The present work combined the evaluation of 21 ecotypes of duckweeds (grown under laboratory conditions) concerning their fiber, monosaccharide, amino acids, soluble sugars, total phenolic, and tannin contents. The biochemical data were used for an integrative analysis with retrieved data from a systematic literature review focusing on the food supply described above.
3.1. Biochemical Analysis of the 21 Ecotypes
The contents of carbon, nitrogen, proteins, soluble sugars, and cell wall (expressed as fiber percentage) of 21 duckweeds ecotypes are shown in
Table 1.
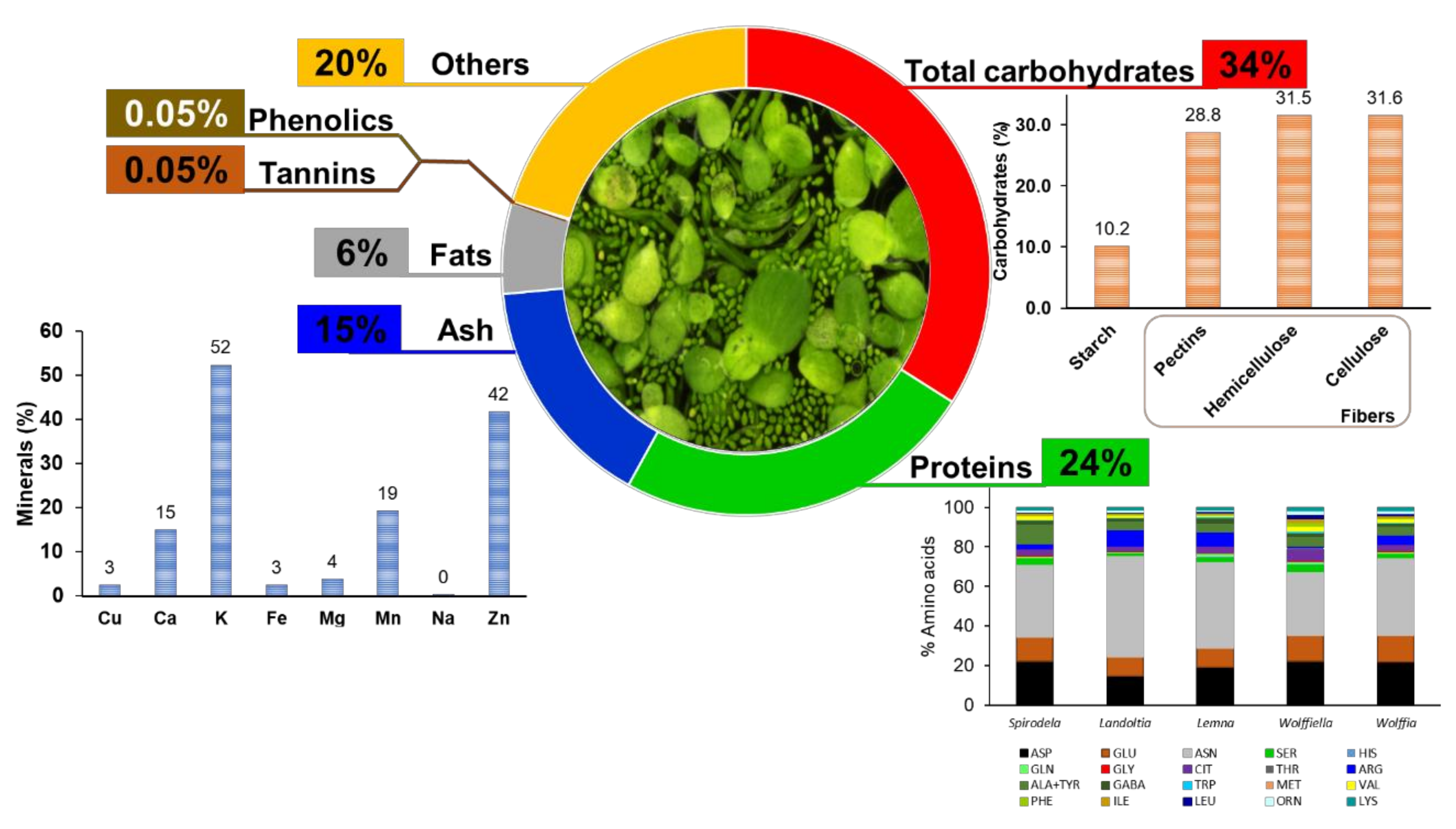
The carbon evaluation can be mostly related to the carbohydrates and the nitrogen to the protein. The carbon content was 37.8% among all genera, with no significant differences; the nitrogen and protein were 4.8 and 29.95%, respectively.
Landoltia punctata 7488 had 8.3% more proteins than the other duckweeds evaluated. The total amino acids also distinguish among the species, being higher in
Lemna gibba DWC 112 (18.2%) and
Wolffiella caudata 9139 (18.6%). The overall average of soluble sugars was 50% of the biomass from the carbon derivatives, and the fibers were 49% (
Table 1).
The nonstructural carbohydrates (glucose, fructose, sucrose, raffinose, and starch—
Table 2) and structural carbohydrates (fucose, arabinose, galactose, rhamnose, glucose, xylose, mannose, and apiose—
Table 3) were quantified. The glucose, fructose, sucrose, and raffinose were found in low proportions compared to starch (
Table 2). Among the non-starch sugars was fructose dominates, being usually higher than glucose, sucrose, and raffinose. Wolffioideae displayed approximately twice (106.9 µg·mg
−1) of the amount of starch compared to Lemnoideae (48 µg·mg
−1). The exceptions are two
Lemna species (
Lemna minor 8627—97.5 µg·mg
−1 and
Lemna gibba DWC112—138.5 µg·mg
−1) with similar starch contents to Wolffioideae (
Table 2).
The structural carbohydrates are the constituents of the cell wall. It is well-known that the apiose levels are a trade-off between the sub-families. Here, on average, was detected 80.1 µg·mg
−1 for Lemnoideae and 19.6 µg·mg
−1 for Wolffioideae (
Table 3). Among the genera,
Lemna species had a higher apiose level (92.5 µg·mg
−1) and
Wolffia the lower (14.1 µg·mg
−1). Another pentose, xylose, was also higher in the
Landoltia (24.7 µg·mg
−1) but also greater in
Wolffia (23.1 µg·mg
−1) and
Wolffiella (21.2 µg·mg
−1) (
Table 3).
Spirodela polyrhiza 7498 and 9624 had 2.5 times more xylose than the other
Spirodela lineages (
Table 3).
Wolffia globosa 884 had 1.48 times less xylose than the other
Wolffia species. Other pectin sugars, such as rhamnose and galactose, were different across the genera. Rhamnose was slightly higher in
Spirodela (4.3 µg·mg
−1) than in
Landoltia, Lemna, Wolffiella, and
Wolffia (3 µg·mg
−1), except for
Wolffiella neotropica 8848 (4.5 µg·mg
−1) and
Wolffia globosa 884 (4.6 µg·mg
−1). Galactose was slightly higher in
Spirodela (18.6 µg·mg
−1) and
Wolffiella (16.5 µg·mg
−1).
Spirodela polyrhiza 9346 had approximately half of the galactose compared to the other
Spirodela species evaluated.
Wolffia globosa 884 (18.6 µg·mg
−1),
Wolffiella gladiata 8261 (18.4 µg·mg
−1), and
Lemna gibba DWC112 (18.0 µg·mg
−1) had more galactose than the average of the evaluated species. The arabinose levels had higher variability between the species of the
Spirodela (ranging from 6.9 to 22.9 µg·mg
−1) and
Wolffiella (ranging from 23 to 44 µg·mg
−1) genera. The glucose levels ranged from 2.2–12.2 µg·mg
−1 across all genera, higher in
Lemna gibba DWC112 and Wolffioideae (
Table 3). All species had a considerable amount of mannose with 8.2 µg·mg
−1, a sugar constituent of the hemicellulose mannans. Furthermore, the duckweeds displayed low fucosylation (see the fucose levels in
Table 3—1.6 µg·mg
−1) similarly to sweet corn (0.2 µg·mg
−1), pumpkin (0.6 µg·mg
−1), oats (0.2 µg·mg
−1), green kiwifruit (1.4 µg·mg
−1), golden kiwifruit (1.1 µg·mg
−1), carrot (1 µg·mg
−1), and blackcurrant (1.2 µg·mg
−1) and differently from apple (3.6 µg·mg
−1) [
36].
The total amino acids were higher in the
Wolffia species (124 µg·mg
−1 dry weight).
Spirodela species have a reduction of 23%,
Lemna 38%,
Landoltia 48%, and
Wolffiella 54% in comparison to
Wolffia (
Table 1). Thus, the quantity and diversity of duckweeds amino acids are related to the species’ ecotype. The amino acids evaluated were divided into essential (
Table 4) and non-essential (
Table 5) to facilitate the interpretation from a food point of view. The human body does not synthesize the essential amino acids, and their supply must come from food intake. Besides alanine and lysine, all the essential amino acids were found in low levels (0–2000 mg·mg
−1 dry weight), while the nonessential amino acids displayed higher concentrations than the essential ones. The evaluated species have more than 70% as aspartic acid (Asp), glutamic acid (Glu), asparagine (Asn), 2% citrulline (Cit), 5% threonine (Thr), 6% alanine + tyrosine (Ala + Tyr), and 2% serine (Ser), and the remaining 15% are distributed into histidine (His), tryptophan (Trp), methionine (Met), valine (Val), isoleucine (Ile), leucine (Leu), lysine (Lys), serine (Ser), glutamine (Gln), Gamma-Aminobutyric Acid (GABA), and ornithine (Orn).
Landoltia had proportionally higher amounts of arginine and asparagine.
Wolffiella had more citrulline, tryptophan, methionine, valine, and phenylalanine, and
Wolffia had more glycine and arginine (
Table 4 and
Table 5).
Tannins are secondary metabolites in plants that confer stringency and bitterness flavors to food [
37]. In most cases, food containing tannins can decrease the quality of a particular food product [
37]. In the 21 samples evaluated, along with the data found in the literature of four occurrences,
Spirodela displayed lower levels of tannins in all genera (20 mg·mg
−1) (
Table 6), which represents less than 1% in the plant biomass. However, the phenolic content was distinct among the genera, with
Spirodela = Lemna = Wolffia > Landoltia = Wolffiella. The low content of tannins in
Spirodela might improve the organoleptic characteristics of this duckweed species due to the non-bitter flavor. This may be a critical factor in the acceptance of
Spirodela as a food product.
3.2. Systematic Review and Bibliometric Analysis
One way to recover data from the literature is through a systematic review. Our revision of 85 studies aimed to evaluate the nutrients of different duckweed genera (
Supplemental Figure S1). From the selected papers, 80 were found in the Scopus search, and five were added manually to analyze the cell walls. The genus
Lemna had the higher frequency data on the publications (206), followed by
Wolffia (165),
Spirodela (117),
Landoltia (47), and
Wolffiella (34) (
Table 7). In addition, the data of the amino acids (72), ash (48), carbohydrates (8), cell wall (13), energy (8), fat (81), fibers (58), minerals (24), protein (114), monosaccharides (37), starch (82), and tannin (24) were collected and evaluated.
A comprehensive systematic analysis of duckweed composition relevant for nutrition and food assessment with the unity of biochemical data to a bibliometric analysis enlightens the duckweed applications toward food. The systematic review includes using rigorous methodology according to current guidelines of conducting and reporting [
22]. The bibliometric analysis showed the distribution of the countries that studied duckweed regarding the relevance for food applications. China is ranked in the first position in numbers of publications (47 papers), followed by the United States (19 papers) and India (18 papers) (
Figure 1A). Asian countries were the significant contributors of scientific knowledge and are the largest duckweeds producers. The panel of author keywords demonstrates that most of the papers used here display words related to the fast growth of duckweeds, and the content of proteins, amino acids, and starch is the most recurrent and significant of human and animal feed (
Figure 1B). The keyword panel corroborates the frequency of the recovered data, being the genus
Lemna the most abundant variable in proteins (
Table 7).
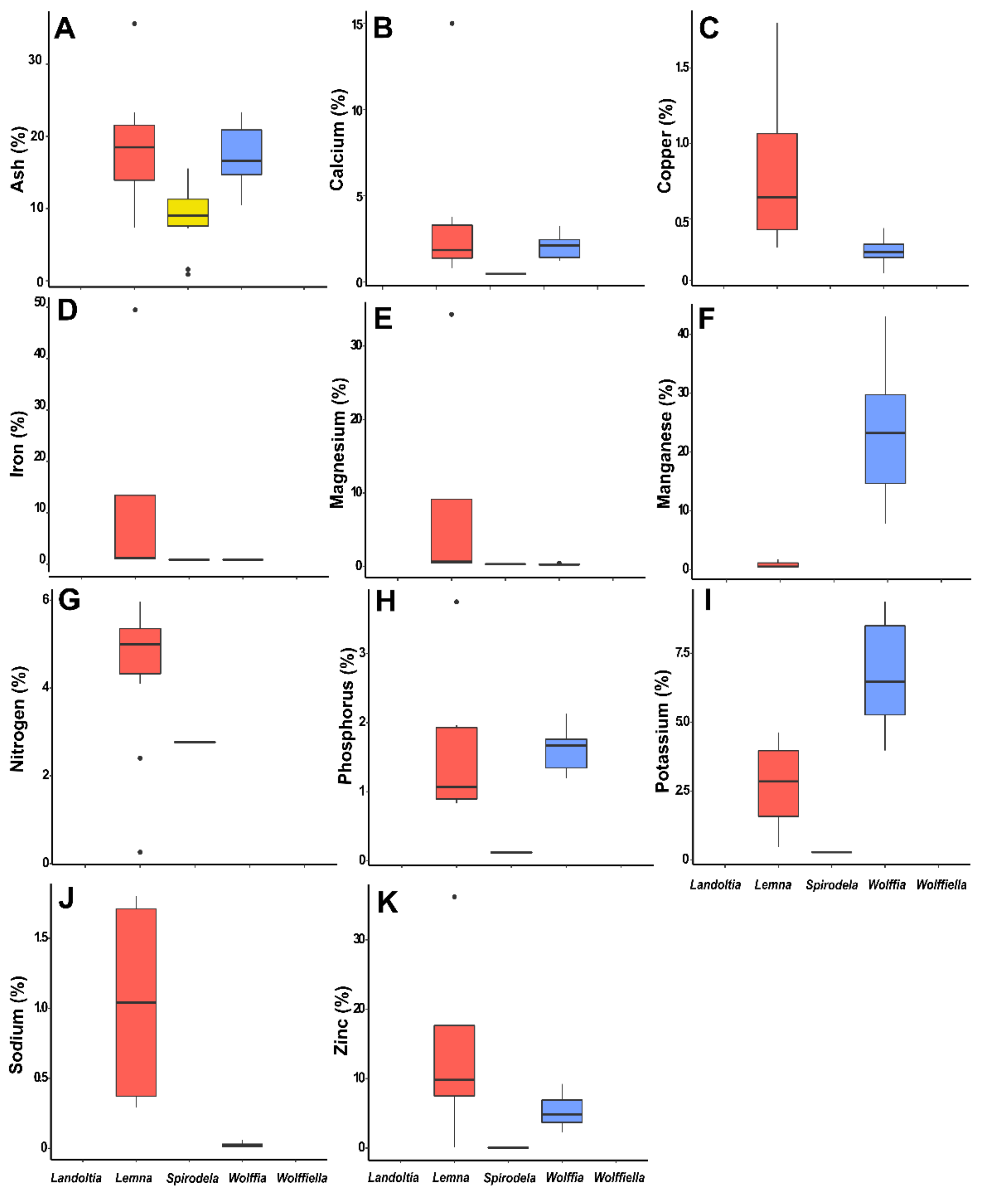
Fresh duckweed’s biomass is 92–94% water [
35], and the ash content is considered high compared to other plants. The ash content of plant tissues is mostly inorganics and is species-dependent [
38]. The ash data were collected from 20 publications with 48 occurrences, of which 12 were from
Spirodela, 15 from
Lemna, and 21 from
Wolffia (
Table 7). The ash content of the
Spirodela biomass ranges from 1 to 16%,
Lemna ranges from 7 to 36%, and
Wolffia from 10 to 23% (
Figure 2). The ash content is widely ranged in plants, being 0.1% for debarked woody chips, 3–12% for corn stover, 1–9% for
Miscanthus straw, 8–26% rice straw, 1–15% sugarcane bagasse, 3–23% wheat straw, and 0.2–6% for woody residues [
39]. Interestingly, the ash content of duckweeds is not influenced by the nutrient disponibility into the water body [
35].
Duckweeds can accumulate minerals (macro and micronutrients) several hundred folds compared to the concentration of minerals in the water they grow in [
40,
41]. As a result, these plants have the capacity for phytoremediation [
17]. The nutrients in duckweeds, obtained from 13 publications were calcium, copper, iron, potassium, magnesium, manganese, sodium, phosphorus, and zinc. Appenroth et al. (2018) evaluated the nutritional value of 11 species of the 16 ecotypes belonging to the genus
Wolffia [
17]. The mineral composition of
Wolffia was rich in potassium and iron and poor in sodium, manganese, zinc, and copper (
Figure 2).
Spirodela polyrhiza has a low content of minerals when compared to
Lemna and
Wolffia, especially for zinc (0.5 mg·g
−1), phosphorus (1.3 mg·g
−1), and potassium (2.8 mg·g
−1).
Lemna has a distinct nutrient pattern, with elevated levels of iron, magnesium, zinc, copper, and sodium (
Figure 2).
Plants are a rich source of minerals. Therefore, they can benefit the human diet and contain antinutrients that require processing before consumption [
1]. Healthy diets contain macro- and micronutrients needed by the organism, but some contain anti-nutrients like phytates and oxalates that inhibit iron and zinc absorption [
1]. The oxalate and phenolic compounds also are considered organoleptic inhibitory factors and interfere with digestion or metabolism [
42]. Druse and raphide shapes of calcium oxalates are reported in
Spirodela polyrhiza [
43] and
Lemna minor [
44] which play a role in calcium regulation of these plants. Nevertheless, the Wolffioideae plants have oxalate in the free form favoring them for food consumption [
17].
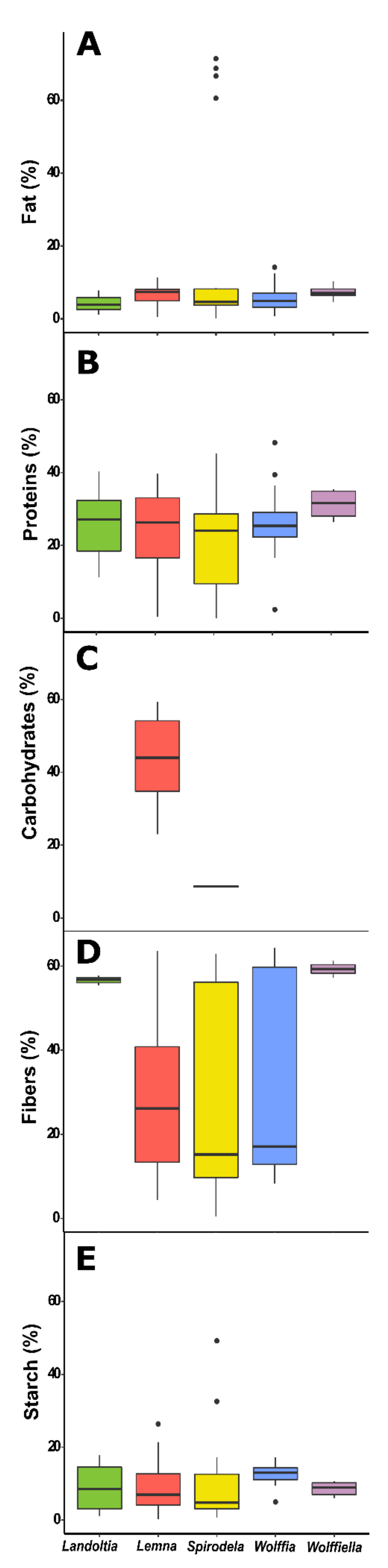
Proteins are macronutrients essential for the human diet once it is a source of amino acids. The protein content is variable among food sources, being 20–36% legumes, 8–25% nuts and seeds, 10–20% meat and fish, 15% eggs, 3.5% milk, and 1–3% vegetables [
45]. Nowadays, a plant-based trend in diets is growing worldwide. Plant-based protein provides 65% of the world’s supply [
45] and has the required amino acids for human intake [
46]. Duckweed protein is considered high quality [
19,
47], which can substitute some protein intakes. The protein data was the most frequently found (43 papers with 114 occurrences) (
Table 7). Variability among the duckweed genus was found, in which
Spirodela was higher, ranging from 0.5 to 45% and
Wolffia from 2 to 48%, followed by
Landoltia (11 to 40%),
Lemna (0.3–33%), and
Wolffiella from 26 to 35% (
Figure 3B). The protein content of duckweeds is related to the plants’ nutrition disponibility, temperature, and age [
19]. The content of plants grown in natural water bodies varies from 7 to 20%, while the plants grown in mineral media or effluents have a high protein content ranging from 30 to 40% [
19], which corroborates several studies.
A balanced diet requires daily protein to compensate for the nitrogen loss and provide protein for metabolism maintenance and growth [
46], equivalent to 10–18% of the total energy [
45]. Duckweed protein is highly digestible, which is important for diets due to the reuse of the amino acids ingested. The digestion was studied in fish [
18,
48,
49,
50,
51,
52,
53,
54,
55], pigs [
56], dogs [
57], and turtles [
58,
59], with a range from 65 to 90%. The digestibility of
Lemna minor was evaluated in comparison to green peas, revealing a lower capacity, but
Lemna minor also led to lower plasma glucose and insulin levels in humans [
60]. Thus, duckweeds can be a promising alternative for diabetics, reducing blood sugar and benefiting health.
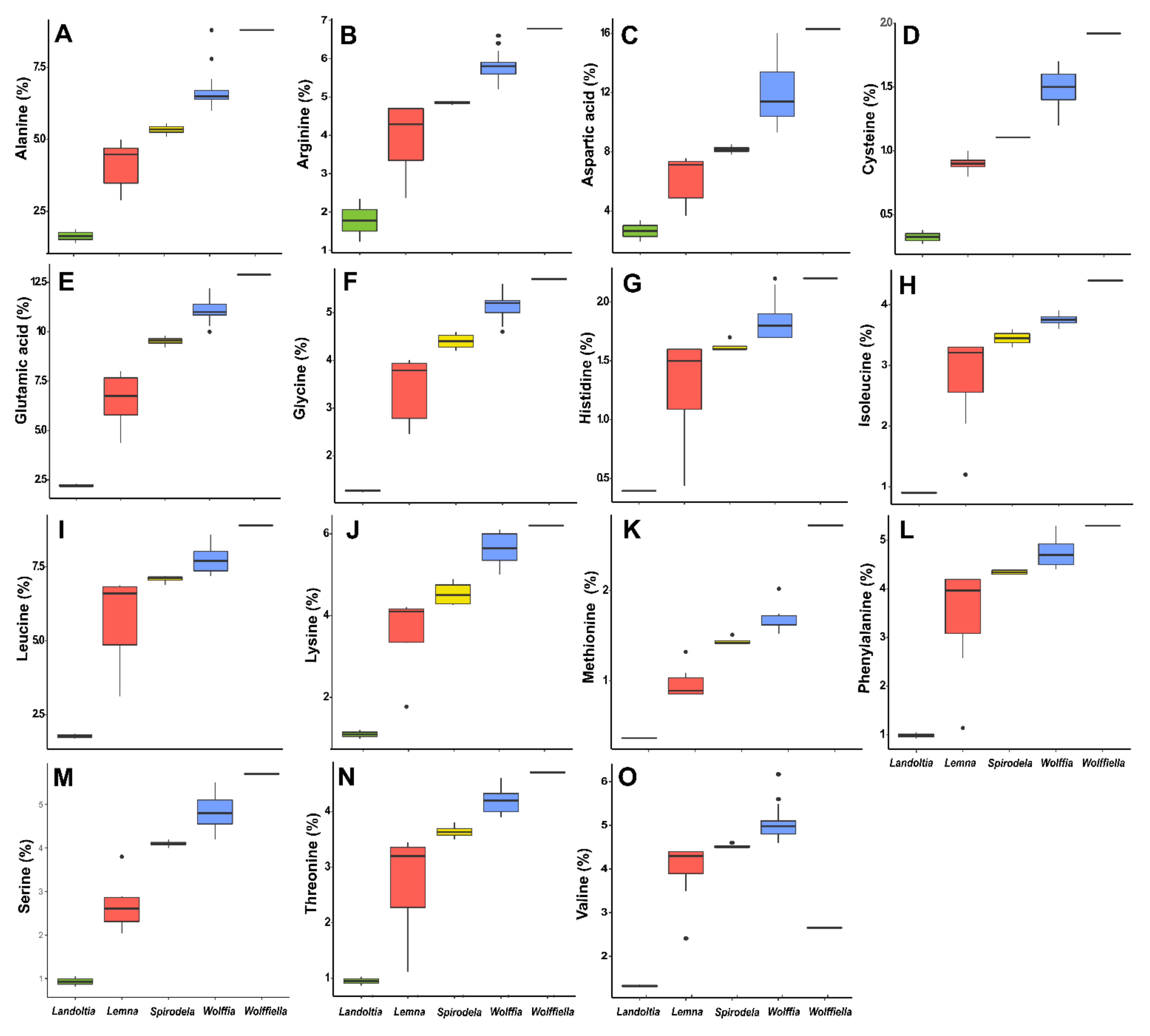
The amino acids can be divided into essential (not produced by the human organism) and nonessentials (produced by the human organism). Furthermore, the essential amino acids are not fully available depending on the plant source, like cereals with low lysine and legumes with low sulfur-containing amino acids [
46]. The amino acids lysine and arginine are essential for animal feed (~6% of the dietary protein), and the levels of these in duckweeds (up to 6%—
Figure 4) are comparable to those found in alfalfa [
40,
61]. Arginine is conditionally essential during pregnancy, critical illness, and infancy [
62], being necessary for human intake. The amino acid recovered data were analyzed as a percentage of the protein and biomass due to the nature of the data (
Figure 4 and
Supplemental Figure S2). The amino acids contents of the 21 ecotypes evaluated here had 72% of asparagine, glutamic acid, and aspartic acid (
Table 5), and asparagine and glutamic acid corresponded to 2% of the biomass (
Supplemental Figure S2). These amino acids play an essential role in protein synthesis and modification, the breakdown of ammonia, and the neurological system. Duckweed’s amino acid composition is similar to that found in other plants [
15,
19,
63]. They are rich in leucine, threonine, valine, isoleucine, and phenylalanine and poor in methionine and tyrosine [
40] (
Figure 4 and
Supplemental Figure S2). The cysteine + methionine was 22% higher than the WHO recommendation, and the levels of threonine, phenylalanine + tyrosine, and leucine also surpassed the intake recommendations [
15]. Porath et al. (1979) found methionine and lysine levels comparable to milk and cheese in seven duckweeds species (
Lemna gibba,
Lemna minor,
Spirodela polyrhiza, and
Wolffia arrhiza) [
47]. However, methionine was found in lower concentrations (100 mg·mg
−1 in the 21 ecotypes evaluated biochemically and 0.3–2.7% of protein) than the recommended (
Table 4 and
Figure 4). In general, for duckweeds, protein is a good source of lysine (2% of dry matter). Besides methionine, the amino acids isoleucine, leucine, phenylalanine, threonine, valine, and tryptophan correspond to the FAO reference pattern, fulfilling the requirement for human nutrition [
17], thus being an effective supplement in diets [
19] (
Figure 4 and
Supplemental Figure S2).
Energy balance is essential to maintain health through a secure nutrient intake [
20]. The metabolism of fatty acids generates twice the energy of the same amount of carbohydrate [
20]. However, low amounts of fats are recommended in the diet. Plants have a distinct nutritional composition, and vegetables have low-fat contents. The fat content in
Spirodela varies from 0.2 to 8%,
Landoltia 1 to 8%,
Lemna 0.5 to 11%,
Wolffiela 6 to 10%, and
Wolffia 0.7 to 14% (
Figure 3A). This data were recovered in 21 papers with the lipid data from 1980 to the present, with 81 publications (
Table 7). A wide range of fat contents was observed in
Spirodela´s literature (0.2–71%) due to the evaluation of the cuticles of four
Spirodela polyrhiza ecotypes by Borisjuk et al. (2018) [
64]. The variance in the fat content of duckweed is related to the growing system, climatic conditions, light intensity, and nutrient availability [
40,
55]. A fatty acid survey of thirty duckweeds species was performed by Yan et al. (2013) [
65]. The authors found that the total content varied between 5 and 14%, and the composition of the fats showed low variability with 80% palmitic acid (C16:0), linoleic acid (C18:2), and α-linolenic acid (C18:3) [
65], which corroborates the one found in plants [
15,
17,
40]. The main fatty acids from duckweeds (linoleic acid—C18:2 and α-linolenic acid—C18:3) are essential due to their role in retinal and brain development and the prevention of cardiovascular diseases [
20]. Thus, the intake of food with these fatty acids is indispensable for human health during all life. The recommended intake level is 2.5% of the total energy of linoleic acid and 0.5% of the total energy of α-linolenic acid [
20].
Saturated fatty food has greater relevance for human nutrition. In duckweeds, the high content of palmitic acid should be a hazard, but the levels of oleic acid, short-chain fatty acids, and medium-chain fatty acids are remarkably low [
20]. The high omega-6 and omega-3 ratios are involved in the development of cardiovascular diseases, cancer, osteoporosis, and inflammatory and autoimmune diseases [
17]. Therefore, it is recommended to consume food with a ratio between the two lipids below 5, which is not natural for some plants and additives [
20]. Furthermore, the omega-6 and omega-3 ratios were 0.36 among the duckweed species that Yan et al. (2013) evaluated. Thus, adding duckweed to the human diet would be interesting to reduce the unfavorable omega-6 and omega-3 ratio [
65].
Duckweeds display low-fat contents. Therefore, the energy of the intake of these plants comes from carbohydrates. Carbohydrates are the most abundant components of food, being polyhydroxy aldehydes; ketones; alcohols; acids; and their derivatives that are classified as sugars (glucose, galactose, fructose, sucrose, lactose, trehalose, sorbitol, and mannitol); oligosaccharides (maltodextrins, raffinose, stachyose, and fructooligosaccharides); and polysaccharides (starch, cellulose, hemicellulose, pectins, and hydrocolloids) [
66]. The carbohydrate content of duckweeds is mainly fibers instead of starch, like in soy grains [
52]. The total carbohydrate content in duckweeds was determined in eight publications with eight occurrences between
Spirodela and
Lemna (
Table 7). The content in
Lemna varies from 23 to 59%, and
Spirodela contains 8% carbohydrates (
Figure 3C). The highest report from
Lemna minor (59%) was calculated based upon the difference of a total content with the mineral content, protein, and lipids [
67]. A similar report was made by Zhao et al. (2014) with 51.2%, although the carbohydrate content seems to include the cell wall content [
68].
The primary energy source in human diets is carbohydrates, which constitutes sugars, starch, and other polysaccharides [
69]. Starch’s easy digestibility and the almost total absorbent capacity in the human trait make it a fast-energy assessment in the diet. Maize, rice, sorghum, wheat, barley, millet, rye, oats, potato, cassava, yam, and taro have 17–75% of starch and are the most consumed cereals and root crops [
70]. Therefore, starchy plants are value-added for the human diet as a source of nutrients and energy and in several industrial applications like thickener, texturizer, gelling agent, paper and board production, biodegradable plastics, and packaging materials [
71]. Due to the remarkable growth capacity, duckweed polysaccharides have importance for biorefinery applications. Starch in duckweeds can accumulate up to 70% into fronds [
72] and have distinct morphological, crystallinity, physicochemical, and thermal properties [
73]. The starch from these plants has a low peak of viscosity that is suitable for high-temperature processes and as materials for delivery systems [
73]. The systematic review resulted in 22 papers from 1997 to 2021, with 82 occurrences among the five genera (
Table 7).
Spirodela had the higher variation of starch content of the evaluated works with a range of 0.7–49%, while the other genera varied as follows:
Landoltia (1–17%),
Lemna (0.2%–10%),
Wolffiella (6–11%), and
Wolffia (5–14%) of the biomass (
Figure 3E). The growth system and conditions are essential for duckweeds biomass and starch production. Thus, the amplitude of the data reported in the literature must reflect this bias due to daytime and the light intensity that prevailed during the plant´s growth. Other factors might also have influenced the variety of the results of starch accumulation in duckweeds, such as temperature, light intensity, photoperiod, co-cultivation, and disponibility of nutrients [
72,
74,
75,
76,
77]. Duckweeds starch properties and granules morphology and the capacity for high starch accumulation make these plants suitable for diverse applications in the food industry and as “starchy vegetables”, especially
Spirodela, Landoltia, and
Lemna species due to the small sizes of the granules [
73].
It is well-known that a diet shift of high fat and high sugars is needed to improve human health. The replacement of high-fat meats with beans, peas, lentils, and other fibers is a public health concern [
70]. The diet fiber comprehends editable carbohydrates polymers mainly of plant origin with ten or more monomeric units that are not digested by endogenous enzymes and not absorbed by the small intestine of humans [
78]. The diet fiber includes cellulose, hemicellulose, pectin, gums, mucilages, and lignin [
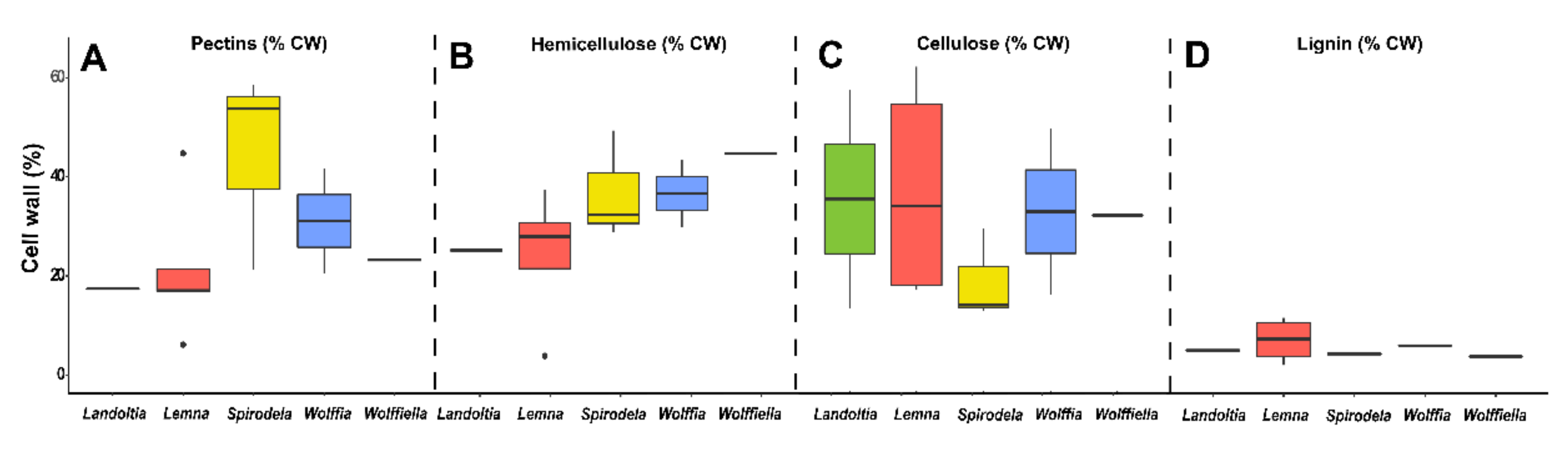
79]. Thus, it is possible to consider food fiber as the plant cell walls. The latter is a complex structure of polysaccharides, phenolic compounds, and structural proteins arranged in a glucose core (cellulose) with hemicellulose, lignin, and structural proteins immersed into a pectin matrix [
80]. Lignin is the phenolic compound responsible for polysaccharides binding, which is found in low proportion in duckweeds [
68,
81,
82,
83], assuming 5–10% of the cell wall (
Figure 5D). Cell wall proteins are quantitatively irrelevant.
The proportion of cellulose in duckweeds (13–17%) is thought to be three times smaller than inland plants [
84,
85], although some publications reported ~40% of cellulose into the cell walls [
68,
83] (
Figure 5C). This discrepancy might be related to the methodology employed during the cell wall fractionation process (
Figure 5C). Cellulose can be used as a delivery system, food stabilizer, functional food ingredient (due to the low or indigestibility), and biodegradable packaging [
86]. Hemicelluloses are composed of the monosaccharide’s glucose, arabinose, xylose, fucose, galactose, and mannose to form different classes of polysaccharides (e.g., xyloglucans, xylans, mannans, and β-glucans), assuming up to 29% of the duckweed cell wall (
Figure 5B). Moreover, cellulose can be employed as a green film for food packing [
87]. Mannans (polysaccharides made of mannose) are found in low proportions in most of the duckweeds (0.1–1% of dry mass), except for
Lemna perpusilla, which can reach 13% [
85] (
Figure 6G). These polymers display several applications in the food industry as edible films, gel formation, stiffeners, viscosity, modifiers, stabilizers, texture improvers, water-absorbent, prebiotics in dairy products, and seasonings [
88].
Pectins are acid polysaccharides whose backbones contain 70% of their structure as galacturonic acids interspaced with rhamnose. These are substituted with neutral polymers composed of fucose, galactose, apiose, xylose, and arabinose. The polymers formed are homogalacturonans, rhamnogalacturonans I and II, xylogalacturonans, and apiogalacturonans [
89]. Pectins in the primary cell wall of dicots and non-graminaceous monocots represent, on average, 35%, while only 2–10% of grasses and commelinids and 5% of woody tissues [
89]. Duckweeds are pectin-rich plants (20–54%) (
Figure 5 and
Figure 6) [
57,
68,
83,
84,
85,
90,
91] with an evolution trend with high proportions of apiogalacturonans (see the apiose levels—
Table 3 and
Figure 6A) in Lemnoideae and xylogalacturonans (see the xylose levels—
Table 3 and
Figure 6I) in Wolffioideae [
92].
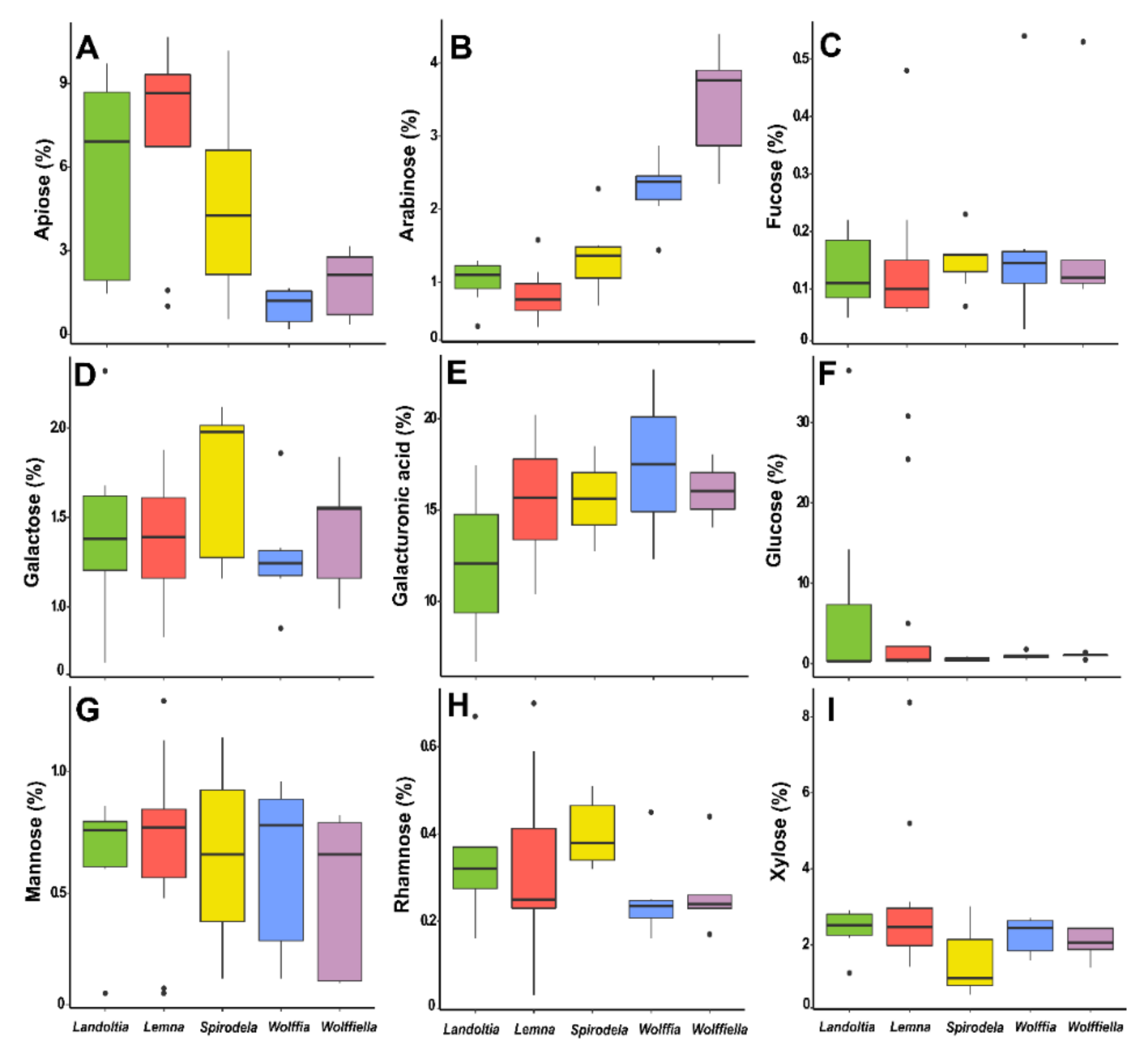
Apiose is higher in
Spirodela (0.5–10%),
Landoltia (1.4–10%), and
Lemna (1–11%) than in
Wolffiella (0.3–3%), and
Wolffia (0.1–2%) (
Figure 6A). The xylose content in the whole cell wall is computed together with the residues of xylans, xyloglucans, and xylogalacturonans. Thus, the pattern of higher xylose contents in the Wolffioideae was not identified (
Figure 6I).
Spirodela had 0.7–3%,
Landoltia 1–3%,
Lemna 1.5–14%,
Wolffiella 1–2%, and
Wolffia 1.6–3% of xylose into the cell walls (
Figure 6I). As previously mentioned, duckweeds have elevated pectins levels, so the concentration of galacturonic acid/uronic acids in this class of polysaccharides is 13–20% in
Spirodela, 7–17% in
Landoltia, 10–20% in
Lemna, 14–18% in
Wolffiella, and 12–23% in
Wolffia (
Figure 6E). The chemical properties of pectins make them suitable as a gelling agent, emulsifier, thickener, stabilizer, edible films or coating, and fat or sugar replacer in low-caloric food [
93,
94]. Such properties regarding the duckweed pectins remain to be studied.
A diet rich in fibers has several advantages, such as a reduction of appetite, lower blood sugar levels, reduced risk of heart disease, alleviating constipation, reduced risk of colorectal cancers, reducing the risk and symptoms of metabolic syndrome and diabetes, lowering cholesterol, balances intestinal pH, and stimulates intestinal fermentation [
79]. The fiber levels were identified in 20 works with 58 occurrences, 15 of
Spirodela, 3 of
Landoltia, 27 of
Lemna, 3 of
Wolffiella, and 10 of
Wolffia, ranging from 0.4 to 63%, 5 to 58%, 4 to 67%, 57 to 67%, and 8 to 64% respectively (
Table 7 and
Figure 3D). Thus, eating vegetables rich in the wall, like duckweeds, could help the fiber functions mentioned above and be advantageous for human health.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}