate | Fruity, strawberry, apple | 0.7 | [50] | 12.30 ± 1.63 | 14.47 ± 0.69 | 21.73 ±1.75 | 13.48 ± 1.23 | * | ns | * |
| Ethyl octanoate | Apple. Fruity | 5 | [49] | 105.79 ± 7.33 | 110.61 ± 0.28 | 153.64 ± 9.01 | 137.63 ± 13.36 | * | ns | ns |
| Ethyl decanoate | Fruity, grape | 200 | [42] | 0.13 ± 0.01 | 0.13 ± 0.00 | 0.19 ± 0.01 | 0.20 ± 0.01 | ** | ns | ns |
| Diethyl succinate | Light fruity, wine | 6000 | [42] | 0.09 ± 0.00 | 0.20 ± 0.01 | 0.06 ± 0.00 | 0.04 ± 0.00 | *** | *** | *** |
| Acetate esters | ||||||||||
| Isoamyl acetate | Banana | 30 | [38] | 75.68 ± 1.03 | 37.22 ± 0.61 | 181.23 ± 6.63 | 142.83 ± 12.21 | *** | ** | ns |
| Pheniethyl acetate | Banana, floral | 250 | [38] | 4.59 ± 0.07 | 2.93 ± 0.02 | 7.79 ± 0.13 | 4.47 ± 0.54 | *** | *** | * |
| Hexyl acetate | Green, floral | 1500 | [40] | 0.02 ± 0.00 | 0.01 ± 0.00 | 0.07 ± 0.00 | 0.05 ± 0.01 | *** | * | ns |
| Fatty acids | ||||||||||
| Butanoic acid | Sweaty | 173 | [51] | 0.35 ± 0.04 | 0.48 ± 0.03 | 0.71 ± 0.05 | 0.74 ± 0.13 | * | ns | ns |
| Propanoic acid | Butter, rancid | 8100 | [41] | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | ns | ns | ns |
| Isobutyric acid | Butter, cheese, rancid | 2300 | [40] | 0.03 ± 0.00 | 0.03 ± 0.00 | 0.05 ± 0.00 | 0.05 ± 0.01 | * | ns | ns |
| Hexanoic acid | Cheese, fatty | 3000 | [42] | 0.41 ± 0.03 | 0.46 ± 0.01 | 0.72 ± 0.04 | 0.72 ± 0.02 | *** | ns | ns |
| Octanoic acid | Cheese, fatty, rancid | 1000 | [42] | 1.08 ± 0.01 | 1.16 ± 0.02 | 1.84 ± 0.09 | 1.63 ± 0.22 | ** | ns | ns |
| cis-Geranic acid | Green | 40 | [52] | 0.81 ± 0.09 | 1.00 ± 0.06 | 1.06 ± 0.06 | 0.70 ± 0.25 | ns | ns | ns |
| trans-Geranic acid | Green | 40 | [52] | 0.00 ± 0.00 | 0.01 ± 0.00 | 0.01 ± 0.00 | 0.01 ± 0.00 | ns | ns | ns |
| Lactones | ||||||||||
| γ-Butyrolactone | Toast, sweet, caramel | 35,000 | [53] | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | ns | ns | ns |
| Volatile phenols | ||||||||||
| 4-Vinylguaiacol | Clove, curry | 40 | [40] | 1.26 ± 0.13 | 1.31 ± 0.00 | 2.05 ± 0.02 | 1.84 ± 0.16 | ** | ns | ns |
| 4-Vinylphenol | Smoky, almond | 180 | [40] | 0.11 ± 0.01 | 0.10 ± 0.02 | 0.19 ± 0.01 | 0.28 ± 0.03 | ** | ns | ns |
| Carbonyl compounds | ||||||||||
| Acetoin | Lactic | 10,000 | [43] | 0.01 ± 0.00 | 0.02 ± 0.00 | 0.01 ± 0.00 | 0.01 ± 0.00 | * | * | * |
| Aldehydes | ||||||||||
| Vanillin | Vanillin | 200 | [54] | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.01 ± 0.00 | 0.01 ± 0.00 | ns | ns | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puelles, M.; Balda, P.; Labarga, D.; Mairata, A.; García-Escudero, E.; Guadalupe, Z.; Ayestarán, B.; Pou, A. Utilization of Vertical Cordon System to Improve Source-Sink Balance and Wine Aroma under Water Shortage Conditions of Maturana Blanca. Agronomy 2022, 12, 1373. https://doi.org/10.3390/agronomy12061373
Puelles M, Balda P, Labarga D, Mairata A, García-Escudero E, Guadalupe Z, Ayestarán B, Pou A. Utilization of Vertical Cordon System to Improve Source-Sink Balance and Wine Aroma under Water Shortage Conditions of Maturana Blanca. Agronomy. 2022; 12(6):1373. https://doi.org/10.3390/agronomy12061373
Chicago/Turabian StylePuelles, Miguel, Pedro Balda, David Labarga, Andreu Mairata, Enrique García-Escudero, Zenaida Guadalupe, Belén Ayestarán, and Alicia Pou. 2022. "Utilization of Vertical Cordon System to Improve Source-Sink Balance and Wine Aroma under Water Shortage Conditions of Maturana Blanca" Agronomy 12, no. 6: 1373. https://doi.org/10.3390/agronomy12061373
APA StylePuelles, M., Balda, P., Labarga, D., Mairata, A., García-Escudero, E., Guadalupe, Z., Ayestarán, B., & Pou, A. (2022). Utilization of Vertical Cordon System to Improve Source-Sink Balance and Wine Aroma under Water Shortage Conditions of Maturana Blanca. Agronomy, 12(6), 1373. https://doi.org/10.3390/agronomy12061373