
Soil Available Phosphorus Deficiency Reduces Boll Biomass and Lint Yield by Affecting Sucrose Metabolism in Cotton-Boll Subtending Leaves

Abstract
:1. Introduction
2. Materials and Methods
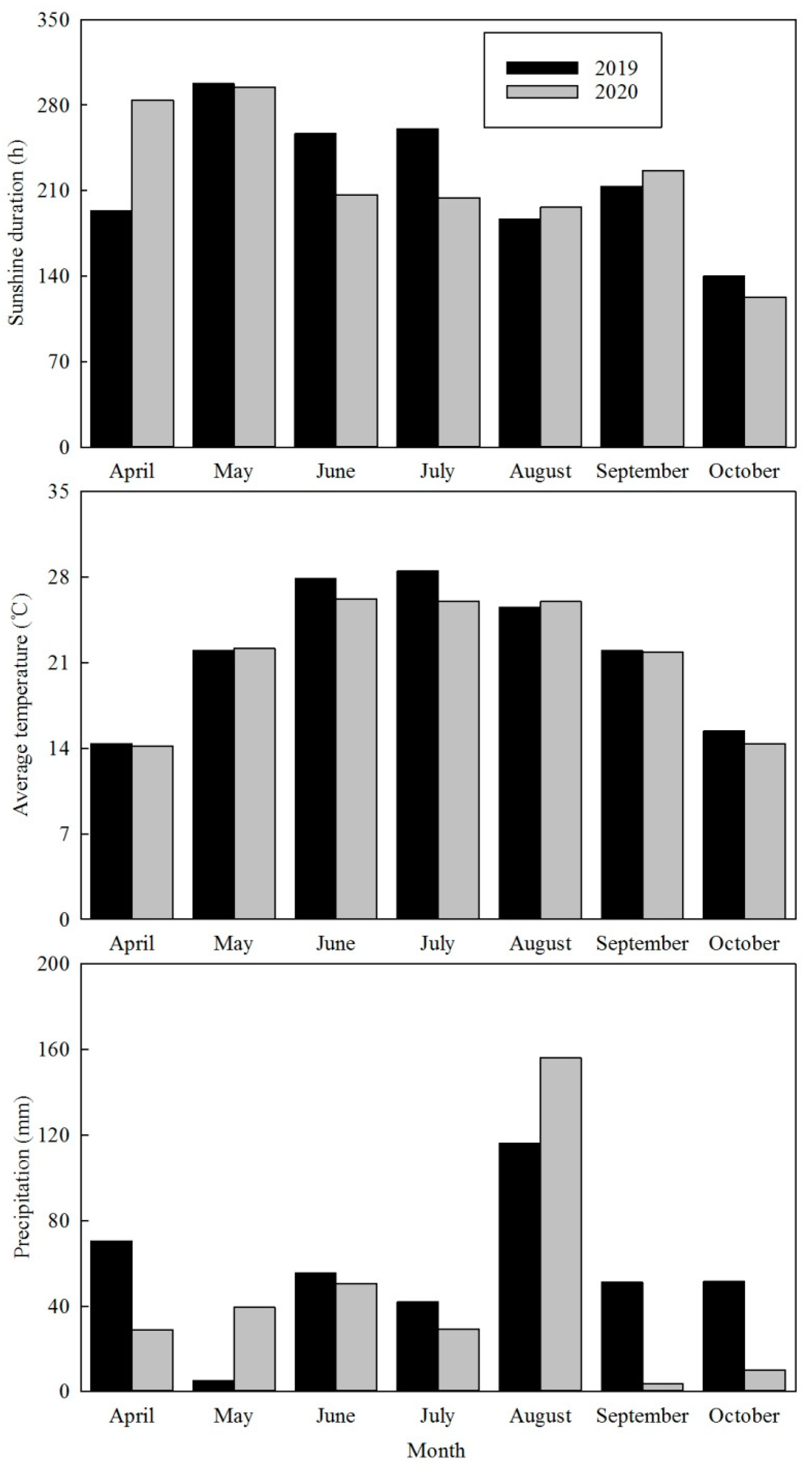
2.1. Description of Experimental Site
2.2. Experimental Design and Field Management
2.3. Sampling and Processing
2.4. Leaf Photosynthesis
2.5. Leaf Phosphorus Concentration and Carbohydrate Content
2.6. Analysis of Leaf Enzymes Activities
2.7. Statistical Analysis
3. Results
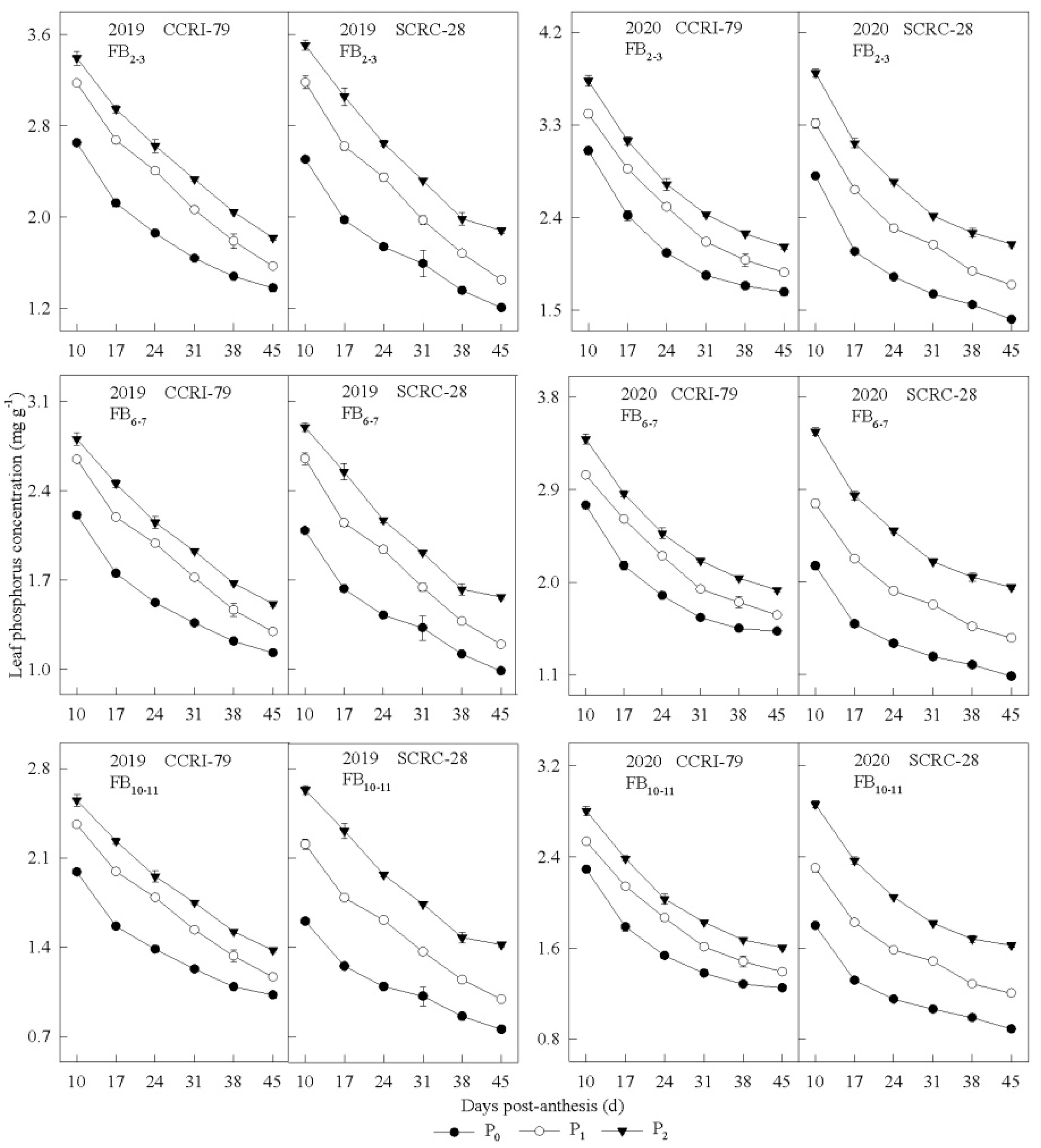
3.1. Phosphorus Concentration in the Subtending Leaves
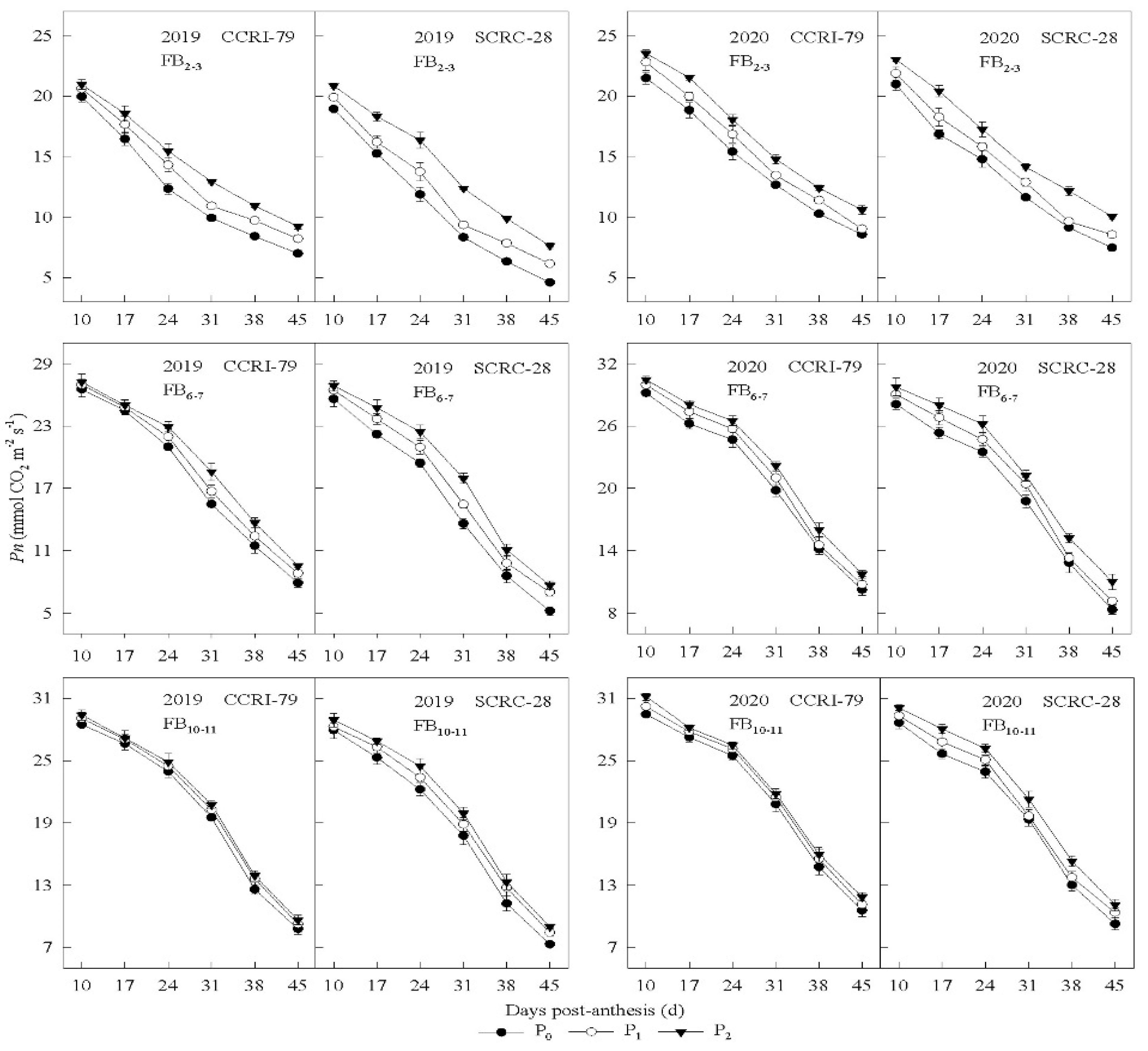
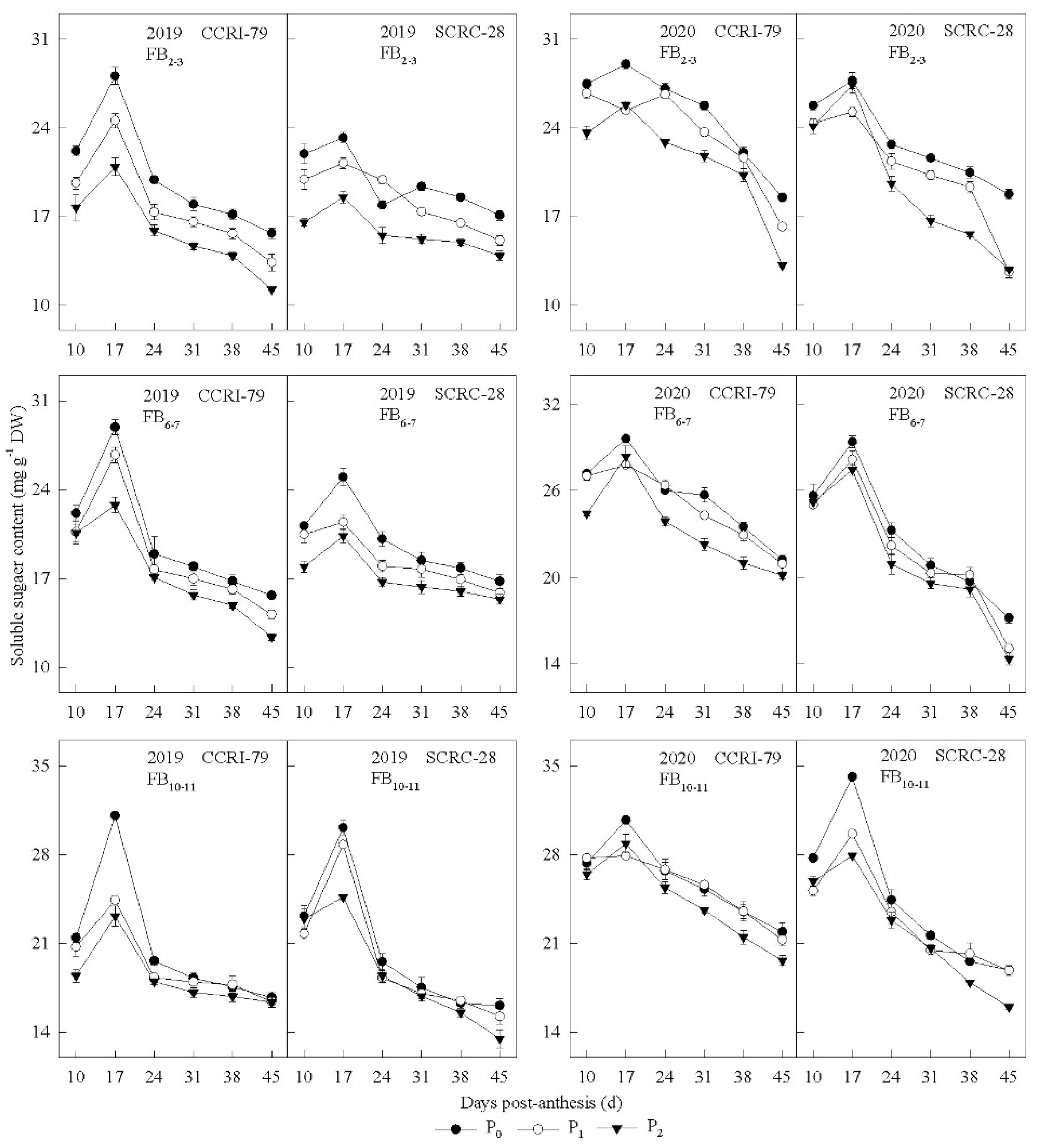
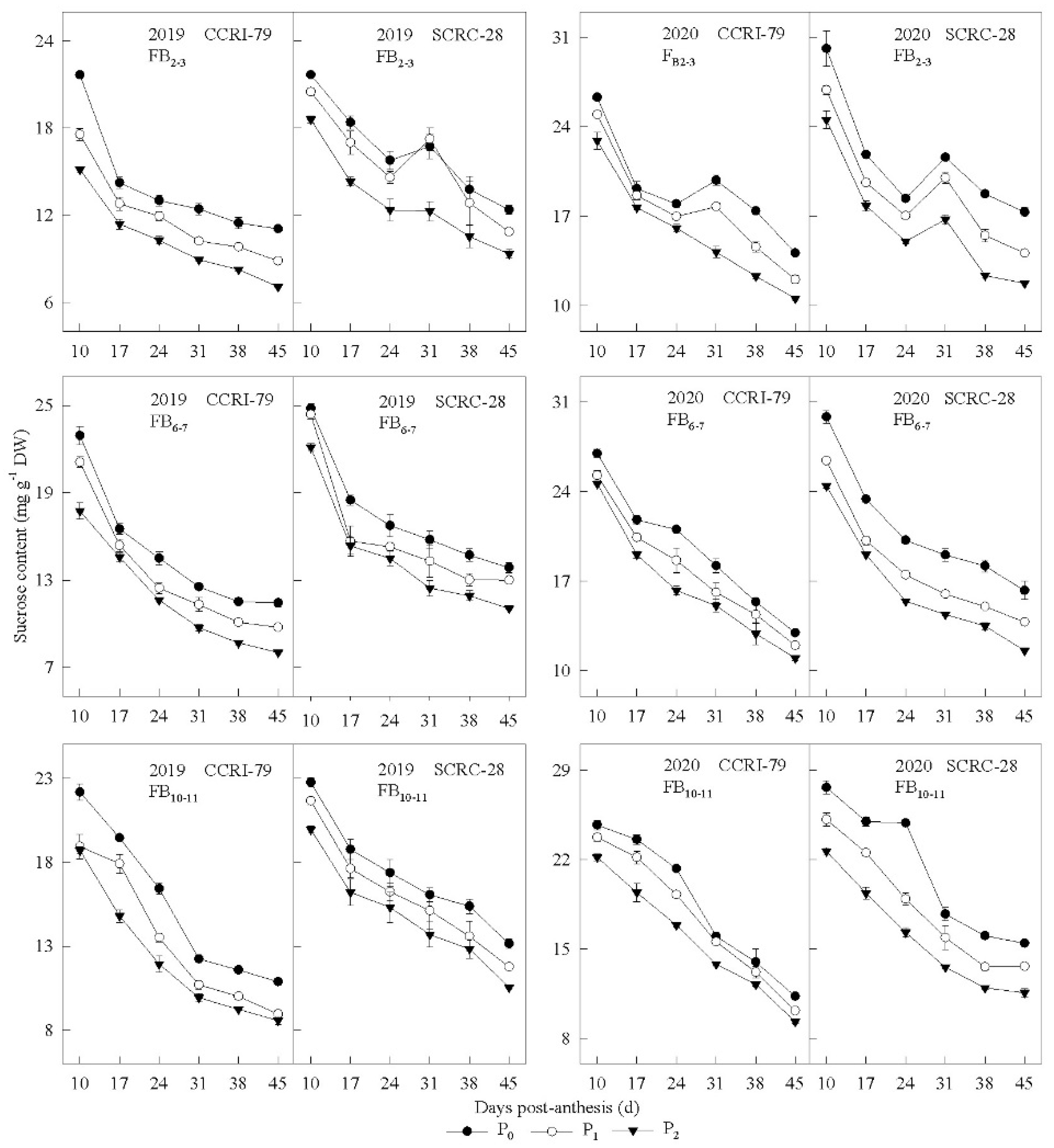
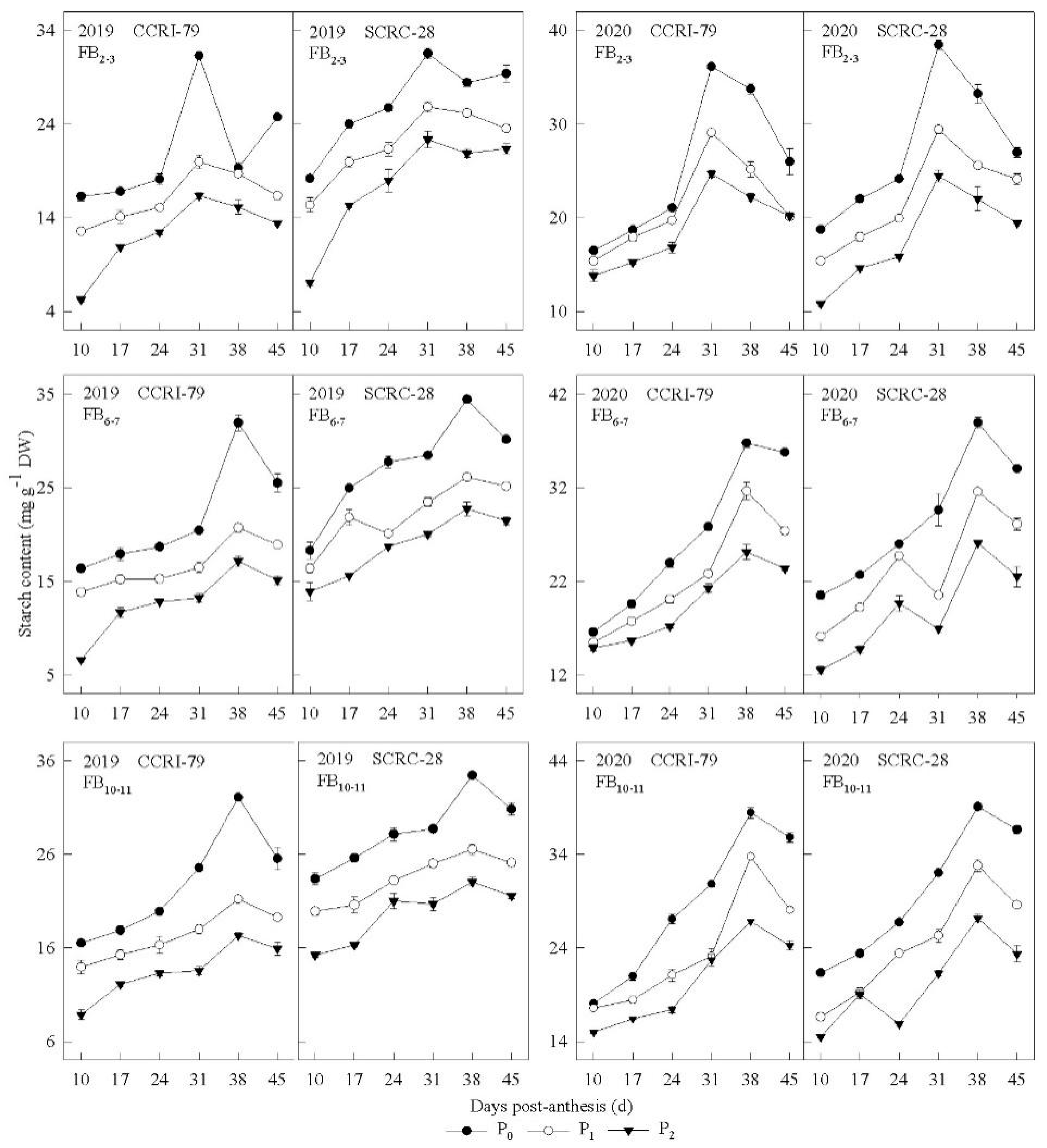
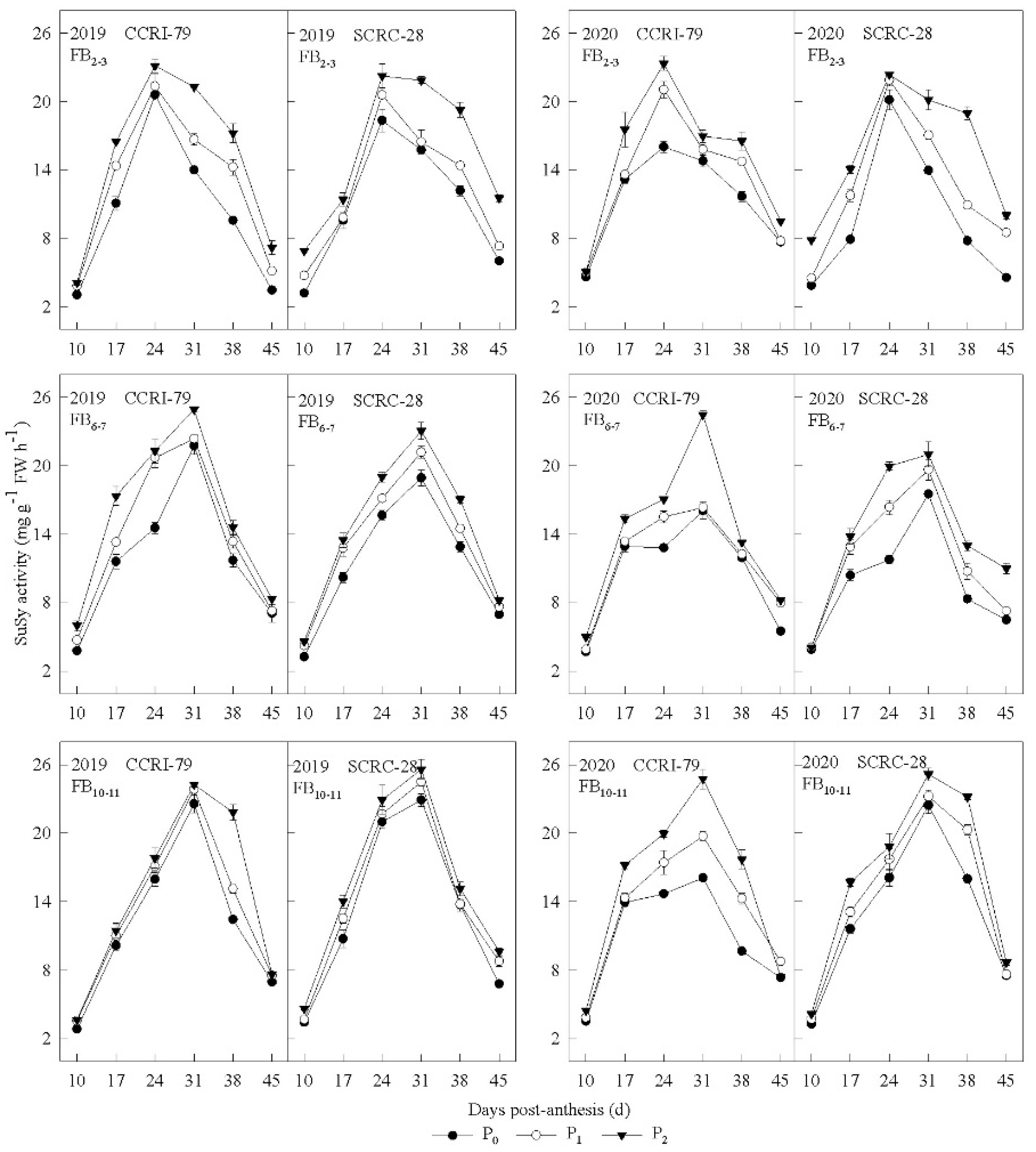
3.2. Pn and Carbohydrate Content in the Subtending Leaves
3.3. Correlation of the Carbohydrate Content and Phosphorus Concentration of Subtending Leaves
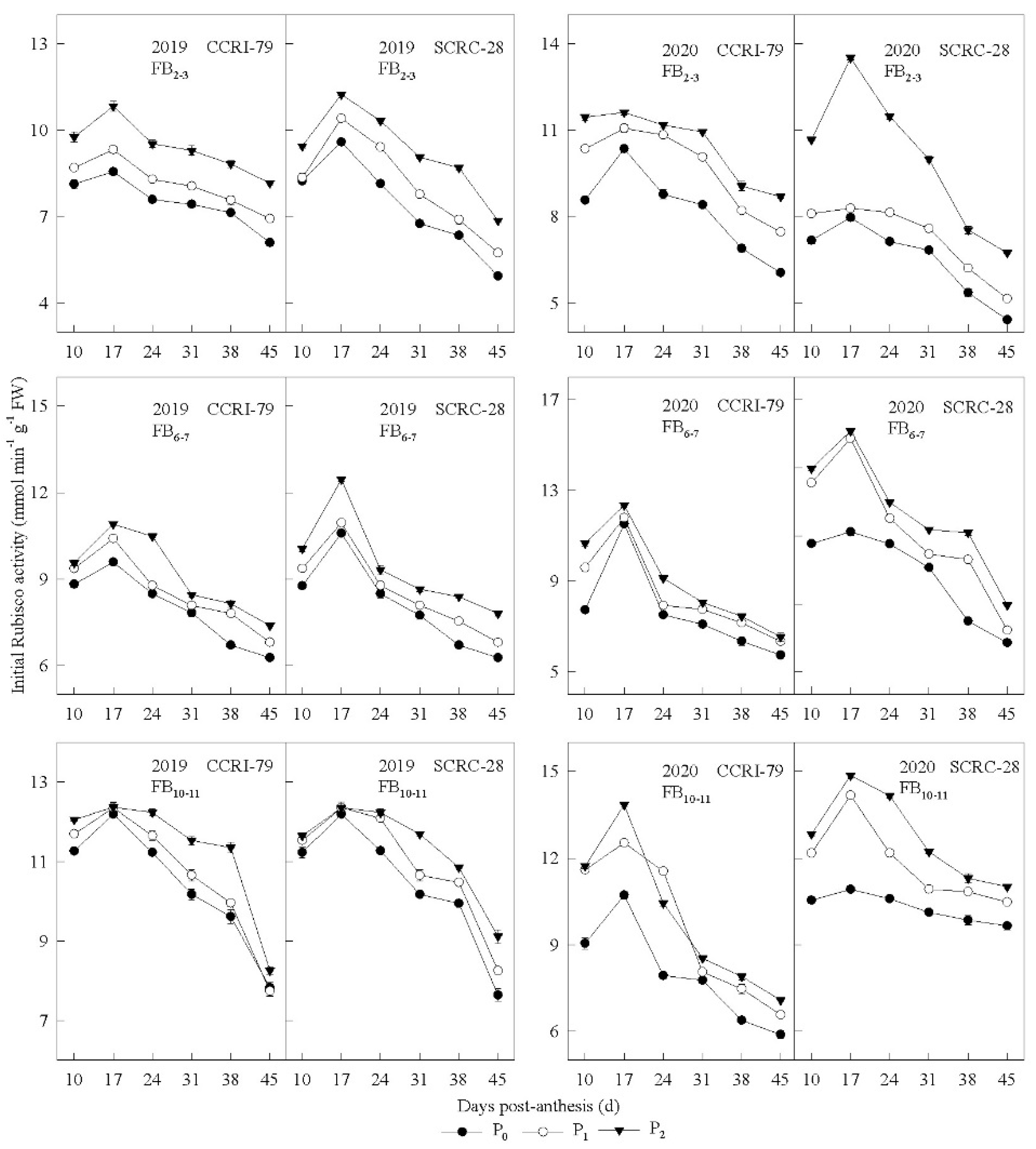
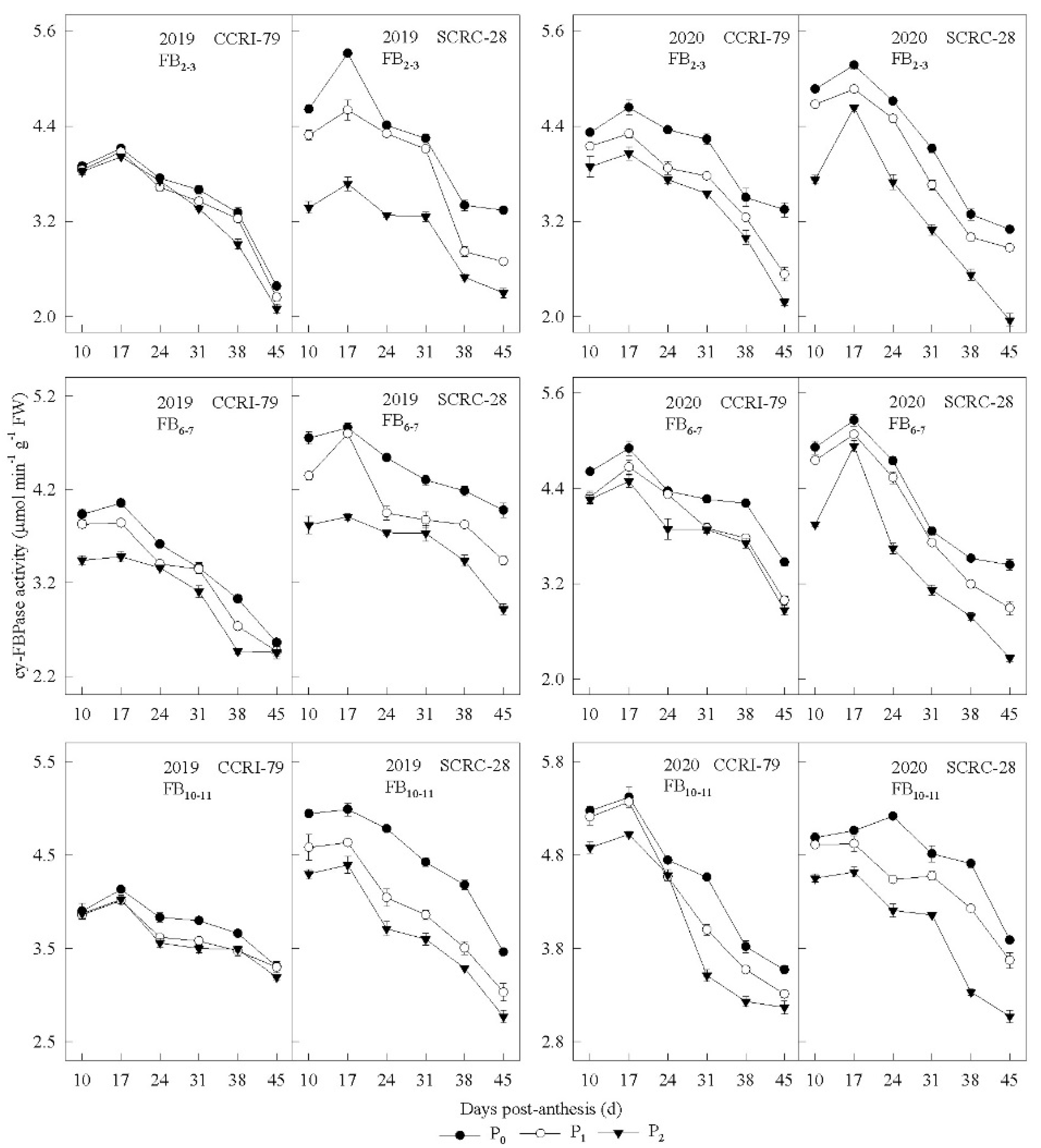
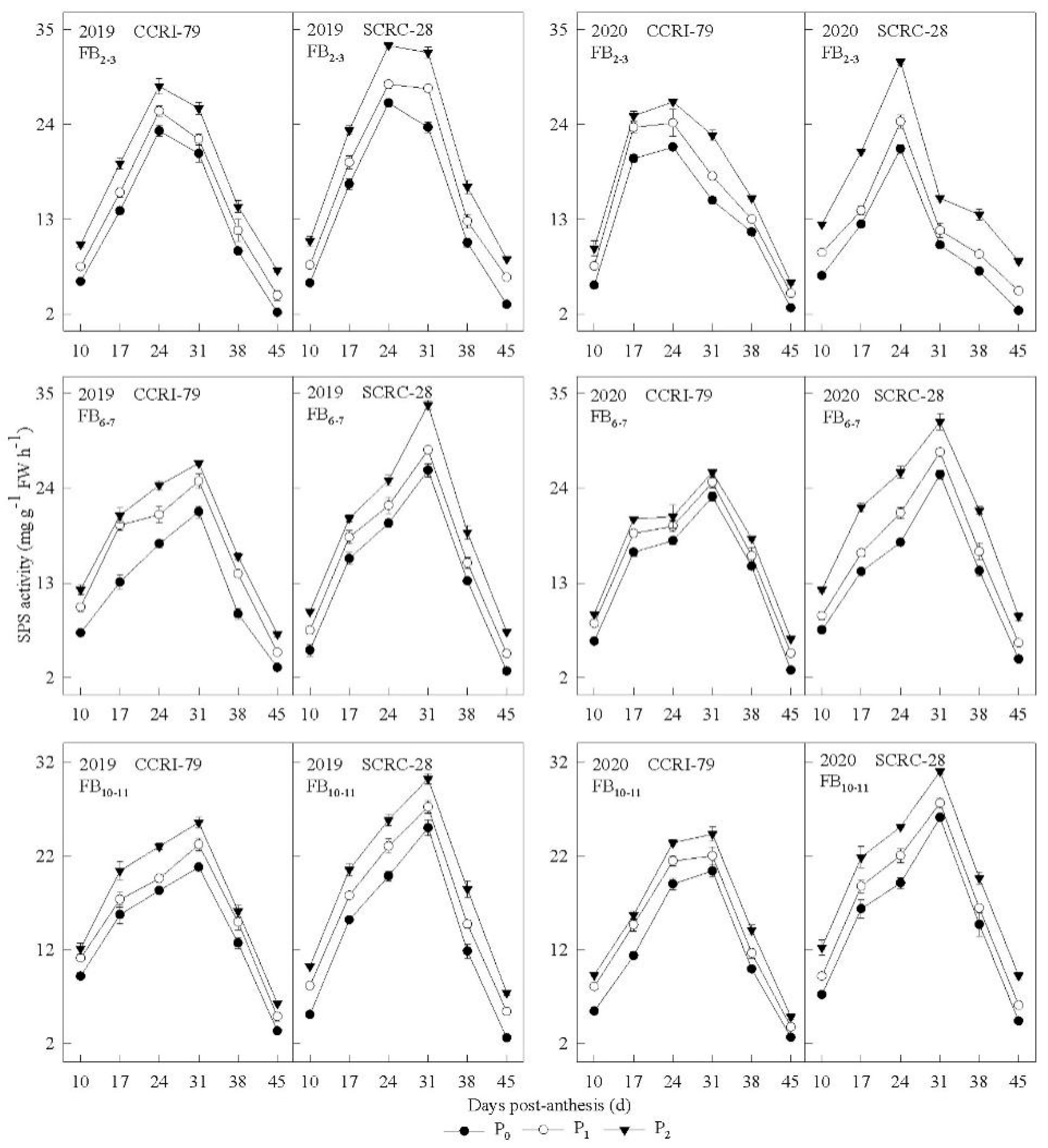
3.4. Sucrose-Metabolizing Enzyme Activities in the Subtending Leaves
3.5. Correlation of Sucrose-Metabolizing Enzyme Activity to the Phosphorus Concentration of Subtending Leaves
3.6. Lint Yield and Yield Components on Different Fruiting Branches
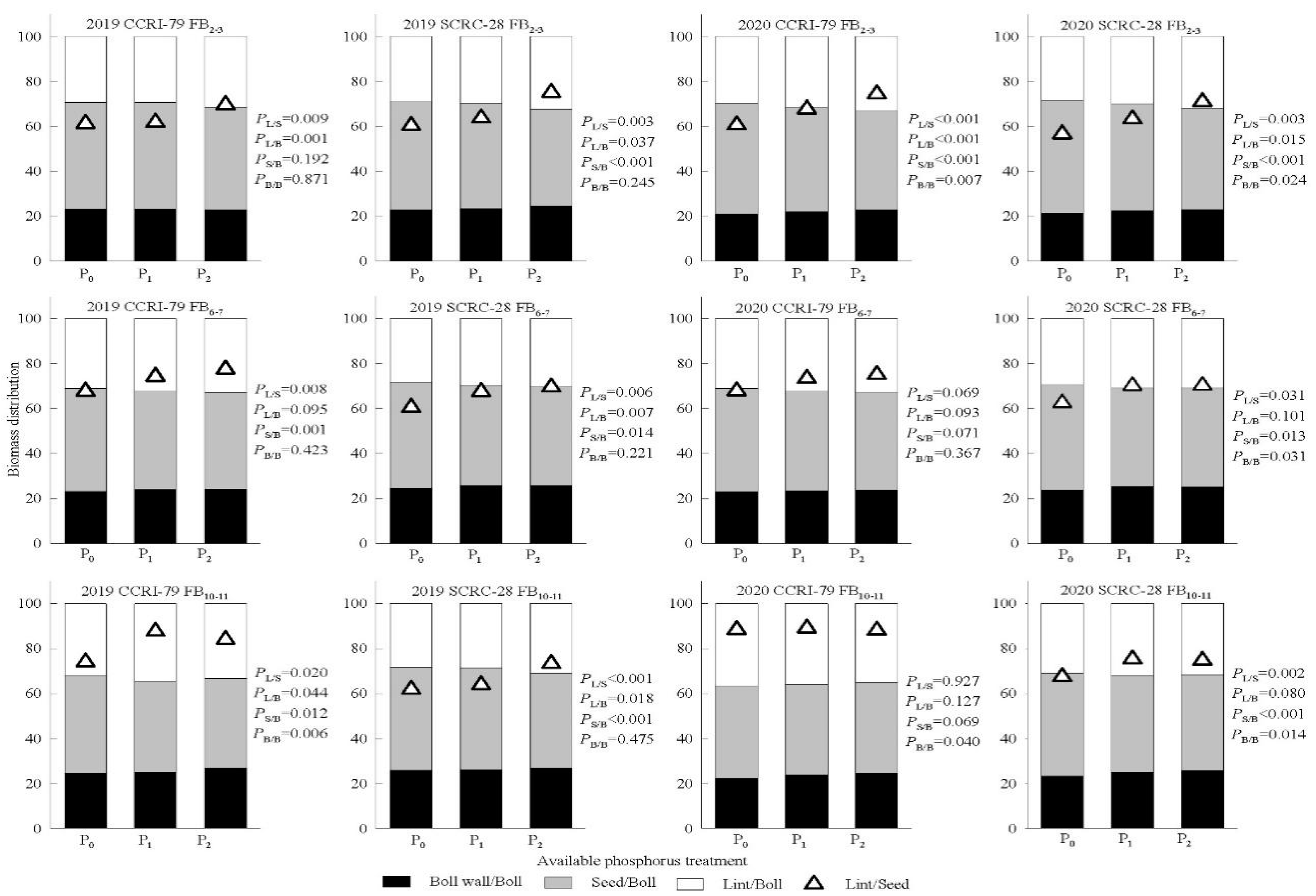
3.7. Cotton Boll Biomass Accumulation and Distribution on Different Fruiting Branches
3.8. Correlation of Source Indices to Lint Yield and Yield Components in the Subtending Leaves
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Saleem, M.F.; Shakeel, A.; Bilal, M.F.; Shahid, M.Q.; Anjum, S.A. Effect of different phosphorus levels on earliness and yield of cotton cultivars. Soil Environ. 2010, 29, 128–135. [Google Scholar]
- Mao, S.C. China Cotton Cultivation, 4th ed.; Shanghai Scientific and Technical Press: Shanghai, China, 2019; pp. 1–319. (In Chinese) [Google Scholar]
- National Bureau of Statistics. Available online: http://www.stats.gov.cn/xxgk/sjfb/zxfb2020/202112/t20211214_1825249.html (accessed on 14 December 2021).
- Dong, H.L. Research progress on fertilization technology of cotton. Cott. Sci. 2007, 19, 378–384, (In Chinese with English Abstract). [Google Scholar]
- Sarkar, R.K.; Majumdar, R.C. Effect of farmyard manure, nitrogen and phosphorus on yield and fibre quality of upland cotton (Gossypium hirsutum) in rice fallow land. Indian J. Agric. Sci. 2002, 72, 607–610. [Google Scholar]
- Saleem, M.F.; Cheema, M.A.; Bilal, M.F.; Anjum, S.A.; Khurshid, I. Fiber quality of cotton (Gossypium hirsutum) cultivars under different phosphorus levels. J. Anim. Plant Sci. 2011, 21, 26–30. [Google Scholar] [CrossRef] [Green Version]
- Bronson, K.F.; Keeling, J.W.; Booker, J.D.; Chua, T.T.; Lascano, R.J. Influence of landscape position, soil series, and phosphorus fertilizer on cotton lint yield. Agron. J. 2003, 95, 949–957. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.L.; Wang, Q.H.; Ye, Z.P.; Stiles, S.; Feng, G. Optimisation of phosphorus fertilisation promotes biomass and phosphorus nutrient accumulation, partitioning and translocation in three cotton (Gossypium hirsutum) genotypes. Crop Pasture Sci. 2020, 71, 56–69. [Google Scholar] [CrossRef]
- Aslam, M.; Memon, M.; Khan, P.; Imtiaz, M. Performance of fertigation technique for phosphorus application in cotton. Soil Environ. 2009, 28, 45–48. [Google Scholar]
- Singh, V.; Pallaghy, C.K.; Singh, D. Phosphorus nutrition and tolerance of cotton to water stress: II. Water relations, free and bound water and leaf expansion rate. Field Crop. Res. 2006, 96, 199–206. [Google Scholar] [CrossRef]
- Sawan, Z.M. Effect of nitrogen, phosphorus fertilization and growth regulators on cotton yield and fiber properties. J. Agron. Crop Sci. 1986, 156, 237–245. [Google Scholar] [CrossRef]
- Janaki, P.; Rajan, R.V.A. Phosphorus utilization efficiency in cotton using P-32 Tracer: Effect of different levels of nitrogen and phosphorus. Adv. Plant Sci. 2008, 21, 559–564. [Google Scholar]
- Katkar, R.N.; Turkhede, A.B.; Solanke, V.M.; Wankhade, S.T.; Sakhare, B.A. Effect of foliar sprays of nutrients and chemicals on yield and quality of cotton under rainfed condition. Res. Crop. 2002, 3, 27–29. [Google Scholar]
- Hearn, A.B. Cotton nutrition. Field Crop Abstr. 1981, 34, 11–33. [Google Scholar]
- Gifford, R.M.; Lutze, J.L.; Barrett, D. Global atmospheric change effects on terrestrial carbon sequestration: Exploration with a global C- and N-cycle model (CQUESTN). Plant Soil 1995, 187, 369–387. [Google Scholar] [CrossRef]
- Wang, X.J.; Guppy, C.N.; Watson, L.; Sale, P.W.G.; Tang, C. Availability of sparingly soluble phosphorus sources to cotton (Gossypium hirsutumL.), wheat (Triticum aestivumL.) and white lupin (Lupinus albusL.) with different forms of nitrogen as evaluated by a32P isotopic dilution technique. Plant Soil 2011, 348, 85–98. [Google Scholar] [CrossRef]
- Ahmad, F.; Uddin, S.; Ahmad, N.; Islam, R. Phosphorus–microbes interaction on growth, yield and phosphorus-use efficiency of irrigated cotton. Arch. Agron. Soil Sci. 2013, 59, 341–351. [Google Scholar] [CrossRef]
- Lu, R.K. The phosphorus level of soil and environmental protection of water body. Phos. Comp. Fert. 2003, 1, 4–8, (In Chinese with English Abstract). [Google Scholar]
- Xu, W.F.; Zhang, Q.; Yuan, W.; Xu, F.Y.; Aslam, M.M.; Miao, R.; Li, Y.; Wang, Q.W.; Li, X.; Zhang, X.; et al. The genome evolution and low-phosphorus adaptation in white lupin. Nat.Commun. 2020, 11, 1069. [Google Scholar] [CrossRef] [Green Version]
- Cordell, D.; Drangert, J.O.; White, S. The story of phosphorus: Global food security and food for thought. Glob. Environ. Chang. 2009, 19, 292–305. [Google Scholar] [CrossRef]
- Gao, Y.; Huang, H.Y.; Zhao, H.Y.; Xia, H.Q.; Sun, M.; Li, Z.Y.; Li, P.C.; Zheng, C.S.; Dong, H.L.; Liu, J.R. Phosphorus affects enzymatic activity and chemical properties of cotton soil. Plant Soil Environ. 2019, 65, 361–368. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.E.; Jacobsen, J.S. Soil Inorganic Phosphorus fractions and their uptake relationships in calcareous soils. Soil Sci. Soc. Am. J. 1990, 54, 1666–1669. [Google Scholar] [CrossRef]
- Cai, J.; Chen, L.; Qu, H.Y.; Lian, J.; Liu, W.; Hu, Y.B.; Xu, G.H. Alteration of nutrient allocation and transporter genes expression in rice under N, P, K, and Mg deficiencies. Acta Physiol. Plant. 2012, 34, 939–946. [Google Scholar] [CrossRef]
- Ismail, C.; Christine, H.; Horst, M. Changes in phloem export of sucrose in leaves in response to phosphorus, potassium and magnesium deficiency in bean plants. J. Exp. Bot. 1994, 45, 1251–1257. [Google Scholar] [CrossRef]
- Groot, C.; Marcelis, L.F.M.; Boogaard, R.; Kaiser, W.M.; Lambers, H. Interaction of nitrogen and phosphorus nutrition in determining growth. Plant Soil 2003, 248, 257–268. [Google Scholar] [CrossRef]
- Liu, J.R.; Peng, J.; Xia, H.Q.; Li, P.C.; Li, Z.Y.; Sun, M.; Zheng, C.S.; Dong, H.L. High soil available phosphorus favors carbon metabolism in cotton leaves in pot trials. J. Plant Growth Regul. 2021, 40, 974–985. [Google Scholar] [CrossRef]
- Hermans, C.; Hammond, J.P.; White, P.J.; Verbruggen, N. How do plants respond to nutrient shortage by biomass allocation? Trends Plant Sci. 2006, 11, 610–617. [Google Scholar] [CrossRef]
- Zheng, J.F.; Bai, Z.Y.; Li, C.D.; Mi, S.Y.; Liu, Y.X. The effect of phosphorus deficiency stress on soluble sugar content and soluble protein content and chromosome of wheat substitution lines. Acta Agric.Boreali-Sin. 2013, 28, 27–31, (In Chinese with English Abstract). [Google Scholar]
- Rao, I.M.; Arulanantham, A.R.; Terry, N. Leaf phosphate status, photosynthesis and carbon partitioning in sugar beet: II. diurnal changes in sugar phosphates, adenylates, and nicotinamide nucleotides. Plant Physiol. 1989, 90, 820–826. [Google Scholar] [CrossRef] [Green Version]
- Crafts-Brandner, S.J. Phosphorus nutrition influence on leaf senescence in soybean. Plant Physiol. 1992, 98, 1128–1132. [Google Scholar] [CrossRef] [Green Version]
- Crafts-Brandner, S.J.; Salvucci, M.E.; Sims, J.L.; Sutton, T.G. Phosphorus nutrition influence on plant growth and nonstructural carbohydrate accumulation in tobacco. Crop Sci. 1990, 30, 609–614. [Google Scholar] [CrossRef]
- Loewe, A.; Einig, W.; Shi, L.; Dizengremel, P.; Hampp, R. Mycorrhiza formation and elevated CO2 both increase the capacity for sucrose synthesis in source leaves of spruce and aspen. New Phytol. 2000, 145, 565–574. [Google Scholar] [CrossRef]
- Ciereszko, I.; Zambrzycka, A.; Rychter, A. Sucrose hydrolysis in bean roots (Phaseolus vulgaris L.) under phosphate deficiency. Plant Sci. 1998, 133, 139–144. [Google Scholar] [CrossRef]
- Yang, C.Q.; Liu, R.X.; Zhang, G.W.; Xu, L.H.; Zhou, Z.G. Effects of waterlogging on sucrose metabolism of the subtending leaf of cotton boll and boll weight during flowering and boll-forming stage. Acta Agrono Sin. 2014, 40, 908–914, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Daie, J. Cytosolic fructose-1,6-bisphosphatase: A key enzyme in the sucrose biosynthetic pathway. Photosynth. Res. 1993, 38, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.A.; Hajirezaei, M.; Peisker, M.; Tschiersch, H.; Sonnewald, U.; Bornke, F. Decreased sucrose-6-phosphate phosphatase level in transgenic tobacco inhibits photosynthesis, alters carbohydrate partitioning, and reduces growth. Planta 2005, 221, 479–492. [Google Scholar] [CrossRef]
- Grof, C.P.L.; Knight, D.P.; McNeil, S.D.; Lunn, J.E.; Campbell, J.A. A modified assay method shows leaf sucrose-phosphate synthase activity is correlated with leaf sucrose content across a range of sugarcane varieties. Aus. J. Plant Physiol. 1998, 25, 499–502. [Google Scholar] [CrossRef]
- Xu, X.Y.; Yang, Y.S.; Liu, C.X.; Sun, Y.M.; Zhang, T.; Hou, M.L.; Huang, S.Z.; Yuan, H.Y. The evolutionary history of the sucrose synthase gene family in higher plants. BMC Plant Biol. 2019, 19, 566. [Google Scholar] [CrossRef] [Green Version]
- Geigenberger, P.; Stitt, M. Sucrose synthase catalyses a readily reversible reaction in vivo in developing potato tubers and other plant tissues. Planta 1993, 189, 329–339. [Google Scholar] [CrossRef]
- Liu, J.; Wang, Y.; Chen, J.; Lv, F.; Ma, Y.; Meng, Y.; Chen, B.; Zhou, Z. Sucrose metabolism in the subtending leaf to cotton boll at different fruiting branch nodes and the relationship to boll weight. J. Agric. Sci. 2014, 152, 790–804. [Google Scholar] [CrossRef]
- Garcia-Caparros, P.; Lao, M.T.; Preciado-Rangel, P.; Sanchez, E. Phosphorus and carbohydrate metabolism in green bean plants subjected to increasing phosphorus concentration in the nutrient solution. Agronomy 2021, 11, 245. [Google Scholar] [CrossRef]
- Brooks, A. Effects of phosphorus nutrition on ribulose-1,5-bisphosphate carboxylase activation, photosynthetic quantum yield and amounts of some Calvin-cycle metabolites in spinach leaves. Aus. J. Plant Physiol. 1986, 13, 221–237. [Google Scholar] [CrossRef]
- Usuda, H.; Shimogawara, K. Phosphate deficiency in maize. II. Enzyme activities. Plant Cell Physiol. 1991, 32, 1313–1317. [Google Scholar]
- Rao, I.M.; Terry, N. Leaf phosphate status, photosynthesis, and carbon partitioning in sugar beet: I. Changes in growth, gas exchange, and calvin cycle enzymes. Plant Physiol. 1989, 90, 814–819. [Google Scholar] [CrossRef] [Green Version]
- Rao, I.M.; Fredeen, A.L.; Terry, N. Leaf Phosphate Status, Photosynthesis, and Carbon Partitioning in Sugar Beet: III. Diurnal Changes in Carbon Partitioning and Carbon Export. Plant Physiol. 1990, 92, 29–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, M.J.; Stitt, M. Effects of nitrogen and phosphorus deficiencies on levels of carbohydrates, respiratory enzymes and metabolites in seedlings of tobacco and their response to exogenous sucrose. Plant Cell Physiol. 1993, 16, 1047–1057. [Google Scholar] [CrossRef]
- Wang, G. The Effects and Mechanism on the Cotton Growth under the Available Phosphorus Levels. Master’s Thesis, Chinese Academy of Agricultural Sciences, Anyang, China, 2016. [Google Scholar]
- Ahmad, I.; Zhou, G.; Zhu, G.; Ahmad, Z.; Ibrahim, M. Response of leaf characteristics of BT cotton plants to ratio of nitrogen, phosphorus and potassium. Pak. J. Bot. 2021, 53, 3. [Google Scholar] [CrossRef]
- Wells, R. Response of leaf ontogeny and photosynthetic activity to reproductive growth in cotton. Plant Physiol. 1988, 87, 274–279. [Google Scholar] [CrossRef] [Green Version]
- Bondada, B.R.; Oosterhuis, D.M. Canopy photosynthesis, specific leaf weight, and yield components of cotton under varying nitrogen supply. J.Plant Nutr. 2001, 24, 469–477. [Google Scholar] [CrossRef]
- Wang, R.; Ji, S.; Zhang, P.; Meng, Y.L.; Zhou, Z.G. Drought effects on cotton yield and fiber quality on different fruiting branches. Crop Sci. 2016, 56, 1265–1276. [Google Scholar] [CrossRef]
- Li, X.G. Effect of Controlled Release N Fertilizer on Spatio-Temporal Distribution of Cotton Bolls and Fiber Quality and its Physiological Basis. Master’s Thesis, Shandong Agricultural University, Tai’an, China, 2009. [Google Scholar]
- Li, Z.T.; Chen, E.Y.; Song, X.L.; Zhang, M.L.; Zhao, Q.L.; Xu, X.L.; Ji, H.; Sun, X.Z. Effects of the rate and the timing of potassium application on cotton (Gossypium hirsutum L.) yield and on fibre quality of cotton bolls at different fruit positions. Plant Nutr. Fert. Sci. 2012, 18, 123–131, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Li, P.C.; Dong, H.L.; Zheng, C.S.; Sun, M.; Liu, A.Z.; Wang, G.P.; Liu, S.D.; Zhang, S.P.; Chen, J.; Li, Y.B.; et al. Optimizing nitrogen application rate and plant density for improving cotton yield and nitrogen use efficiency in the North China Plain. PLoS ONE 2017, 12, 15. [Google Scholar] [CrossRef] [Green Version]
- Kuai, J.; Zhou, Z.G.; Wang, Y.H.; Meng, Y.L.; Chen, B.L.; Zhao, W.Q. The effects of short-term waterlogging on the lint yield and yield components of cotton with respect to boll position. Eur. J. Agron. 2015, 67, 61–74. [Google Scholar] [CrossRef]
- Hendrix, D.L. Rapid extraction and analysis of nonstructural carbohydrates in plant tissues. Crop Sci. 1993, 33, 1306–1311. [Google Scholar] [CrossRef]
- Seifter, S.; Dayton, S. The estimation of glycogen with the anthrone reagent. Arch. Biochem. 1950, 25, 191–200. [Google Scholar] [PubMed]
- Liu, J.R.; Ma, Y.N.; Lv, F.J.; Chen, J.; Zhou, Z.G.; Wang, Y.H.; Abudurezike, A.; Oosterhuis, D.M. Changes of sucrose metabolism in leaf subtending to cotton boll under cool temperature due to late planting. Field Crop. Res. 2013, 144, 200–211. [Google Scholar] [CrossRef]
- Jang, H.K.; Lee, S.W.; Lee, Y.H.; Hahn, T.R. Purification and characterization of a recombinant pea cytoplasmic fructose-1,6-bisphosphatase. Protein Expr. Purif. 2003, 28, 42–48. [Google Scholar] [CrossRef]
- Huber, S.C.; Israel, D.W. Biochemical basis for partitioning of photosynthetically fixed carbon between starch and sucrose in soybean (Glycine maxMerr.) leaves. Plant Physiol. 1982, 69, 691–696. [Google Scholar] [CrossRef] [Green Version]
- Singh, V.; Pallaghy, C.K.; Singh, D. Phosphorus nutrition and tolerance of cotton to water stress I. Seed cotton yield and leaf morphology. Field Crop. Res. 2006, 96, 191–198. [Google Scholar] [CrossRef]
- Ackerson, R.C. Osmoregulation in cotton in response to water stress: III. Effects of phosphorus fertility. Plant Physiol. 1985, 77, 309–312. [Google Scholar] [CrossRef] [Green Version]
- Ashley, D.A. 14C-labelled photosynthate translocation and utilization in cotton plants. Crop Sci. 1972, 12, 69–74. [Google Scholar] [CrossRef]
- Constable, G.A.; Rawson, H.M. Carbon production and utilization in cotton: Inferences from a carbon budget. Aus. J. Plant Physiol. 1980, 7, 539–553. [Google Scholar] [CrossRef]
- Wullschleger, S.D.; Oosterhuis, D.M. Photosynthetic carbon production and use by developing cotton leaves and bolls. Crop Sci. 1990, 30, 1259–1264. [Google Scholar] [CrossRef]
- Hu, W.; Dai, Y.; Zhao, W.; Meng, Y.; Chen, B.; Wang, Y.; Zhou, Z. Effects of long-term elevation of air temperature on sucrose metabolism in cotton leaves at different positions. J. Agron. Crop Sci. 2017, 203, 539–552. [Google Scholar] [CrossRef]
- Huber, S.C. Biochemical Mechanism for regulation of sucrose accumulation in leaves during photosynthesis. Plant Physiol. 1989, 91, 656–662. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.C.; Wang, J.; Zhu, M.Y.; Jiang, J. Molecular mechanism of carbohydrate metabolism participation in tomato response to low phosphorus stress. Mol. Plant Breed. 2015, 13, 2833–2842, (In Chinese with English Abstract). [Google Scholar]
- Wasaki, J.; Shinano, T.; Onishi, K.; Yonetani, R.; Yazaki, J.; Fujii, F.; Shimbo, K.; Ishikawa, M.; Shimatani, Z.; Nagata, Y.; et al. Transcriptomic analysis indicates putative metabolic changes caused by manipulation of phosphorus availability in rice leaves. J. Exp. Bot. 2006, 57, 2049–2059. [Google Scholar] [CrossRef] [Green Version]
- Jiang, D.; Tao, Q.N.; Cao, W.X. Effects of waterlogging on accumulation and redistribution of water solublecarbohydrate metabolism in culm of wheat. Acta Agron. Sin. 2002, 28, 230–234, (In Chinese with English Abstract). [Google Scholar]
- Qiu, J.; Israel, D.W. Diurnal starch accumulation and utilization in phosphorus-deficient soybean plants. Plant Physiol. 1992, 98, 316–323. [Google Scholar] [CrossRef] [Green Version]
- Poudyal, S.; Owen, J.S., Jr.; Sharkey, T.D.; Fernandez, R.T.; Cregg, B. Phosphorus requirement for biomass accumulation is higher compared to photosynthetic biochemistry for three ornamental shrubs. Sci. Hortic. 2021, 275, 109719. [Google Scholar] [CrossRef]
- Natr, L. Mineral nutrients—A ubiquitous stress factor for photosynthesis. Photosynthetica 1992, 27, 271–294. [Google Scholar]
- Hendrix, D.L.; Grange, R.I. Carbon partitioning and export from mature cotton leaves. Plant Physiol. 1991, 95, 228–233. [Google Scholar] [CrossRef] [Green Version]
- Lv, F.J.; Liu, J.R.; Ma, Y.N.; Chen, J.; Abudurezikekey, A.K.; Wang, Y.H.; Chen, B.L.; Meng, Y.L.; Zhou, Z.G. Effect of shading on cotton yield and quality on different fruiting branches. Crop Sci. 2013, 53, 2670–2678. [Google Scholar] [CrossRef]
- Usuda, H.; Shimogawara, K. Phosphate deficiency in maize. IV. Changes in amounts of sucrose phosphate synthase during the course of phosphate deprivation. Plant Cell Physiol. 1993, 34, 767–770. [Google Scholar] [CrossRef]
- Wu, S.; Li, M.; Zhang, C.; Tan, Q.; Yang, X.; Sun, X.; Pan, Z.; Deng, X.; Hu, C. Effects of phosphorus on fruit soluble sugar and citric acid accumulations in citrus. Plant Physiol. Biochem. 2021, 160, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Lunn, J.E.; Hatch, M.D. Primary partitioning and storage of photosynthate in sucrose andstarch in leaves of C4 plants. Planta 1995, 197, 385–391. [Google Scholar] [CrossRef]
- Schachtman, D.P.; Reid, R.J.; Ayling, S.M. Phosphorus uptake by plants: From soil to cell. Plant Physiol. 1998, 116, 447–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, S.; Nakamoto, H.; Ku, M.S.; Edwards, G.E. Influence of leaf age on photosynthesis, enzyme activity, and metabolite levels in wheat. Plant Physiol. 1987, 84, 1244–1248. [Google Scholar] [CrossRef] [Green Version]
- Li, H.J.; Wang, J.W.; Ali, S.; Iqbal, B.; Zhang, H.; Wang, S.S.; Chen, B.L.; Zhou, Z.G. Agronomic traits at the seedling stage, yield, and fiber quality in two cotton (Gossypium hirsutum L.) cultivars in response to phosphorus deficiency. Soil Sci. Plant Nutr. 2020, 66, 308–316. [Google Scholar] [CrossRef]
- Iqbal, B.; Kong, F.; Ullah, I.; Ali, S.; Li, H.; Wang, J.; Khattak, W.A.; Zhou, Z. Phosphorus application improves the cotton yield by enhancing reproductive organ biomass and nutrient accumulation in two cotton cultivars with different phosphorus sensitivity. Agronomy 2020, 10, 153. [Google Scholar] [CrossRef] [Green Version]
- Sun, M.; Li, P.C.; Zheng, C.S.; Liu, S.; Liu, A.Z.; Han, H.M.; Liu, J.R.; Dong, H.L. Effects of low phosphorus stress on root morphology and physiological characteristics of different cotton genotypes at the seedling stage. Cott. Sci. 2018, 30, 45–52, (In Chinese with English Abstract). [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | OM 1 | TN | AN | AK | pH |
|---|---|---|---|---|---|
| (g kg−1) | (mg kg−1) | ||||
| 2019 | 12.9 | 0.86 | 64.4 | 163.6 | 7.7 |
| 2020 | 13.1 | 0.85 | 63.3 | 180.4 | 7.6 |
| Year | FBP 1 | AP Treatment | Maximum Sucrose Content (mg g−1 DW) | Minimum Sucrose Content (mg g−1 DW) | Sucrose Transformation Rate (%) | |||
|---|---|---|---|---|---|---|---|---|
| CCRI-79 | SCRC-28 | CCRI-79 | SCRC-28 | CCRI-79 | SCRC-28 | |||
| 2019 | FB2–3 | P0 3 | 21.7 a 2 | 21.7 a | 11.1 a | 12.4 a | 48.9 b | 42.9 c |
| P1 | 17.6 b | 20.5 b | 8.9 b | 10.9 b | 49.5 b | 47.0 b | ||
| P2 | 15.2 c | 18.6 c | 7.1 c | 9.3 c | 53.3 a | 49.8 a | ||
| CV (%) 4 | 18.2 | 7.6 | 22.2 | 14.0 | 4.7 | 7.4 | ||
| FB6–7 | P0 | 22.9 a | 24.8 a | 11.4 a | 13.8 a | 50.1 b | 44.2 c | |
| P1 | 21.1 b | 24.4 a | 9.7 b | 13.0 b | 53.8 a | 46.7 b | ||
| P2 | 17.7 c | 22.1 b | 8.0 c | 11.1 c | 54.8 a | 50.1 a | ||
| CV (%) | 12.8 | 6.0 | 17.6 | 11.3 | 4.7 | 6.3 | ||
| FB10–11 | P0 | 22.2 a | 22.7 a | 10.9 a | 13.2 a | 50.9 b | 42.1 b | |
| P1 | 18.9 b | 21.7 b | 9.0 b | 11.8 b | 52.6 ab | 45.7 a | ||
| P2 | 18.7 b | 19.9 c | 8.6 b | 10.5 c | 54.1 a | 47.1 a | ||
| CV (%) | 9.7 | 6.6 | 13.0 | 11.0 | 3.0 | 5.7 | ||
| 2020 | FB2–3 | P0 | 26.3 a | 30.1 a | 14.1 a | 17.3 a | 46.3 c | 42.3 b |
| P1 | 25.0 b | 26.9 b | 12.1 b | 14.1 b | 51.7 b | 47.5 a | ||
| P2 | 22.9 c | 24.6 c | 10.6 c | 11.8 c | 53.9 a | 52.0 a | ||
| CV (%) | 6.9 | 10.3 | 14.6 | 19.4 | 7.7 | 10.3 | ||
| FB6–7 | P0 | 27.0 a | 29.8 a | 13.0 a | 16.3 a | 51.9 b | 45.5 b | |
| P1 | 25.3 b | 26.4 b | 12.0 b | 13.8 b | 52.6 b | 47.7 b | ||
| P2 | 24.6 b | 24.4 c | 11.0 c | 11.6 c | 55.3 a | 52.7 a | ||
| CV (%) | 4.8 | 10.2 | 8.2 | 17.0 | 3.4 | 7.6 | ||
| FB10–11 | P0 | 24.7 a | 27.6 a | 11.3 a | 15.4 a | 52.0 b | 44.1 b | |
| P1 | 23.7 b | 25.1 b | 10.2 b | 13.6 b | 54.1 ab | 45.7 b | ||
| P2 | 22.2 c | 22.6 c | 9.3 c | 11.6 c | 58.0 a | 48.9 a | ||
| CV (%) | 5.4 | 9.9 | 9.7 | 14.3 | 5.5 | 5.2 | ||
| Cultivar | Comparison with | Days Post-Anthesis (d) | ||||||
|---|---|---|---|---|---|---|---|---|
| 10 | 17 | 24 | 31 | 38 | 45 | Mean | ||
| CCRI-79 | Soluble sugar | 0.071 | −0.538 * 1 | −0.002 | −0.050 | 0.077 | −0.131 | −0.096 |
| Sucrose | −0.002 | −0.314 | −0.269 | −0.016 | 0.083 | 0.066 | −0.075 | |
| Starch | −0.312 | −0.444 | −0.459 | −0.146 | −0.376 | −0.241 | −0.330 | |
| SCRC-28 | Soluble sugar | −0.251 | −0.597 ** | −0.406 | −0.357 | −0.201 | −0.667 ** | −0.413 |
| Sucrose | −0.218 | −0.565 * | −0.715 ** | −0.220 | −0.451 | −0.447 | −0.436 | |
| Starch | −0.887 ** | −0.872 ** | −0.863 ** | −0.473 * | −0.773 ** | −0.796 ** | −0.777 | |
| Cultivar | Comparison with | Days Post-Anthesis (d) | ||||||
|---|---|---|---|---|---|---|---|---|
| 10 | 17 | 24 | 31 | 38 | 45 | Mean | ||
| CCRI-79 | Initial Rubisco 1 | −0.014 | 0.046 | 0.161 | 0.199 | 0.021 | 0.237 | 0.108 |
| cy-FBPase 2 | −0.166 | −0.180 | −0.212 | −0.280 | −0.218 | −0.423 | −0.247 | |
| SPS 3 | 0.046 | 0.760 ** 5 | 0.772 ** | 0.258 | 0.495 * | 0.452 | 0.464 | |
| SuSy 4 | 0.616 ** | 0.699 ** | 0.678 ** | −0.089 | −0.343 | 0.337 | 0.316 | |
| SCRC-28 | Initial Rubisco | −0.053 | 0.173 | 0.039 | −0.042 | −0.101 | −0.110 | −0.016 |
| cy-FBPase | −0.771 ** | −0.573 * | −0.783 ** | −0.811 ** | −0.884 ** | −0.880 | −0.784 | |
| SPS | 0.720 ** | 0.634 ** | 0.861 ** | −0.068 | 0.208 | 0.738 ** | 0.516 | |
| SuSy | 0.770 ** | 0.369 | 0.553 * | −0.115 | 0.240 | 0.641 ** | 1.410 | |
| Cultivar | FBP 1 | AP Treatment | 2019 | 2020 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Boll Number (Boll Plant−1) | Boll Biomass (g Boll−1) | Lint Percentage (%) | Lint Yield (kg ha−1) | Boll Number (Boll Plant−1) | Boll Biomass (g Boll−1) | Lint Percentage (%) | Lint Yield (kg ha−1) | |||
| CCRI-79 | FB1–4 | P0 2 | 4.3 b 3 | 5.3 c | 38.0 b | 349.7 c | 4.0 b | 5.4 c | 37.7 c | 342.3 c |
| P1 | 5.5 ab | 6.2 b | 38.2 b | 527.1 b | 6.9 a | 6.0 b | 40.4 b | 681.0 b | ||
| P2 | 6.3 a | 6.9 a | 41.0 a | 727.4 a | 7.4 a | 7.0 a | 42.7 a | 885.8 a | ||
| CV (%) 4 | 18.8 | 13.3 | 4.4 | 35.3 | 30.1 | 12.8 | 6.1 | 43.1 | ||
| FB5–8 | P0 | 3.1 b | 5.9 c | 40.3 b | 295.1 b | 3.9 b | 6.1 b | 40.4 a | 384.9 c | |
| P1 | 3.4 ab | 6.4 b | 42.6 a | 367.2 b | 4.5 ab | 6.5 b | 42.3 a | 496.1 b | ||
| P2 | 4.2 a | 7.1 a | 43.7 a | 524.1 a | 4.8 a | 7.1 a | 42.9 a | 592.2 a | ||
| CV (%) | 16.6 | 9.7 | 4.0 | 29.6 | 10.4 | 8.1 | 3.1 | 21.1 | ||
| FB9–12 | P0 | 3.4 a | 6.2 b | 42.5 b | 332.5 b | 3.0 a | 6.5 b | 46.9 a | 367.0 b | |
| P1 | 3.2 a | 6.4 b | 46.7 a | 396.9 ab | 3.5 a | 7.1 a | 47.1 a | 464.9 a | ||
| P2 | 3.7 a | 7.3 a | 45.6 a | 469.1 a | 3.6 a | 7.7 a | 46.8 a | 510.3 a | ||
| CV (%) | 7.3 | 8.7 | 4.9 | 17.1 | 9.5 | 8.5 | 0.3 | 16.4 | ||
| SCRC-28 | FB1–4 | P0 | 3.2 b | 5.0 c | 37.6 c | 246.0 c | 3.5 c | 4.7 c | 36.1 b | 248.1 c |
| P1 | 4.8 a | 6.3 b | 38.9 b | 473.1 b | 6.5 b | 6.2 b | 38.7 ab | 641.6 b | ||
| P2 | 5.6 a | 6.7 a | 42.9 a | 633.6 a | 7.1 a | 7.2 a | 41.5 a | 868.5 a | ||
| CV (%) | 27.0 | 14.2 | 6.9 | 43.2 | 33.8 | 20.3 | 6.9 | 53.6 | ||
| FB5–8 | P0 | 2.2 b | 5.4 b | 37.7 b | 176.8 c | 3.2 b | 5.5 c | 38.4 b | 268.8 b | |
| P1 | 2.9 b | 6.3 a | 40.3 a | 285.9 b | 3.7 ab | 6.5 b | 41.2 a | 388.5 b | ||
| P2 | 3.8 a | 6.7 a | 41.0 a | 408.7 a | 4.6 a | 7.5 a | 41.3 a | 579.6 a | ||
| CV (%) | 27.0 | 11.3 | 4.4 | 39.9 | 18.5 | 15.3 | 4.1 | 38.0 | ||
| FB9–12 | P0 | 2.5 b | 5.3 b | 38.2 b | 196.2 b | 1.9 b | 5.5 c | 40.3 b | 172.6 b | |
| P1 | 3.0 ab | 6.1 a | 39.0 b | 277.5 a | 2.4 a | 5.9 b | 42.9 a | 241.9 a | ||
| P2 | 3.3 a | 6.5 a | 42.3 a | 345.3 a | 2.4 a | 6.5 a | 42.7 a | 262.1 a | ||
| CV (%) | 13.8 | 10.3 | 5.5 | 27.3 | 12.9 | 8.9 | 3.6 | 20.8 | ||
| Significance | ||||||||||
| Cultivar (C) | NS 5 | ** | ** | ** | ** | ** | ** | ** | ||
| FBP | ** | ** | ** | ** | ** | ** | ** | ** | ||
| AP | ** | ** | ** | ** | ** | ** | ** | ** | ||
| C × FBP | ** | ** | ** | NS | NS | ** | ** | ** | ||
| C × AP | * | ** | * | NS | NS | ** | NS | NS | ||
| FBP × AP | NS | * | * | ** | ** | ** | ** | ** | ||
| C × FBP × AP | NS | NS | ** | NS | NS | * | NS | NS | ||
| Cultivar | FBP 1 | AP Treatment | 2019 | 2020 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Boll Dry Mass (g) | Proportion (%) | Boll Dry Mass (g) | Proportion (%) | |||||||
| Boll Wall | Lint | Seed | L/S 2 | Boll Wall | Lint | Seed | L/S | |||
| CCRI-79 | FB1–4 | P0 | 1.2 b 3 | 1.5 c | 2.5 b | 61.2 b | 1.1 c | 1.6 c | 2.7 c | 60.6 c |
| P1 | 1.4 ab | 1.8 b | 3.0 a | 61.8 b | 1.3 b | 1.9 b | 2.8 b | 67.7 b | ||
| P2 | 1.6 a | 2.2 a | 3.2 a | 69.6 a | 1.6 a | 2.3 a | 3.1 a | 74.4 a | ||
| CV (%) 4 | 12.4 | 17.6 | 11.2 | 7.3 | 16.7 | 17.0 | 7.7 | 10.2 | ||
| FB5–8 | P0 | 1.4 c | 1.8 c | 2.7 b | 67.6 b | 1.4 b | 1.9 c | 2.8 a | 67.8 a | |
| P1 | 1.5 b | 2.1 b | 2.8 b | 74.2 a | 1.5 b | 2.1 b | 2.9 a | 73.3 a | ||
| P2 | 1.7 a | 2.3 a | 3.0 a | 77.5 a | 1.7 a | 2.4 a | 3.1 a | 75.1 a | ||
| CV (%) | 11.7 | 13.0 | 6.4 | 6.9 | 9.8 | 11.1 | 4.8 | 5.2 | ||
| FB9–12 | P0 | 1.5 b | 2.0 c | 2.7 ab | 74.0 b | 1.5 b | 2.3 b | 2.7 b | 88.3 a | |
| P1 | 1.6 b | 2.2 b | 2.5 b | 87.5 a | 1.7 a | 2.5 a | 2.9 ab | 89.0 a | ||
| P2 | 2.0 a | 2.4 a | 2.9 a | 84.0 a | 1.9 a | 2.7 a | 3.1 a | 88.0 a | ||
| CV (%) | 13.9 | 9.9 | 6.2 | 8.6 | 12.9 | 7.3 | 7.9 | 0.5 | ||
| SCRC-28 | FB1–4 | P0 | 1.1 c | 1.5 c | 2.4 b | 60.3 b | 1.0 c | 1.4 c | 2.4 b | 56.6 b |
| P1 | 1.5 b | 1.9 b | 3.0 a | 63.6 b | 1.4 b | 1.9 b | 3.0 a | 63.3 ab | ||
| P2 | 1.6 a | 2.2 a | 2.9 a | 75.1 a | 1.7 a | 2.3 a | 3.3 a | 71.0 a | ||
| CV (%) | 17.6 | 19 | 10.2 | 11.7 | 24.4 | 26.5 | 15.5 | 11.3 | ||
| FB5–8 | P0 | 1.3 b | 1.5 b | 2.5 b | 60.5 b | 1.3 c | 1.6 c | 2.6 c | 62.5 b | |
| P1 | 1.6 a | 1.9 a | 2.8 ab | 67.5 a | 1.6 b | 2.0 b | 2.9 b | 70.2 a | ||
| P2 | 1.7 a | 2.0 a | 3.0 a | 69.5 a | 1.9 a | 2.4 a | 3.3 a | 70.3 a | ||
| CV (%) | 14 | 14.5 | 7.7 | 7.2 | 17.8 | 20.0 | 11.7 | 6.6 | ||
| FB9–12 | P0 | 1.4 b | 1.5 c | 2.4 b | 61.8 b | 1.3 cc | 1.7 c | 2.5 b | 67.4 b | |
| P1 | 1.6 ab | 1.8 b | 2.8 a | 63.8 b | 1.5 b | 1.9 b | 2.6 b | 74.7 a | ||
| P2 | 1.7 a | 2.0 a | 2.7 ab | 73.3 a | 1.7 a | 2.1 a | 2.8 a | 75.2 a | ||
| CV (%) | 12.3 | 14.2 | 7.2 | 9.2 | 13.6 | 9.2 | 5.3 | 6.0 | ||
| Significance | ||||||||||
| Cultivar (C) | NS 5 | ** | * | ** | * | ** | ** | ** | ||
| FBP | ** | ** | ** | ** | ** | ** | ** | ** | ||
| AP | ** | ** | ** | ** | ** | ** | ** | ** | ||
| C × FBP | * | ** | NS | ** | ** | ** | ** | ** | ||
| C × AP | NS | NS | ** | * | * | * | * | NS | ||
| FBP × AP | NS | * | ** | ** | NS | ** | * | ** | ||
| C × FBP × AP | NS | NS | NS | ** | NS | NS | ** | NS | ||
| Cultivar | Correlation with | Pn1 | Soluble Sugar | Sucrose | Starch | Tr 2 | Initial Rubisco 3 | cy-FBPase 4 | SPS 5 | SuSy 6 |
|---|---|---|---|---|---|---|---|---|---|---|
| CCRI-79 | Boll biomass | 0.466 | −0.708 ** 7 | −0.526 * | −0.854 ** | 0.780 ** | 0.486 * | −0.670 ** | 0.728 ** | 0.804 ** |
| Boll number | −0.615 ** | −0.755 ** | −0.047 | −0.335 | 0.095 | −0.300 | −0.383 | 0.570* | −0.253 | |
| Lint/seed ratio | 0.827 ** | 0.049 | 0.103 | −0.061 | 0.065 | 0.912 ** | 0.103 | 0.312 | 0.602 ** | |
| Lint yield | −0.256 | −0.888 ** | −0.035 | −0.467 | 0.196 | 0.083 | −0.443 | 0.831 ** | 0.633 ** | |
| SCRC-28 | Boll biomass | 0.470 * | −0.129 | −0.837 ** | −0.603 ** | 0.723 ** | 0.389 | −0.276 | 0.442 | 0.715 ** |
| Boll number | −0.546 * | −0.772 ** | −0.127 | −0.643 ** | 0.204 | −0.053 | −0.815 ** | 0.149 | 0.325 | |
| Lint/seed ratio | 0.182 | −0.781 ** | −0.602 ** | −0.686 ** | 0.192 | 0.684 ** | −0.526 * | 0.746 ** | 0.771 ** | |
| Lint yield | −0.357 | −0.814 ** | −0.324 | −0.757 ** | 0.313 | 0.161 | −0.869 ** | 0.377 | 0.499 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, M.; Li, P.; Wang, N.; Zheng, C.; Sun, X.; Dong, H.; Han, H.; Feng, W.; Shao, J.; Zhang, Y. Soil Available Phosphorus Deficiency Reduces Boll Biomass and Lint Yield by Affecting Sucrose Metabolism in Cotton-Boll Subtending Leaves. Agronomy 2022, 12, 1065. https://doi.org/10.3390/agronomy12051065
Sun M, Li P, Wang N, Zheng C, Sun X, Dong H, Han H, Feng W, Shao J, Zhang Y. Soil Available Phosphorus Deficiency Reduces Boll Biomass and Lint Yield by Affecting Sucrose Metabolism in Cotton-Boll Subtending Leaves. Agronomy. 2022; 12(5):1065. https://doi.org/10.3390/agronomy12051065
Chicago/Turabian StyleSun, Miao, Pengcheng Li, Ning Wang, Cangsong Zheng, Xuezhen Sun, Helin Dong, Huimin Han, Weina Feng, Jingjing Shao, and Yufeng Zhang. 2022. "Soil Available Phosphorus Deficiency Reduces Boll Biomass and Lint Yield by Affecting Sucrose Metabolism in Cotton-Boll Subtending Leaves" Agronomy 12, no. 5: 1065. https://doi.org/10.3390/agronomy12051065
APA StyleSun, M., Li, P., Wang, N., Zheng, C., Sun, X., Dong, H., Han, H., Feng, W., Shao, J., & Zhang, Y. (2022). Soil Available Phosphorus Deficiency Reduces Boll Biomass and Lint Yield by Affecting Sucrose Metabolism in Cotton-Boll Subtending Leaves. Agronomy, 12(5), 1065. https://doi.org/10.3390/agronomy12051065