
Evolutionary Variation of Accumulative Day Length and Accumulative Active Temperature Required for Growth Periods in Global Soybeans


 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Field Experiments
2.2. Measurement of Growth-Period Traits
2.3. Statistical Analysis
2.4. SNP Genotyping and Clustering Analysis
3. Results
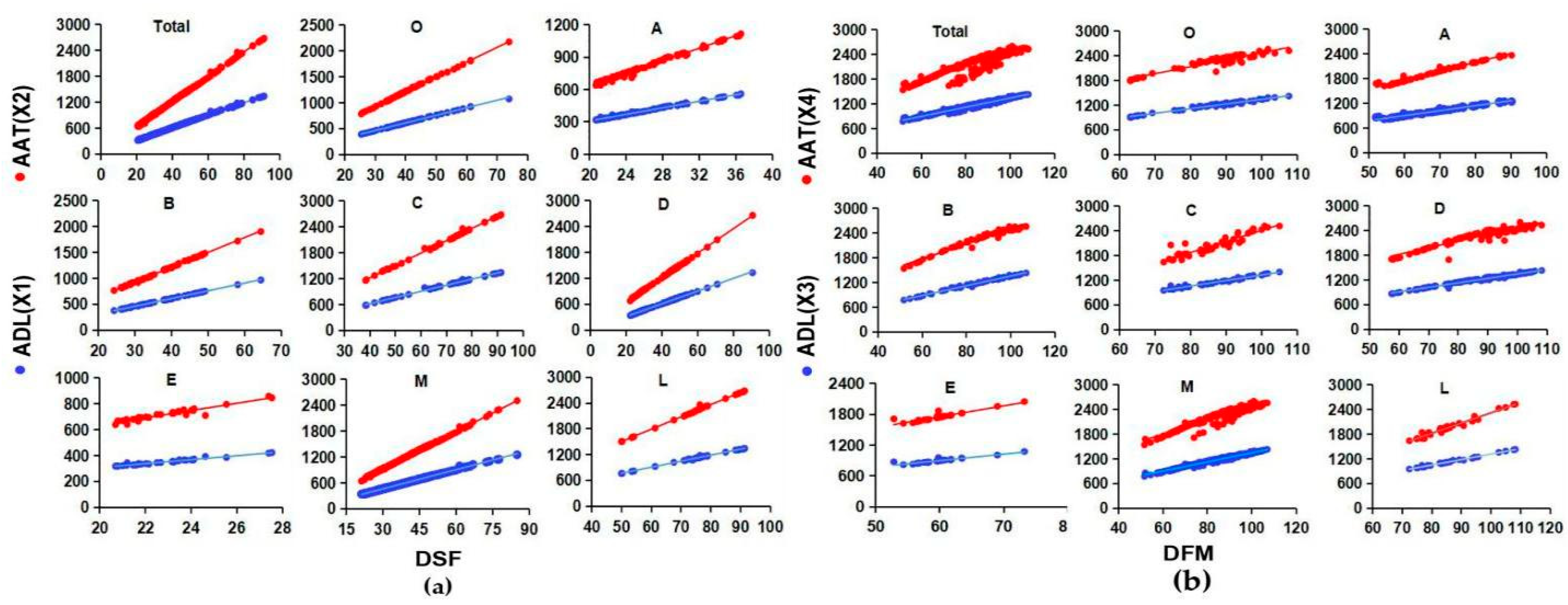
3.1. Wide Variation of ADL and AAT Required for DSF and DTM in Global Soybeans
3.2. Evolutionary Changes from the Center of Origin to Various Geographic Regions in ADL and AAT Required for Growth-Period Traits in Global Soybeans
3.3. Evolutionary Changes from the Primary MG Set to Early and Late MG Sets in ADL and AAT Required for Growth-Period Traits in Global Soybeans
3.4. Relative Importance of ADL, AAT and ADL×AAT in Determining Growth Periods of Geographic and MG Subpopulations
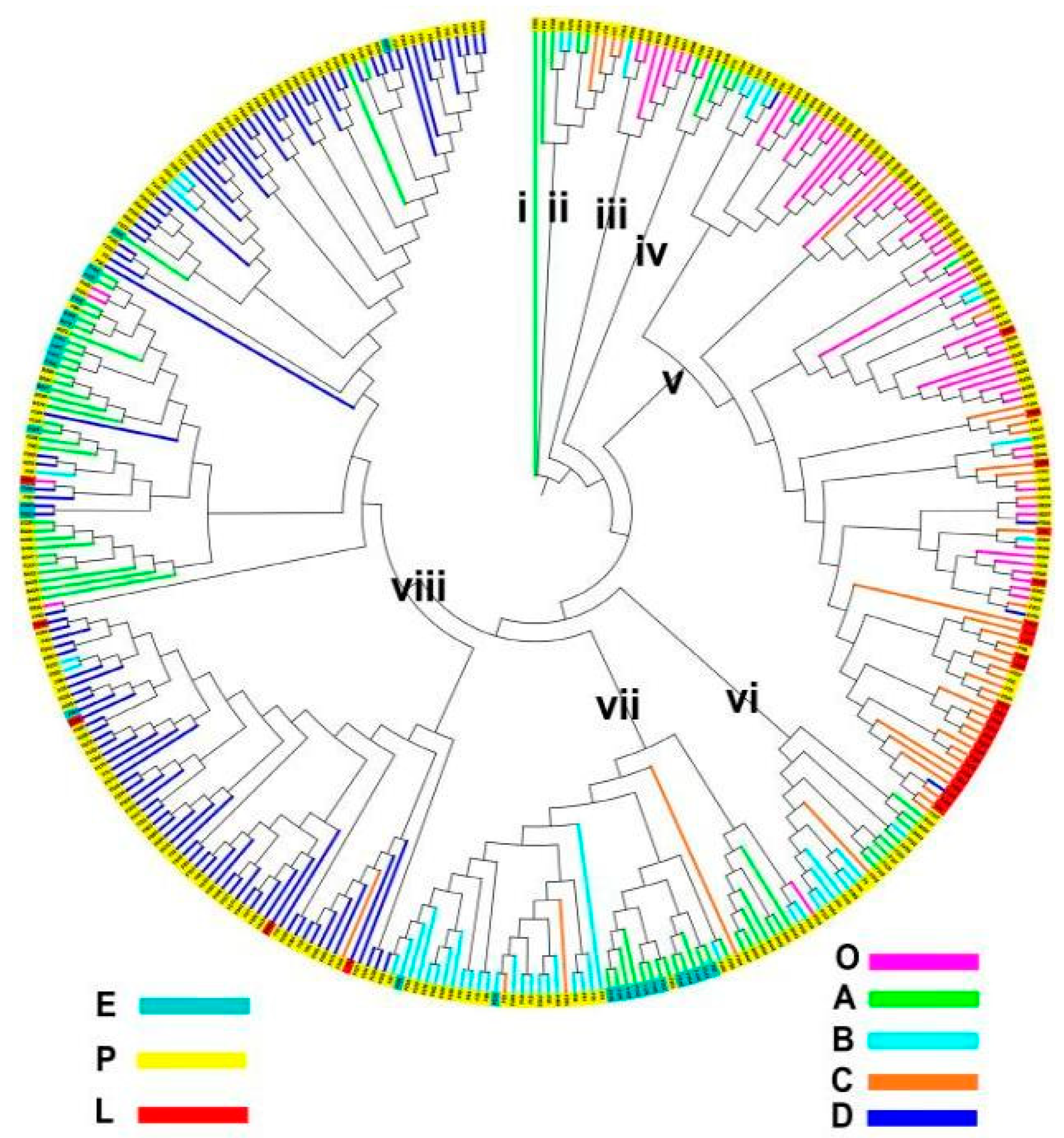
3.5. Genetic Clustering of the Global Soybeans and ADL and AAT Variation among and within the Clusters
4. Discussion
4.1. Separating Phenological Traits into Degree and Duration of Accumulative Day Length and Accumulative Active Temperature (ADL and AAT) Helped to Understand Directly the Functions of Eco-Factors in Geographic Dissemination Process
4.2. The Relative Importance of ADL, AAT and ADL×AAT in Determining Growth Period Traits and the Understanding of ADL × AAT Function
4.3. Responses of Growth Periods to ADL and AAT in Genotypic Clusters
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gaynor, L.; Lawn, R.; James, A. Agronomic studies on irrigated soybean in southern New South Wales. I. Phenological adaptation of genotypes to sowing date. Crop. Pasture Sci. 2012, 62, 1056–1066. [Google Scholar] [CrossRef]
- Garner, W.W. Comparative responses of long-day and short-day plants to relative length of day and night. Plant Physiol. 1933, 8, 347–356. [Google Scholar] [CrossRef] [Green Version]
- Garner, W.W.; Allard, H.A. Further studies in photoperiodism. The response of the plant to relative length of day and night. J. Agric. Res. 1923, 23, 871–920. [Google Scholar] [CrossRef] [Green Version]
- Garner, W.W.; Allard, H.A. Effect of the relative length of day and night and other factors of the environment on growth and reproduction in plants. J. Agric. Res. 1920, 18, 553–606. [Google Scholar] [CrossRef]
- Song, Y.H.; Ito, S.; Imaizumi, T. Flowering time regulation: Photoperiod- and temperature-sensing in leaves. Trends Plant Sci. 2013, 18, 575–583. [Google Scholar] [CrossRef] [Green Version]
- Newell, T.; Hymowitzc, A. Taxonomy of the genus Glycine, domestication and uses of soybeans. Econ. Bot. 1981, 35, 272–288. [Google Scholar]
- Watanabe, S.; Harada, K.; Abe, J. Genetic and molecular bases of photoperiod responses of flowering in soybean. Breed. Sci. 2012, 61, 531–543. [Google Scholar] [CrossRef] [Green Version]
- Hartman, G.L.; West, E.D.; Herman, T.K. Crops that feed the World 2. Soybean—Worldwide production, use, and constraints caused by pathogens and pests. Food Secur. 2011, 3, 5–17. [Google Scholar] [CrossRef]
- Liu, X.; He, J.; Wang, Y.; Xing, G.; Li, Y.; Yang, S.; Zhao, T.; Gai, J. Geographic differentiation and phylogeographic relationships among world soybean populations. Crop J. 2020, 8, 260–272. [Google Scholar] [CrossRef]
- Wen, Z.; Zhao, T.; Ding, Y.; Gai, J. Genetic diversity, geographic differentiation and evolutionary relationship among ecotypes of Glycine max and G. soja in China. Sci. Bull. 2009, 54, 4393–4403. [Google Scholar] [CrossRef] [Green Version]
- Zhao, T.; Gai, J. The Origin and Evolution of Cultivated Soybean [Glycine max (L.) Merr.]. Sci. Agric. Sin. 2004, 37, 954–962. [Google Scholar]
- Gai, J.Y.; Xu, D.H.; Gao, Z.; Shimamoto, Y.; Kitajima, S. Studies on the evolutionary relationship among eco-types of G. max and G. soja in China. Acta Agron. Sin. 2000, 26, 513–520. [Google Scholar]
- Hartwig, E.E.; Caldwell, B.E. Varietal development. In Soybeans: Improvement, Production, and Uses; Caldwell, B.E., Ed.; American Society of Agronomy: Madison, MI, USA, 1973. [Google Scholar]
- Norman, A.G. Soybean Physiology, Agronomy, and Utilization; Academic Press: Cambridge, MA, USA, 1978. [Google Scholar]
- Gaynor, L.G.; Lawn, R.J.; James, A.T. Agronomic studies on irrigated soybean in southern New South Wales. I. Phenological adaptation of genotypes to sowing date. Crop Pasture Ence 2011, 62, 1067–1077. [Google Scholar] [CrossRef]
- Han, T.; Wu, C.; Tong, Z.; Mentreddy, R.S.; Tan, K.; Gai, J. Postflowering photoperiod regulates vegetative growth and reproductive development of soybean. Environ. Exp. Bot. 2006, 55, 120–129. [Google Scholar] [CrossRef]
- Cober, E.R.; Stewart, D.W.; Voldeng, H.D. Photoperiod and Temperature Responses in Early-Maturing, Near-Isogenic Soybean Lines. Crop Sci. 2001, 41, 721–727. [Google Scholar] [CrossRef]
- Morandi, E.N.; Casano, L.M.; Reggiardo, L.M. Post-flowering photoperiodic effect on reproductive efficiency and seed growth in soybean. Field Crop. Res. 1988, 18, 227–241. [Google Scholar] [CrossRef]
- Wang, Z.; Reddy, V.R.; Acock, M.C. Testing for Early Photoperiod Insensitivity in Soybean. Agron. J. 1998, 90, 389–392. [Google Scholar] [CrossRef] [Green Version]
- Borthwick, H.A.; Parker, M.W. Effectiveness of photoperiodic treatments of plants of different age. Jpn. J. Crop Ence 1938, 100, 245–249. [Google Scholar] [CrossRef]
- Cober, E.R.; Molnar, S.J.; Charette, M.; Voldeng, H.D. A new locus for early maturity in soybean. Crop Sci. 2010, 50, 524–527. [Google Scholar] [CrossRef]
- Li, X.M.; Wu, C.X.; Ma, Q.B. Morphology and anatomy of the differentiation of flower bud and the process of flowering reversion in soybean cv. Zigongdongdou. Acta Agron. Sin. 2005, 31, 1437–1442. [Google Scholar]
- Hamner, K.K.; Nanda, K.C. Investigations on the effect of “light break” on the nature of the endogenous rhythm in the flowering response of biloxi soybean (Glycine, max, L. Merr.). Planta 1962, 58, 164–174. [Google Scholar] [CrossRef]
- Tomasz, P.I.; Steven, K.M. The Effect of Grafting on the Flowering of Near-Isogenic Lines of Soybean. Crop Sci. 2003, 43, 1760–1763. [Google Scholar] [CrossRef]
- Shanmugasundaram, S.; Wang, C.C.; Toung, T.S. Photoperiodic Response of Flowering in Two-Branched Soybean Plants. Bot. Gaz. 1979, 140, 4141–4147. [Google Scholar] [CrossRef]
- Liu, X. A Study on Geographic Differentiation, Genetic Dissection and Phylogenetic Relationship of the World Soybean. Ph.D. Thesis, Nanjing Agricultural University, Nanjing, China, 2015. [Google Scholar]
- Liu, X.; Wu, J.A.; Ren, H.; Qi, Y.; Li, C.; Cao, J.; Zhang, X.; Zhang, Z.; Cai, Z.; Gai, J. Genetic variation of world soybean maturity date and geographic distribution of maturity groups. Breed. Sci. 2017, 67, 221–232. [Google Scholar] [CrossRef] [Green Version]
- Fehr, W.R.; Caviness, C.E. Stages of soybean development. In Special Report 80, Cooperative Extension Service, Agriculture and Home Economic Experiment Station; Iowa State University: Ames, IA, USA, 1977; pp. 1–11. [Google Scholar]
- Nyquist, W.E.; Baker, R.J. Estimation of heritability and prediction of selection response in plant populations. Crit. Rev. Plant Sci. 1991, 10, 235–322. [Google Scholar] [CrossRef]
- Liu, X.; Li, C.; Cao, J.; Zhang, X.; Wang, C.; He, J.; Xing, G.; Wang, W.; Zhao, J.; Gai, J. Growth period QTL-allele constitution of global soybeans and its differential evolution changes in geographic adaptation versus maturity group extension. Plant J. 2021, 108, 1624–1643. [Google Scholar] [CrossRef]
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef] [Green Version]
- Wall, J.D.; Pritchard, J.K. Haplotype blocks and linkage disequilibrium in the human genome. Nat. Rev. Genet. 2003, 4, 587–597. [Google Scholar] [CrossRef]
- He, J.; Meng, S.; Zhao, T.; Xing, G.; Yang, S.; Li, Y.; Guan, R.; Lu, J.; Wang, Y.; Xia, Q.; et al. An innovative procedure of genome-wide association analysis fits studies on germplasm population and plant breeding. Theor. Appl. Genet. 2017, 130, 2327–2343. [Google Scholar] [CrossRef]
- Lewis, P.O.; Kumar, S.; Tamura, K.; Nei, M. MEGA: Molecular Evolutionary Genetics Analysis, Version 1.02. Syst. Biol. 1995, 44, 576–577. [Google Scholar] [CrossRef]
- Aiken, L.S.; West, S.G. Multiple Regression: Testing and Interpreting Interactions; Sage: New York, NY, USA, 1991. [Google Scholar]
- Setiyono, T.D.; Weiss, A.; Specht, J.; Bastidas, A.M.; Cassman, K.G.; Dobermann, A. Understanding and modeling the effect of temperature and daylength on soybean phenology under high-yield conditions. Field Crop. Res. 2007, 100, 257–271. [Google Scholar] [CrossRef] [Green Version]
- Choi, D.H.; Ban, H.Y.; Seo, B.S.; Lee, K.J.; Lee, B.W. Phenology and Seed Yield Performance of Determinate Soybean Cultivars Grown at Elevated Temperatures in a Temperate Region. PLoS ONE 2016, 11, e0165977. [Google Scholar] [CrossRef]
- Tan, Q.; Liu, Y.; Dai, L.; Pan, T. Shortened key growth periods of soybean observed in China under climate change. Sci. Rep. 2021, 11, 8197. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Trait | Geo-Pop. | Class Midpoint Value | N | Mean | GCV (%) | Range | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DSF (d) | (PropDSF %) | 23.5 | 30.5 | 37.5 | 44.5 | 51.5 | 58.5 | 65.5 | 72.5 | 79.5 | 86.5 | >90 | ||||
| O (32.2 B) | 3 | 4 | 31 | 12 | 8 | 4 | 2 | 0 | 1 | 0 | 0 | 65 | 41.9 B | 22.8 | 25.6–77.0 | |
| A (28.6 D) | 47 | 20 | 8 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 75 | 26.4 C | 16.9 | 20.6–37.1 | |
| B (31.0 BC) | 4 | 15 | 15 | 10 | 5 | 1 | 1 | 0 | 0 | 0 | 0 | 51 | 38.1 B | 22.8 | 24.3–66.6 | |
| C (44.2 A) | 0 | 0 | 2 | 5 | 4 | 1 | 5 | 6 | 15 | 2 | 5 | 45 | 70.6 A | 22.9 | 39.0–95.5 | |
| D (30.2 C) | 18 | 37 | 4 | 26 | 22 | 7 | 1 | 1 | 0 | 0 | 1 | 117 | 39.4 B | 32.5 | 22.6–94.8 | |
| All (32.1) | 72 | 76 | 60 | 53 | 39 | 13 | 9 | 7 | 16 | 2 | 6 | 353 | 41.0 | 40.7 | 20.6–95.5 | |
| ADLDSF (d·h) | (PDLDSF %) | <320 | 370 | 470 | 570 | 670 | 770 | 870 | 970 | 1070 | 1170 | >1220 | ||||
| O (34.2 B) | 0 | 3 | 10 | 26 | 12 | 8 | 3 | 2 | 1 | 0 | 0 | 65 | 621.9 B | 22.0 | 388.4–1098.8 | |
| A (29.9 C) | 5 | 46 | 19 | 5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 75 | 397.5 C | 16.3 | 311.7–552.5 | |
| B (32.9 B) | 0 | 6 | 16 | 14 | 9 | 4 | 1 | 1 | 0 | 0 | 0 | 51 | 565.9 B | 22.3 | 368.6–975.7 | |
| C (47.7 A) | 0 | 0 | 0 | 2 | 6 | 3 | 1 | 5 | 6 | 15 | 7 | 45 | 1035.4 A | 22.2 | 579.0–1382.3 | |
| D (32.2 B) | 0 | 25 | 30 | 6 | 25 | 25 | 3 | 1 | 1 | 0 | 1 | 117 | 584.9 B | 31.6 | 336.8–1372.7 | |
| All (34.1) | 5 | 80 | 75 | 53 | 52 | 40 | 8 | 9 | 8 | 15 | 8 | 353 | 606.6 | 39.6 | 311.7–1382.3 | |
| AATDSF (d·°C) | (PATDSF%) | <650 | 760 | 980 | 1200 | 1420 | 1640 | 1860 | 2080 | 2300 | 2520 | >2630 | ||||
| O (34.5 B) | 0 | 5 | 14 | 27 | 12 | 4 | 2 | 0 | 1 | 0 | 0 | 65 | 1211.1 B | 21.8 | 763.5–2202.8 | |
| A (30.3 C) | 6 | 51 | 18 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 75 | 785.4 C | 15.5 | 622.4–1077.6 | |
| B (33.2 B) | 0 | 9 | 15 | 17 | 8 | 1 | 1 | 0 | 0 | 0 | 0 | 51 | 1105.5 B | 21.6 | 734.5–1883.9 | |
| C (49.8 A) | 0 | 0 | 0 | 3 | 8 | 1 | 5 | 7 | 14 | 3 | 4 | 45 | 2017.8 A | 22.8 | 1125.4–2726.2 | |
| D (32.8 B) | 0 | 39 | 18 | 12 | 36 | 9 | 1 | 1 | 0 | 0 | 1 | 117 | 1143.7 B | 30.6 | 661.3–2708.3 | |
| All (34.8) | 6 | 104 | 65 | 59 | 64 | 15 | 9 | 8 | 15 | 3 | 5 | 353 | 1185.9 | 39.1 | 622.6–2726.2 | |
| DFM (d) | (PropDFM %) | 46.5 | 53.5 | 60.5 | 67.5 | 74.5 | 81.5 | 88.5 | 95.5 | 102.5 | 109.5 | 116.5 | ||||
| O (67.8 C) | 0 | 0 | 8 | 3 | 3 | 7 | 12 | 20 | 9 | 2 | 1 | 65 | 88.1 A | 15.9 | 60.1–114.1 | |
| A (71.4 A) | 2 | 20 | 12 | 18 | 8 | 9 | 5 | 1 | 0 | 0 | 0 | 75 | 66.5 B | 18.1 | 47.8–93.4 | |
| B (68.9 BC) | 2 | 4 | 2 | 3 | 6 | 7 | 6 | 7 | 7 | 7 | 0 | 51 | 85.1 A | 21.4 | 45.8–112.8 | |
| C (55.8 D) | 0 | 0 | 0 | 1 | 8 | 11 | 9 | 7 | 6 | 3 | 0 | 45 | 87.8 A | 12.7 | 70.3–111.1 | |
| D (69.8 B) | 0 | 4 | 4 | 14 | 8 | 15 | 14 | 21 | 22 | 14 | 1 | 117 | 88.7 A | 17.3 | 52.1–114.6 | |
| All (67.9) | 4 | 28 | 26 | 39 | 33 | 49 | 46 | 56 | 44 | 26 | 2 | 353 | 83.3 | 20.2 | 45.8–114.6 | |
| ADLDFM (d·h) | (PDLDFM %) | <720 | 760 | 840 | 920 | 1000 | 1080 | 1160 | 1240 | 1320 | 1400 | >1440 | ||||
| O (65.8 B) | 0 | 0 | 7 | 4 | 0 | 3 | 12 | 19 | 11 | 8 | 1 | 65 | 1191.9 A | 14.2 | 834.0–1479.4 | |
| A (70.1 A) | 1 | 20 | 11 | 8 | 15 | 8 | 6 | 6 | 0 | 0 | 0 | 75 | 936.2 B | 16.9 | 685.0–1274.8 | |
| B (67.1 B) | 2 | 3 | 3 | 1 | 5 | 8 | 4 | 7 | 6 | 9 | 3 | 51 | 1159.1 A | 19.2 | 652.0–1476.4 | |
| C (52.3 C) | 0 | 0 | 2 | 5 | 8 | 12 | 4 | 3 | 6 | 4 | 1 | 45 | 1120.8 A | 14.7 | 859.5–1458.0 | |
| D (67.8 B) | 0 | 3 | 2 | 12 | 6 | 10 | 19 | 16 | 26 | 19 | 4 | 117 | 1200.6 A | 14.9 | 746.8–1487.4 | |
| All (65.9) | 3 | 26 | 25 | 30 | 34 | 41 | 45 | 51 | 49 | 40 | 9 | 353 | 1126.7 | 18.2 | 652.0–1487.4 | |
| AATDFM (d·°C) | (PATDFM %) | <1400 | 1470 | 1610 | 1750 | 1890 | 2030 | 2170 | 2310 | 2450 | 2590 | >2660 | ||||
| O (65.5 B) | 0 | 0 | 3 | 7 | 2 | 3 | 5 | 14 | 15 | 12 | 4 | 65 | 2289.7 A | 13.0 | 1661.1–2694.1 | |
| A (69.7 A) | 3 | 17 | 10 | 7 | 14 | 9 | 7 | 2 | 6 | 0 | 0 | 75 | 1814.7 C | 17.6 | 1339.8–2477.6 | |
| B (66.8 B) | 2 | 2 | 3 | 1 | 4 | 6 | 5 | 6 | 6 | 10 | 6 | 51 | 2228.1 A | 18.0 | 1251.5–2732.9 | |
| C (50.2 C) | 0 | 3 | 6 | 6 | 8 | 5 | 6 | 0 | 5 | 4 | 2 | 45 | 2016.9 B | 18.3 | 1437.5–2715.7 | |
| D (67.2 B) | 0 | 5 | 1 | 7 | 10 | 6 | 13 | 17 | 31 | 19 | 8 | 117 | 2289.8 A | 14.0 | 1449.1–2755.2 | |
| All (65.2) | 5 | 27 | 23 | 28 | 38 | 29 | 36 | 39 | 63 | 45 | 20 | 353 | 2145.1 | 17.9 | 1251.5–2755.2 | |
| Trait | MG | Class Midpoint Value(Frequency) | N | Mean | GCV (%) | Range | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DSF (d) | (PropDSF%) | 23.5 | 30.5 | 37.5 | 44.5 | 51.5 | 58.5 | 65.5 | 72.5 | 79.5 | 86.5 | >90 | ||||
| E (29.3 B) | 29 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 31 | 23.0 C | 8.22 | 20.6–27.7 | |
| P (30.9 B) | 43 | 74 | 60 | 53 | 36 | 12 | 8 | 2 | 3 | 1 | 0 | 292 | 39.1 B | 30.70 | 22.6–88.6 | |
| L (46.9 A) | 0 | 0 | 0 | 0 | 3 | 1 | 1 | 5 | 13 | 1 | 6 | 30 | 77.5 A | 16.06 | 51.0–95.5 | |
| All (32.1) | 72 | 76 | 60 | 53 | 39 | 13 | 9 | 7 | 16 | 2 | 6 | 353 | 41.0 | 40.7 | 20.5–95.5 | |
| ADLDSF (d·h) | (PDLDSF%) | <320 | 370 | 470 | 570 | 670 | 770 | 870 | 970 | 1070 | 1170 | >1220 | ||||
| E (30.5 C) | 5 | 26 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 31 | 346.0 C | 7.7 | 311.7–418.9 | |
| P (32.8 B) | 0 | 54 | 75 | 53 | 52 | 36 | 8 | 8 | 2 | 3 | 1 | 292 | 580.5 B | 30.0 | 336.8–1285.2 | |
| L (50.7 A) | 0 | 0 | 0 | 0 | 0 | 4 | 0 | 1 | 6 | 12 | 7 | 30 | 1129.6 A | 15.7 | 751.7–1382.3 | |
| All (34.1) | 5 | 80 | 75 | 53 | 52 | 40 | 8 | 9 | 8 | 15 | 8 | 353 | 606.6 | 39.6 | 311.7–1382.3 | |
| AATDSF (d·°C) | (PETDSF%) | <650 | 760 | 980 | 1200 | 1420 | 1640 | 1860 | 2080 | 2300 | 2520 | >2630 | ||||
| E (31.4 B) | 5 | 26 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 31 | 691.3 C | 6.8 | 622.6–819.1 | |
| P (33.2 B) | 1 | 78 | 65 | 59 | 62 | 13 | 8 | 3 | 2 | 1 | 0 | 292 | 1132.8 B | 29.2 | 638.2–2539.0 | |
| L (53.6 A) | 0 | 0 | 0 | 0 | 2 | 2 | 1 | 5 | 13 | 2 | 5 | 30 | 2213.7 A | 16.2 | 1458.5–2726.2 | |
| All (34.8) | 6 | 104 | 65 | 59 | 64 | 15 | 9 | 8 | 15 | 3 | 5 | 353 | 1185.9 | 39.1 | 622.6–2726.2 | |
| DFM (d) | (PropDFM%) | 46.5 | 53.5 | 60.5 | 67.5 | 74.5 | 81.5 | 88.5 | 95.5 | 102.5 | 109.5 | 116.5 | ||||
| E (70.7 A) | 1 | 21 | 7 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 31 | 55.6 B | 8.5 | 47.8–71.5 | |
| P (69.1 A) | 3 | 7 | 19 | 37 | 26 | 42 | 39 | 51 | 44 | 24 | 0 | 292 | 85.7 A | 17.9 | 45.8–112.8 | |
| L (53.1 B) | 0 | 0 | 0 | 1 | 6 | 7 | 7 | 5 | 0 | 2 | 2 | 30 | 87.6 A | 14.3 | 70.3–114.6 | |
| All (67.9) | 4 | 28 | 26 | 39 | 33 | 49 | 46 | 56 | 44 | 26 | 2 | 353 | 83.3 | 20.2 | 45.8–114.6 | |
| ADLDFM (d·h) | (PDLDFM %) | <720 | 760 | 840 | 920 | 1000 | 1080 | 1160 | 1240 | 1320 | 1400 | >1440 | ||||
| E (69.5 A) | 1 | 20 | 8 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 31 | 791.5 B | 8.2 | 685.0–1006.8 | |
| P (67.2 B) | 2 | 6 | 15 | 24 | 28 | 34 | 41 | 48 | 49 | 38 | 7 | 292 | 1165.0 A | 15.7 | 652.0–1476.4 | |
| L (49.3 C) | 0 | 0 | 2 | 5 | 5 | 7 | 4 | 3 | 0 | 2 | 2 | 30 | 1099.7 A | 16.1 | 859.5–1487.4 | |
| All (65.9) | 3 | 26 | 25 | 30 | 34 | 41 | 45 | 51 | 49 | 40 | 9 | 353 | 1126.7 | 18.2 | 652.0–1487.4 | |
| AATDFM (d·°C) | (PETDFM %) | <1400 | 1470 | 1610 | 1750 | 1890 | 2030 | 2170 | 2310 | 2450 | 2590 | >2660 | ||||
| E (68.6 A) | 3 | 18 | 8 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 31 | 1514.9 C | 8.3 | 1340.0–1931.0 | |
| P (66.8 A) | 2 | 5 | 10 | 24 | 29 | 27 | 33 | 38 | 63 | 43 | 18 | 292 | 2235.1 A | 14.8 | 1252.0–2755.0 | |
| L (46.4 B) | 0 | 4 | 5 | 4 | 7 | 2 | 3 | 1 | 0 | 2 | 2 | 30 | 1920.6 B | 19.0 | 1437.0–2700.0 | |
| All (65.2) | 5 | 27 | 23 | 28 | 38 | 29 | 36 | 39 | 63 | 45 | 20 | 353 | 2145.1 | 17.9 | 1251.5–2755.2 | |
| Trait | Source of Variation | Total | Sub-Population | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| O | A | B | C | D | E | P | L | |||
| DSF | Model R2 | 99.90 | 99.9 | 99.7 | 99.9 | 99.8 | 99.9 | 98.7 | 99.9 | 99.6 |
| ADLDSF (x1) | 38.05 | 29.33 | 67.51 | 84.79 | 0.54 | 44.66 | 71.35 | 37.24 | 1.43 | |
| AATDSF (x2) | 44.79 | 15.38 | 23.65 | 0.23 | 97.56 | 19.92 | 27.36 | 54.30 | 84.74 | |
| (x1 x2)DSF | 17.10 | 55.25 | 8.55 | 14.89 | 1.66 | 35.37 | 0.01 | 8.37 | 13.48 | |
| Residual | 0.06 | 0.04 | 0.29 | 0.09 | 0.24 | 0.05 | 1.28 | 0.09 | 0.35 | |
| DFM | Model R2 | 98.90 | 98.7 | 98.3 | 99.4 | 98.6 | 99.5 | 88.9 | 98.8 | 99.2 |
| ADLDFM (x3) | 86.98 | 88.69 | 1.57 | 71.87 | 93.24 | 82.3 | 2.53 | 78.64 | 93.98 | |
| AATDFM (x4) | 11.42 | 9.57 | 34.44 | 27.43 | 4.25 | 16.26 | 39.08 | 19.85 | 4.76 | |
| (x3 x4)DFM | 0.54 | 0.52 | 62.30 | 0.07 | 1.07 | 0.97 | 47.25 | 0.29 | 0.46 | |
| Residual | 1.06 | 1.23 | 1.69 | 0.63 | 1.43 | 0.48 | 11.14 | 1.22 | 0.8 | |
| Genetic Distance Cluster | Geographic Subpopulation | MG-Set Subpopulation | Total | ||||||
|---|---|---|---|---|---|---|---|---|---|
| O | A | B | C | D | E | P | L | ||
| i | 1 | 1 | 1 | ||||||
| ii | 4 | 2 | 4 | 10 | 10 | ||||
| iii | 7 | 1 | 8 | 8 | |||||
| iv | 1 | 5 | 6 | 6 | |||||
| v | 52 | 3 | 8 | 37 | 4 | 79 | 25 | 104 | |
| vi | 8 | 8 | 1 | 17 | 17 | ||||
| vii | 1 | 21 | 26 | 3 | 1 | 14 | 38 | 52 | |
| viii | 4 | 33 | 6 | 1 | 112 | 17 | 134 | 5 | 156 |
| Total | 65 | 75 | 51 | 46 | 117 | 31 | 293 | 30 | 354 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, C.; Liu, X.; Hao, X.; Pan, Y.; Zong, C.; Zeng, W.; Wang, W.; Xing, G.; He, J.; Gai, J. Evolutionary Variation of Accumulative Day Length and Accumulative Active Temperature Required for Growth Periods in Global Soybeans. Agronomy 2022, 12, 962. https://doi.org/10.3390/agronomy12040962
Wang C, Liu X, Hao X, Pan Y, Zong C, Zeng W, Wang W, Xing G, He J, Gai J. Evolutionary Variation of Accumulative Day Length and Accumulative Active Temperature Required for Growth Periods in Global Soybeans. Agronomy. 2022; 12(4):962. https://doi.org/10.3390/agronomy12040962
Chicago/Turabian StyleWang, Can, Xueqin Liu, Xiaoshuai Hao, Yongpeng Pan, Chunmei Zong, Weiying Zeng, Wubin Wang, Guangnan Xing, Jianbo He, and Junyi Gai. 2022. "Evolutionary Variation of Accumulative Day Length and Accumulative Active Temperature Required for Growth Periods in Global Soybeans" Agronomy 12, no. 4: 962. https://doi.org/10.3390/agronomy12040962
APA StyleWang, C., Liu, X., Hao, X., Pan, Y., Zong, C., Zeng, W., Wang, W., Xing, G., He, J., & Gai, J. (2022). Evolutionary Variation of Accumulative Day Length and Accumulative Active Temperature Required for Growth Periods in Global Soybeans. Agronomy, 12(4), 962. https://doi.org/10.3390/agronomy12040962