
Genome-Wide Association Mapping Revealed SNP Alleles Associated with Spike Traits in Wheat


 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Experimental Design and Layout
2.3. Yield Related Traits
2.4. Statistical Analysis of the Studied Yield Components
2.5. DNA Extraction, Genotyping-By-Sequencing and SNP Calling
2.6. Genome-Wide Association Study (GWAS) for the Studied Yield Components
2.7. Candidate Genes and Gene Annotation for Yield Component Traits
3. Results
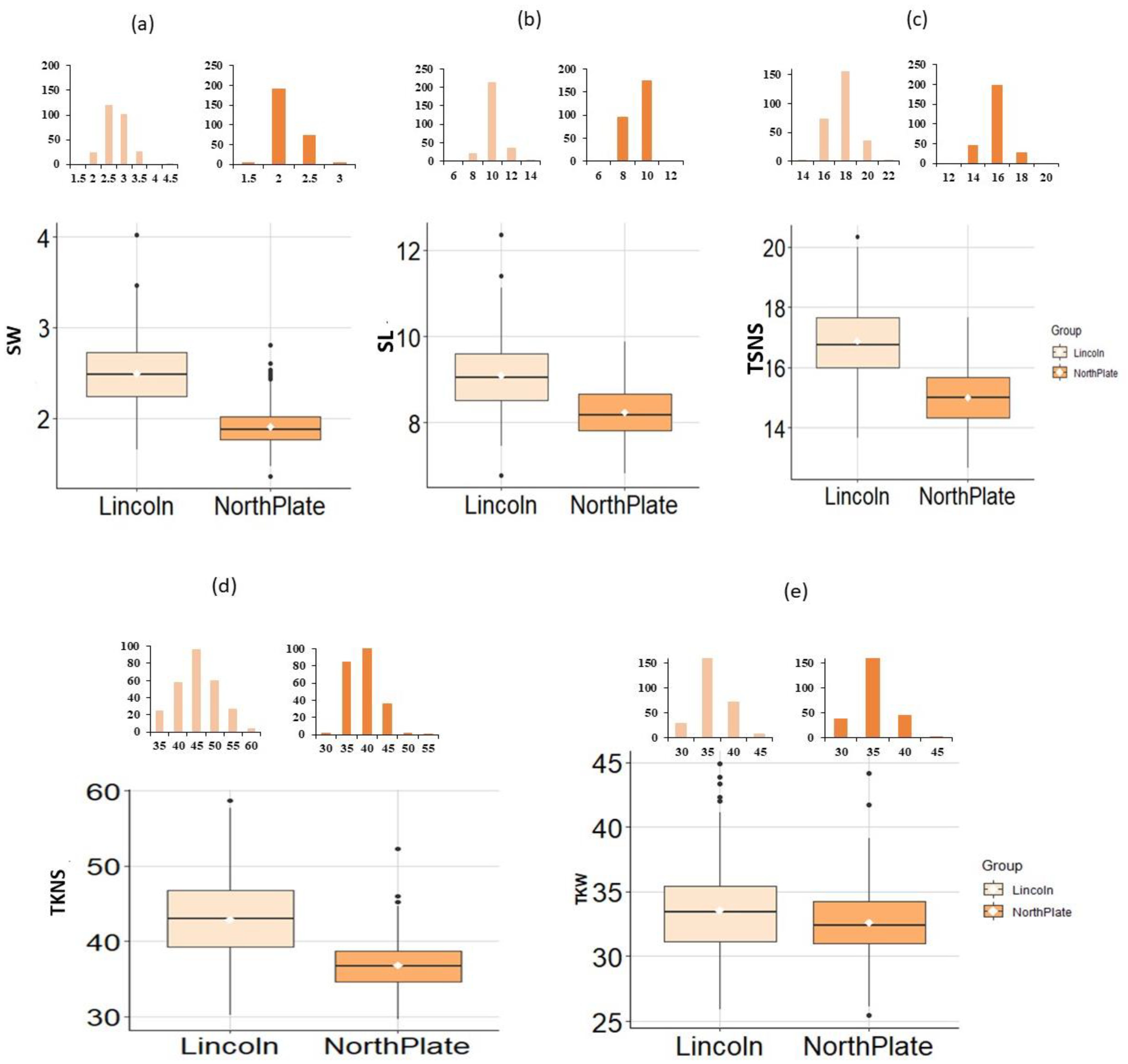
3.1. Analysis of Variance for the Yield Components Traits
3.2. Phenotypic Analysis for the Yield Related Traits
3.3. Correlation Coefficients for Yield-Related Traits
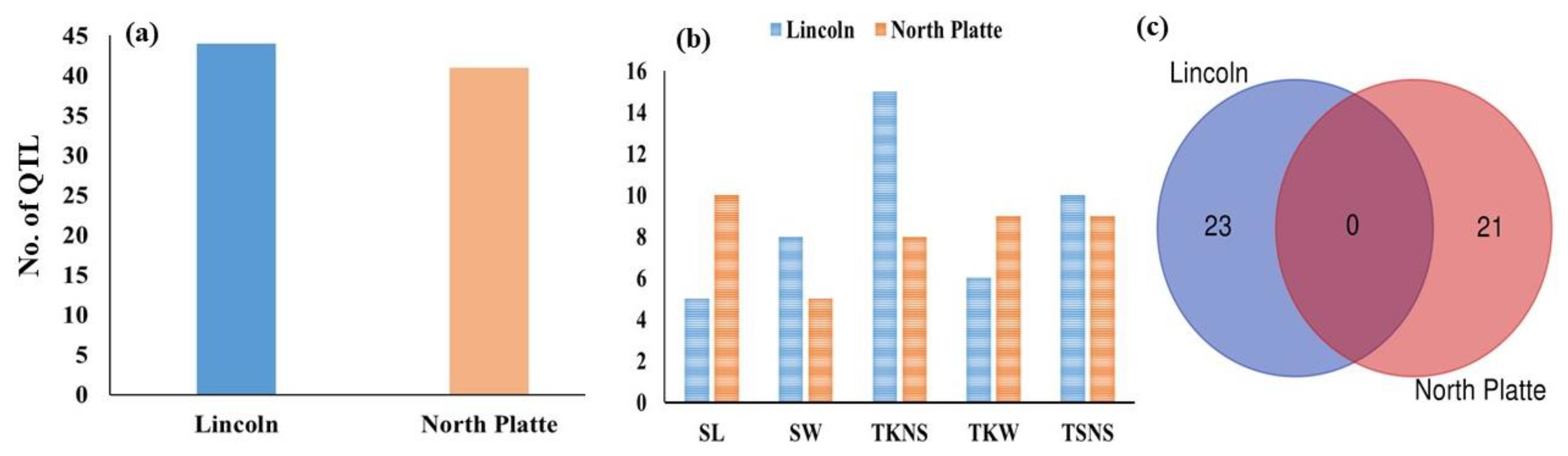
3.4. Genome Wide Association Studies for Yield-Related Traits
3.5. Common Markers Associated with Yield-Related Traits
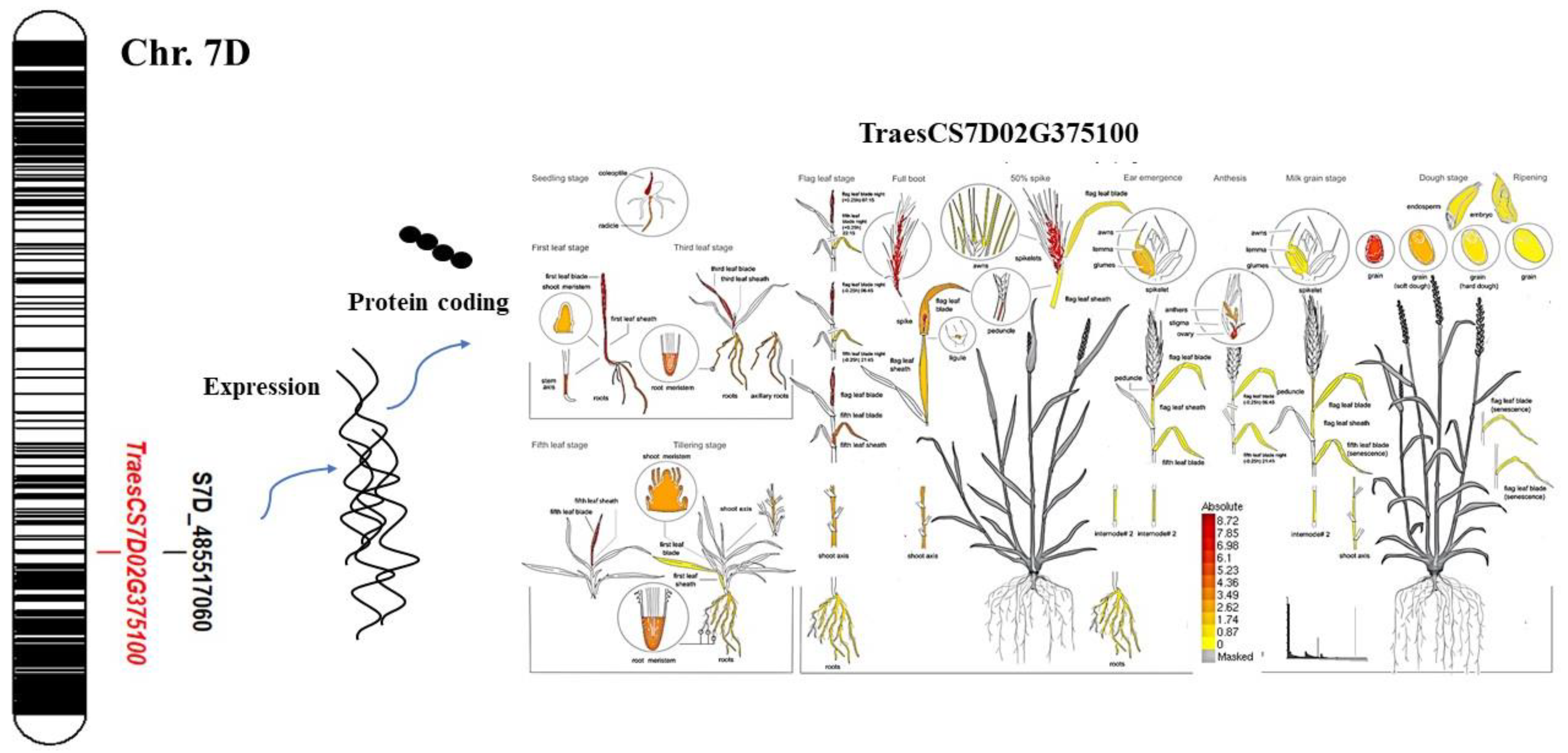
3.6. Gene Annotation for Yield-Related Traits
4. Discussion
4.1. Genetic Variation for Yield-Related Traits
4.2. Genome Wide Association Mapping for Yield-Related Traits
4.3. Gene Annotation for Yield-Related Traits
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ward, B.P.; Brown-Guedira, G.; Kolb, F.L.; Van Sanford, D.A.; Tyagi, P.; Sneller, C.H.; Griffey, C.A. Genome-Wide Association Studies for Yield-Related Traits in Soft Red Winter Wheat Grown in Virginia. PLoS ONE 2019, 14, e0208217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondal, S.; Sallam, A.; Sehgal, D.; Sukumaran, S.; Farhad, M.; Navaneetha Krishnan, J.; Kumar, U.; Biswal, A. Advances in Breeding for Abiotic Stress Tolerance in Wheat. In Genomic Designing for Abiotic Stress Resistant Cereal Crops; Springer: Cham, Switzerland, 2021; pp. 71–103. [Google Scholar]
- Mourad, A.M.I.; Alomari, D.Z.; Alqudah, A.M.; Sallam, A.; Salem, K.F.M. Recent Advances in Wheat (Triticum Spp.) Breeding. In Advances in Plant Breeding Strategies: Cereals; Springer: Cham, Switzerland, 2019; Volume 5. [Google Scholar] [CrossRef]
- Sun, C.; Zhang, F.; Yan, X.; Zhang, X.; Dong, Z.; Cui, D.; Chen, F. Genome-Wide Association Study for 13 Agronomic Traits Reveals Distribution of Superior Alleles in Bread Wheat from the Yellow and Huai Valley of China. Plant Biotechnol. J. 2017, 15, 953–969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sallam, A.; Hashad, M.; Hamed, E.-S.; Omara, M. Genetic Variation of Stem Characters in Wheat and Their Relation to Kernel Weight under Drought and Heat Stresses. J. Crop Sci. Biotechnol. 2015, 18, 137–146. [Google Scholar] [CrossRef]
- Mourad, A.M.I.; Belamkar, V.; Baenziger, P.S. Molecular Genetic Analysis of Spring Wheat Core Collection Using Genetic Diversity, Population Structure, and Linkage Disequilibrium. BMC Genom. 2020, 21, 434. [Google Scholar] [CrossRef]
- Li, F.; Wen, W.; Liu, J.; Zhang, Y.; Cao, S.; He, Z.; Rasheed, A.; Jin, H.; Zhang, C.; Yan, J.; et al. Genetic Architecture of Grain Yield in Bread Wheat Based on Genome-Wide Association Studies. BMC Plant Biol. 2019, 19, 168. [Google Scholar] [CrossRef] [PubMed]
- Nadolska-Orczyk, A.; Rajchel, I.K.; Orczyk, W.; Gasparis, S. Major Genes Determining Yield-Related Traits in Wheat and Barley. Theor. Appl. Genet. 2017, 130, 1081–1098. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Wen, W.; He, Z.; Liu, J.; Jin, H.; Cao, S.; Geng, H.; Yan, J.; Zhang, P.; Wan, Y.; et al. Genome-Wide Linkage Mapping of Yield-Related Traits in Three Chinese Bread Wheat Populations Using High-Density SNP Markers. Theor. Appl. Genet. 2018, 131, 1903–1924. [Google Scholar] [CrossRef]
- Rasheed, A.; Wen, W.; Gao, F.; Zhai, S.; Jin, H.; Liu, J.; Guo, Q.; Zhang, Y.; Dreisigacker, S.; Xia, X.; et al. Development and Validation of KASP Assays for Genes Underpinning Key Economic Traits in Bread Wheat. Theor. Appl. Genet. 2016, 129, 1843–1860. [Google Scholar] [CrossRef]
- Gao, F.; Wen, W.; Liu, J.; Rasheed, A.; Yin, G.; Xia, X.; Wu, X.; He, Z. Genome-Wide Linkage Mapping of QTL for Yield Components, Plant Height and Yield-Related Physiological Traits in the Chinese Wheat Cross Zhou 8425B/Chinese Spring. Front. Plant Sci. 2015, 6, 1099. [Google Scholar] [CrossRef] [Green Version]
- Cuthbert, J.L.; Somers, D.J.; Brûlé-Babel, A.L.; Brown, P.D.; Crow, G.H.; Liu, J.; He, Z.; Rasheed, A.; Wen, W.; Yan, J.; et al. A Multiparental Cross Population for Mapping QTL for Agronomic Traits in Durum Wheat (T Riticum Turgidum Ssp. Durum). Theor. Appl. Genet. 2016, 130, 803–807. [Google Scholar]
- Cui, F.; Zhao, C.; Ding, A.; Li, J.; Wang, L.; Li, X.; Bao, Y.; Li, J.; Wang, H. Construction of an Integrative Linkage Map and QTL Mapping of Grain Yield-Related Traits Using Three Related Wheat RIL Populations. Theor. Appl. Genet. 2014, 127, 659–675. [Google Scholar] [CrossRef] [PubMed]
- Cui, F.; Li, J.; Ding, A.; Zhao, C.; Wang, L.; Wang, X.; Li, S.; Bao, Y.; Li, X.; Feng, D.; et al. Conditional QTL Mapping for Plant Height with Respect to the Length of the Spike and Internode in Two Mapping Populations of Wheat. Theor. Appl. Genet. 2011, 122, 1517–1536. [Google Scholar] [CrossRef] [PubMed]
- Buerstmayr, H.; Ban, T.; Anderson, J.A. QTL Mapping and Marker-Assisted Selection for Fusarium Head Blight Resistance in Wheat: A Review. Plant Breed. 2009, 128, 1–26. [Google Scholar] [CrossRef]
- Collard, B.C.Y.; Jahufer, M.Z.Z.; Brouwer, J.B.; Pang, E.C.K. An Introduction to Markers, Quantitative Trait Loci (QTL) Mapping and Marker-Assisted Selection for Crop Improvement: The Basic Concepts. Euphytica 2005, 142, 169–196. [Google Scholar] [CrossRef]
- Brondani, C.; Rangel, P.; Brondani, R.; Ferreira, M. QTL Mapping, and Introgression of Yield-Related Traits from Oryza Glumaepatula to Cultivated Rice (Oryza Sativa) Using Microsatellite Markers. Theor. Appl. Genet. 2002, 104, 1192–1203. [Google Scholar] [CrossRef] [PubMed]
- Eltaher, S.; Baenziger, P.S.; Belamkar, V.; Emara, H.A.; Nower, A.A.; Salem, K.F.M.; Alqudah, A.M.; Sallam, A. GWAS Revealed Effect of Genotype × Environment Interactions for Grain Yield of Nebraska Winter Wheat. BMC Genom. 2021, 22, 2. [Google Scholar] [CrossRef]
- Sallam, A.; Amro, A.; Elakhdar, A.; Dawood, M.F.A.; Moursi, Y.S.; Baenziger, P.S. Marker--Trait Association for Grain Weight of Spring Barley in Well-Watered and Drought Environments. Mol. Biol. Rep. 2019, 46, 2907–2918. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Nie, X.; Shen, C.; You, C.; Li, W.; Zhao, W.; Zhang, X.; Lin, Z. Population Structure and Genetic Basis of the Agronomic Traits of Upland Cotton in China Revealed by a Genome-Wide Association Study Using High-Density SNP S. Plant Biotechnol. J. 2017, 15, 1374–1386. [Google Scholar] [CrossRef]
- Sallam, A.; Eltaher, S.; Alqudah, A.M.; Belamkar, V.; Baenziger, P.S. Combined GWAS and QTL Mapping Revealed Candidate Genes and SNP Network Controlling Recovery and Tolerance Traits Associated with Drought Tolerance in Seedling Winter Wheat. Genomics 2022, 114, 110358. [Google Scholar] [CrossRef]
- Scherer, A.; Christensen, G.B. Concepts and Relevance of Genome-Wide Association Studies. Sci. Prog. 2016, 99, 59–67. [Google Scholar] [CrossRef]
- Bajgain, P.; Zhang, X.; Anderson, J.A. Genome-Wide Association Study of Yield Component Traits in Intermediate Wheatgrass and Implications in Genomic Selection and Breeding. G3 Genes Genomes Genet. 2019, 9, 2429–2439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gahlaut, V.; Jaiswal, V.; Singh, S.; Balyan, H.S.; Gupta, P.K. Multi-Locus Genome Wide Association Mapping for Yield and Its Contributing Traits in Hexaploid Wheat under Different Water Regimes. Sci. Rep. 2019, 9, 19486. [Google Scholar] [CrossRef]
- Milner, S.G.; Maccaferri, M.; Huang, B.E.; Mantovani, P.; Massi, A.; Frascaroli, E.; Tuberosa, R.; Salvi, S. A Multiparental Cross Population for Mapping QTL for Agronomic Traits in Durum Wheat (T Riticum Turgidum Ssp. Durum). Plant Biotechnol. J. 2016, 14, 735–748. [Google Scholar] [CrossRef] [Green Version]
- Jia, H.; Wan, H.; Yang, S.; Zhang, Z.; Kong, Z.; Xue, S.; Zhang, L.; Ma, Z. Genetic Dissection of Yield-Related Traits in a Recombinant Inbred Line Population Created Using a Key Breeding Parent in China’s Wheat Breeding. Theor. Appl. Genet. 2013, 126, 2123–2139. [Google Scholar] [CrossRef]
- Azadi, A.; Mardi, M.; Hervan, E.M.; Mohammadi, S.A.; Moradi, F.; Tabatabaee, M.T.; Pirseyedi, S.M.; Ebrahimi, M.; Fayaz, F.; Kazemi, M.; et al. QTL Mapping of Yield and Yield Components under Normal and Salt-Stress Conditions in Bread Wheat (Triticum Aestivum L.). Plant Mol. Biol. Report. 2015, 33, 102–120. [Google Scholar] [CrossRef]
- Cuthbert, J.L.; Somers, D.J.; Brûlé-Babel, A.L.; Brown, P.D.; Crow, G.H. Molecular Mapping of Quantitative Trait Loci for Yield and Yield Components in Spring Wheat (Triticum Aestivum L.). Theor. Appl. Genet. 2008, 117, 595–608. [Google Scholar] [CrossRef] [PubMed]
- Eltaher, S.; Sallam, A.; Belamkar, V.; Emara, H.A.; Nower, A.A.; Salem, K.F.M.; Poland, J.; Baenziger, P.S. Genetic Diversity and Population Structure of F3:6 Nebraska Winter Wheat Genotypes Using Genotyping-By-Sequencing. Front. Genet. 2018, 9, 76. [Google Scholar] [CrossRef]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A Robust, Simple Genotyping-by-Sequencing (GBS) Approach for High Diversity Species. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef] [Green Version]
- Poland, J.A.; Brown, P.J.; Sorrells, M.E.; Jannink, J.-L. Development of High-Density Genetic Maps for Barley and Wheat Using a Novel Two-Enzyme Genotyping-by-Sequencing Approach. PLoS ONE 2012, 7, e32253. [Google Scholar] [CrossRef] [Green Version]
- Poland, J.; Endelman, J.; Dawson, J.; Rutkoski, J.; Wu, S.; Manes, Y.; Dreisigacker, S.; Crossa, J.; Sánchez-Villeda, H.; Sorrells, M.; et al. Genomic Selection in Wheat Breeding Using Genotyping-by-Sequencing. Plant Genome 2012, 5, 103–113. [Google Scholar] [CrossRef] [Green Version]
- Lorenz, A.J.; Hamblin, M.T.; Jannink, J.-L. Performance of Single Nucleotide Polymorphisms versus Haplotypes for Genome-Wide Association Analysis in Barley. PLoS ONE 2010, 5, e14079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chia, J.-M.; Song, C.; Bradbury, P.J.; Costich, D.; De Leon, N.; Doebley, J.; Elshire, R.J.; Gaut, B.; Geller, L.; Glaubitz, J.C.; et al. Maize HapMap2 Identifies Extant Variation from a Genome in Flux. Nat. Genet. 2012, 44, 803–807. [Google Scholar] [CrossRef] [PubMed]
- Thomson, M.J. High-Throughput SNP Genotyping to Accelerate Crop Improvement. Plant Breed. Biotechnol. 2014, 2, 195–212. [Google Scholar] [CrossRef]
- Eltaher, S.; Mourad, A.M.I.; Baenziger, P.S.; Wegulo, S.; Belamkar, V.; Sallam, A. Identification and Validation of High LD Hotspot Genomic Regions Harboring Stem Rust Resistant Genes on 1B, 2A (Sr38), and 7B Chromosomes in Wheat. Front. Genet. 2021, 1875. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for Association Mapping of Complex Traits in Diverse Samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef]
- Zhang, X.; Pérez-Rodríguez, P.; Semagn, K.; Beyene, Y.; Babu, R.; López-Cruz, M.A.; San Vicente, F.; Olsen, M.; Buckler, E.; Jannink, J.-L.; et al. Genomic Prediction in Biparental Tropical Maize Populations in Water-Stressed and Well-Watered Environments Using Low-Density and GBS SNPs. Heredity 2015, 114, 291–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belamkar, V.; Guttieri, M.J.; Hussain, W.; Jarquin, D.; El-basyoni, I.; Poland, J.; Lorenz, A.J.; Baenziger, P.S. Genomic Selection in Preliminary Yield Trials in a Winter Wheat Breeding Program. G3 Genes Genomes Genet. 2018, 8, 2735–2747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, W.; Baenziger, P.S.; Belamkar, V.; Guttieri, M.J.; Venegas, J.P.; Easterly, A.; Sallam, A.; Poland, J. Genotyping-by-Sequencing Derived High-Density Linkage Map and Its Application to QTL Mapping of Flag Leaf Traits in Bread Wheat. Sci. Rep. 2017, 7, 16394. [Google Scholar] [CrossRef]
- Alqudaha, A.M.; Sallamb, A.; Baenzigerc, P.S.; Börnera, A. GWAS: Fast-Forwarding Gene Identification in Temperate Cereals: Barley as a Case Study-A Review. J. Adv. Res. 2019, 22, 119–135. [Google Scholar] [CrossRef]
- Mourad, A.M.I.; Sallam, A.; Belamkar, V.; Wegulo, S.; Bowden, R.; Jin, Y.; Mahdy, E.; Bakheit, B.; El-Wafaa, A.A.; Poland, J.; et al. Genome-Wide Association Study for Identification and Validation of Novel SNP Markers for Sr6 Stem Rust Resistance Gene in Bread Wheat. Front. Plant Sci. 2018, 9, 380. [Google Scholar] [CrossRef] [Green Version]
- Mohammadi, R.; Haghparast, R.; Amri, A.; Ceccarelli, S. Yield Stability of Rainfed Durum Wheat and GGE Biplot Analysis of Multi-Environment Trials. Crop Pasture Sci. 2009, 61, 92–101. [Google Scholar] [CrossRef]
- Mohammadi, R.; Amri, A. Genotype × Environment Interaction and Genetic Improvement for Yield and Yield Stability of Rainfed Durum Wheat in Iran. Euphytica 2013, 192, 227–249. [Google Scholar] [CrossRef]
- Mehari, M.; Tesfay, M.; Yirga, H.; Mesele, A.; Abebe, T.; Workineh, A.; Amare, B. GGE Biplot Analysis of Genotype-by-Environment Interaction and Grain Yield Stability of Bread Wheat Genotypes in South Tigray, Ethiopia. Commun. Biometry Crop Sci. 2015, 10, 17–26. [Google Scholar]
- Matlala, M.; Shimelis, H.; Mashilo, J. Genotype-by-Environment Interaction of Grain Yield among Candidate Dryland Wheat Genotypes. South Afr. J. Plant Soil 2019, 36, 299–306. [Google Scholar] [CrossRef]
- Agrama, H.A.; Eizenga, G.C.; Yan, W. Association Mapping of Yield, and Its Components in Rice Cultivars. Mol. Breed. 2007, 19, 341–356. [Google Scholar] [CrossRef]
- Tsilo, T.J.; Hareland, G.A.; Simsek, S.; Chao, S.; Anderson, J.A. Genome Mapping of Kernel Characteristics in Hard Red Spring Wheat Breeding Lines. Theor. Appl. Genet. 2010, 121, 717–730. [Google Scholar] [CrossRef]
- Edae, E.A.; Byrne, P.F.; Haley, S.D.; Lopes, M.S.; Reynolds, M.P. Genome-Wide Association Mapping of Yield and Yield Components of Spring Wheat under Contrasting Moisture Regimes. Theor. Appl. Genet. 2014, 127, 791–807. [Google Scholar] [CrossRef]
- Bhusal, N.; Sarial, A.K.; Sharma, P.; Sareen, S. Mapping QTLs for Grain Yield Components in Wheat under Heat Stress. PLoS ONE 2017, 12, e0189594. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Bowman, B.C.; Hu, Y.-G.; Liang, X.; Zhao, W.; Wheeler, J.; Klassen, N.; Bockelman, H.; Bonman, J.M.; Chen, J. Evaluation of Agronomic Traits and Drought Tolerance of Winter Wheat Accessions from the USDA-ARS National Small Grains Collection. Agronomy 2017, 7, 51. [Google Scholar] [CrossRef]
- Mwadzingeni, L.; Shimelis, H.; Rees, D.J.G.; Tsilo, T.J. Genome-Wide Association Analysis of Agronomic Traits in Wheat under Drought-Stressed and Non-Stressed Conditions. PLoS ONE 2017, 12, e0171692. [Google Scholar] [CrossRef] [Green Version]
- Sallam, A.; Alqudah, A.M.; Dawood, M.F.A.; Baenziger, P.S.; Börner, A. Drought Stress Tolerance in Wheat and Barley: Advances in Physiology, Breeding and Genetics Research. Int. J. Mol. Sci. 2019, 20, 3137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dawood, M.F.A.; Moursi, Y.S.; Amro, A.; Baenziger, P.S.; Sallam, A. Investigation of Heat-Induced Changes in the Grain Yield and Grains Metabolites, with Molecular Insights on the Candidate Genes in Barley. Agronomy 2020, 10, 1730. [Google Scholar] [CrossRef]
- Liu, G.; Jia, L.; Lu, L.; Qin, D.; Zhang, J.; Guan, P.; Ni, Z.; Yao, Y.; Sun, Q.; Peng, H. Mapping QTLs of Yield-Related Traits Using RIL Population Derived from Common Wheat and Tibetan Semi-Wild Wheat. Theor. Appl. Genet. 2014, 127, 2415–2432. [Google Scholar] [CrossRef] [PubMed]
- Zanke, C.D.; Ling, J.; Plieske, J.; Kollers, S.; Ebmeyer, E.; Korzun, V.; Argillier, O.; Stiewe, G.; Hinze, M.; Neumann, K.; et al. Whole Genome Association Mapping of Plant Height in Winter Wheat (Triticum Aestivum L.). PLoS ONE 2014, 9, e113287. [Google Scholar]
- Zanke, C.D.; Ling, J.; Plieske, J.; Kollers, S.; Ebmeyer, E.; Korzun, V.; Argillier, O.; Stiewe, G.; Hinze, M.; Neumann, F.; et al. Analysis of Main Effect QTL for Thousand Grain Weight in European Winter Wheat (Triticum Aestivum L.) by Genome-Wide Association Mapping. Front. Plant Sci. 2015, 6, 644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.; Zhang, H.; Deng, Z.; Wu, R.; Li, D.; Wang, M.; Tian, J. Genome-Wide Association Study for Kernel Weight-Related Traits Using SNPs in a Chinese Winter Wheat Population. Euphytica 2016, 212, 173–185. [Google Scholar] [CrossRef]
- Lopes, M.S.; Reynolds, M.P.; McIntyre, C.L.; Mathews, K.L.; Kamali, M.R.J.; Mossad, M.; Feltaous, Y.; Tahir, I.S.A.; Chatrath, R.; Ogbonnaya, F.; et al. QTL for Yield and Associated Traits in the Seri/Babax Population Grown across Several Environments in Mexico, in the West Asia, North Africa, and South Asia Regions. Theor. Appl. Genet. 2013, 126, 971–984. [Google Scholar] [CrossRef]
- Wang, S.-X.; Zhu, Y.-L.; Zhang, D.-X.; Shao, H.; Liu, P.; Hu, J.-B.; Zhang, H.; Zhang, H.-P.; Chang, C.; Lu, J.; et al. Genome-Wide Association Study for Grain Yield and Related Traits in Elite Wheat Varieties and Advanced Lines Using SNP Markers. PLoS ONE 2017, 12, e0188662. [Google Scholar] [CrossRef] [Green Version]
- Narasimhamoorthy, B.; Gill, B.S.; Fritz, A.K.; Nelson, J.C.; Brown-Guedira, G.L. Advanced Backcross QTL Analysis of a Hard Winter Wheat × Synthetic Wheat Population. Theor. Appl. Genet. 2006, 112, 787–796. [Google Scholar] [CrossRef]
- Mago, R.; Tabe, L.; McIntosh, R.A.; Pretorius, Z.; Kota, R.; Paux, E.; Wicker, T.; Breen, J.; Lagudah, E.S.; Ellis, J.G.; et al. A Multiple Resistance Locus on Chromosome Arm 3BS in Wheat Confers Resistance to Stem Rust (Sr2), Leaf Rust (Lr27) and Powdery Mildew. Theor. Appl. Genet. 2011, 123, 615–623. [Google Scholar] [CrossRef]
- Bednarek, J.; Boulaflous, A.; Girousse, C.; Ravel, C.; Tassy, C.; Barret, P.; Bouzidi, M.F.; Mouzeyar, S. Down-Regulation of the TaGW2 Gene by RNA Interference Results in Decreased Grain Size and Weight in Wheat. J. Exp. Bot. 2012, 63, 5945–5955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, D.; Yan, J.; He, Z.; Wu, L.; Xia, X. Characterization of a Cell Wall Invertase Gene TaCwi-A1 on Common Wheat Chromosome 2A and Development of Functional Markers. Mol. Breed. 2012, 29, 43–52. [Google Scholar] [CrossRef]
- Hou, J.; Jiang, Q.; Hao, C.; Wang, Y.; Zhang, H.; Zhang, X. Global Selection on Sucrose Synthase Haplotypes during a Century of Wheat Breeding. Plant Physiol. 2014, 164, 1918–1929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laido, G.; Marone, D.; Russo, M.A.; Colecchia, S.A.; Mastrangelo, A.M.; De Vita, P.; Papa, R. Linkage Disequilibrium and Genome-Wide Association Mapping in Tetraploid Wheat (Triticum Turgidum L.). PLoS ONE 2014, 9, e95211. [Google Scholar] [CrossRef] [Green Version]
- Dunwell, J.M.; Purvis, A.; Khuri, S. Cupins: The Most Functionally Diverse Protein Superfamily? Phytochemistry 2004, 65, 7–17. [Google Scholar] [CrossRef]
- Afzal, M.; Sielaff, M.; Curella, V.; Neerukonda, M.; El Hassouni, K.; Schuppan, D.; Tenzer, S.; Longin, C.F.H. Characterization of 150 Wheat Cultivars by LC-MS-Based Label-Free Quantitative Proteomics Unravels Possibilities to Design Wheat Better for Baking Quality and Human Health. Plants 2021, 10, 424. [Google Scholar] [CrossRef]
- Theobald, D.L.; Mitton-Fry, R.M.; Wuttke, D.S. Nucleic Acid Recognition by OB-Fold Proteins. Annu. Rev. Biophys. Biomol. Struct. 2003, 32, 115–133. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Chen, X.; Chen, Z.; Cai, R.; Zhang, H.; Xiang, Y. Identification and Expression Analysis of BURP Domain-Containing Genes in Medicago Truncatula. Front. Plant Sci. 2016, 7, 485. [Google Scholar] [CrossRef] [Green Version]
- Ding, X.; Hou, X.; Xie, K.; Xiong, L. Genome-Wide Identification of BURP Domain-Containing Genes in Rice Reveals a Gene Family with Diverse Structures and Responses to Abiotic Stresses. Planta 2009, 230, 149–163. [Google Scholar] [CrossRef]
- Xu, H.; Li, Y.; Yan, Y.; Wang, K.; Gao, Y.; Hu, Y. Genome-Scale Identification of Soybean BURP Domain-Containing Genes and Their Expression under Stress Treatments. BMC Plant Biol. 2010, 10, 197. [Google Scholar] [CrossRef] [Green Version]
- Gan, D.; Jiang, H.; Zhang, J.; Zhao, Y.; Zhu, S.; Cheng, B. Genome-Wide Analysis of BURP Domain-Containing Genes in Maize and Sorghum. Mol. Biol. Rep. 2011, 38, 4553–4563. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Wei, G.; Wang, L.; Dong, Q.; Zhao, Y.; Chen, B.; Xiang, Y. Genome-Wide Analysis of BURP Domain-Containing Genes in Populus Trichocarpa. J. Integr. Plant Biol. 2011, 53, 743–755. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Cao, Y.; Qiu, J.; Gao, Z.; Ou, Z.; Wang, Y.; Zheng, Y. Expression of a Vacuole-Localized BURP-Domain Protein from Soybean (SALI3-2) Enhances Tolerance to Cadmium and Copper Stresses. PLoS ONE 2014, 9, e98830. [Google Scholar] [CrossRef] [PubMed]
- Bassüner, R.; Bäumlein, H.; Huth, A.; Jung, R.; Wobus, U.; Rapoport, T.A.; Saalbach, G.; Müntz, K. Abundant Embryonic MRNA in Field Bean (Vicia Faba L.) Codes for a New Class of Seed Proteins: CDNA Cloning and Characterization of the Primary Translation Product. Plant Mol. Biol. 1988, 11, 321–334. [Google Scholar] [CrossRef]
- Chesnokov, Y.V.; Meister, A.; Manteuffel, R. A Chimeric Green Fluorescent Protein Gene as an Embryogenic Marker in Transgenic Cell Culture of Nicotiana Plumbaginifolia Viv. Plant Sci. 2002, 162, 59–77. [Google Scholar] [CrossRef]
- Hanks, S.K.; Hunter, T. The Eukaryotic Protein Kinase Superfamily: Kinase (Catalytic) Domain Structure and Classification 1. FASEB J. 1995, 9, 576–596. [Google Scholar] [CrossRef]
- Stone, J.M.; Walker, J.C. Plant Protein Kinase Families and Signal Transduction. Plant Physiol. 1995, 108, 451–457. [Google Scholar] [CrossRef] [Green Version]
- Lehti-Shiu, M.D.; Shiu, S.-H. Diversity, Classification and Function of the Plant Protein Kinase Superfamily. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 2619–2639. [Google Scholar]
- Saidi, Y.; Hearn, T.J.; Coates, J.C. Function and Evolution of ‘green’ GSK3/Shaggy-like Kinases. Trends Plant Sci. 2012, 17, 39–46. [Google Scholar] [CrossRef]
- Jelsema, C.L.; Morre, D.J.; Ruddat, M. Phospholipid Metabolism of Wheat Grains: Phospholipid Composition and Terminal Enzymes of the CDP-Amine Phospholipid Biosynthetic Pathway. Bot. Gaz. 1982, 143, 26–31. [Google Scholar] [CrossRef]
- González-Thuillier, I.; Pellny, T.K.; Tosi, P.; Mitchell, R.A.C.; Haslam, R.; Shewry, P.R. Accumulation and Deposition of Triacylglycerols in the Starchy Endosperm of Wheat Grain. J. Cereal Sci. 2021, 98, 103167. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Thuillier, I.; Salt, L.; Chope, G.; Penson, S.; Skeggs, P.; Tosi, P.; Powers, S.J.; Ward, J.L.; Wilde, P.; Shewry, P.R.; et al. Distribution of Lipids in the Grain of Wheat (Cv. Hereward) Determined by Lipidomic Analysis of Milling and Pearling Fractions. J. Agric. Food Chem. 2015, 63, 10705–10716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brock, C.J. Wheat: Production Properties and Quality; Bushuk, W., Rasper, V.F., Eds.; Blackie/Chapman and Hall: London, UK, 1994; ISBN 0-7514-0181-1. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | SW | SL | TNSN | TKNS | TKW | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean of Square | Pr (>F) | Mean of Square | Pr (>F) | Mean of Square | Pr (>F) | Mean of Square | Pr (>F) | Mean of Square | Pr (>F) | |
| Genotypes | 0.5599 | 2.2 × 10−15 *** | 3.079 | 2.2 × 10−15 *** | 6.55 | 2.2 × 10−15 *** | 118.86 | 2.2 × 10−15 *** | 16.819 | 6.74 × 10−5 *** |
| Environment | 18.0789 | 0.0001648 *** | 76.152 | 0.000273 *** | 671.28 | 1.09 × 10−5 *** | 788.51 | 0.00824 *** | 39.589 | 4.57 × 10−5 *** |
| Iblock (Environment) | 0.4371 | 0.1797082 | 1.889 | 0.239888 | 9.87 | 0.07659 | 96.45 | 0.20626 | 26.308 | 0.372 |
| Genotypes × Environment | 0.583 | <2.2 × 10−15 *** | 2.986 | 2.2 × 10−15 *** | 6.94 | 2.2 × 10−15 *** | 144.16 | 2.2 × 10−15 *** | 16.836 | 6.46 × 10−5 *** |
| Residuals | 0.22 | 1.14 | 2.8 | 55 | 13.145 | |||||
| Heritability | 72.6 | 72.3 | 71.2 | 72.3 | 56.1 | |||||
| Traits | Number of SNPs | Log 10 p Value | Range of R2 | Range of Allele Effect | |||
|---|---|---|---|---|---|---|---|
| Min | Max | Min R2 | Max R2 | Min Allele Effect | Max Allele Effect | ||
| SW | 8 | 3.63 × 10−7 | 3.27 × 10−5 | 5.75 | 8.87 | 0.19 A | 0.42 A |
| SL | 5 | 1.16 × 10−7 | 1.17 × 10−5 | 6.07 | 9.01 | 0.43 T | 0.62 T |
| TSNS | 10 | 3.63 × 10−7 | 1.01 × 10−5 | 6.03 | 8.69 | 0.64 T | 1.44 A |
| TKNS | 15 | 3.63 × 10−7 | 3.27 × 10−5 | 5.70 | 8.96 | 2.90 G | 6.71 A |
| TKW | 6 | 1.1 × 10−6 | 1.77 × 10−5 | 5.92 | 8.74 | 2.40 C | 2.89 C |
| Traits | Number of SNPs | Log 10 p Value | Range of R2 | Range of Allele Effect | |||
|---|---|---|---|---|---|---|---|
| Min | Max | Min R2 | Max R2 | Min Allele Effect | Max Allele Effect | ||
| SW | 5 | 3.72 × 10−7 | 2.02 × 10−5 | 6.91 | 8.26 | 0.40 C | 0.65 C |
| SL | 10 | 3.91 × 10−7 | 3.06 × 10−5 | 5.90 | 8.36 | 0.12 T | 0.25 A |
| TSNS | 9 | 3.72 × 10−7 | 2.67 × 10−5 | 5.45 | 8.03 | 0.46 G | 1.55 C |
| TKNS | 8 | 3.91 × 10−7 | 2.11 × 10−5 | 7.00 | 13.36 | 1.85 A | 3.06 C |
| TKW | 9 | 1.58 × 10−7 | 1.88 × 10−5 | 9.54 | 12.03 | 1.44 G | 2.84 T |
| Traits | Marker ID | Chromosome | Position | Log 10 p Value | R2 | Target Allele | Allele Effect |
|---|---|---|---|---|---|---|---|
| SW | S5A_380823821 | 5A | 380823821 | 1.17 × 10−5 | 6.06 | T | 0.43 |
| SL | 4.8 × 10−6 | 5.9 | 0.19 | ||||
| TNKS | 1.06 × 10−5 | 6.32 | 3.06 | ||||
| TSNS | 1.01 × 10−5 | 6.03 | 0.64 | ||||
| SW | S5A_46628103 | 5A | 46628103 | 4.68 × 10−7 | 5.82 | A | 0.19 |
| TNKS | 3.27 × 10−5 | 6.47 | 3.00 | ||||
| SW | S5D_548379143 | 5D | 548379143 | 1.05 × 10−6 | 9.11 | T | 0.50 |
| SL | 3.63 × 10−7 | 8.87 | 0.23 | ||||
| TNKS | 3.63 × 10−7 | 8.96 | 3.53 | ||||
| TSNS | 3.63 × 10−7 | 8.69 | 0.74 | ||||
| TNKS | S6D_469537865 | 6D | 469537865 | 4.8 × 10−6 | 6.53 | A | 6.71 |
| TSNS | 1.06 × 10−5 | 6.56 | 1.44 | ||||
| TNKS | S7B_164151731 | 7B | 164151731 | 1.06 × 10−5 | 5.87 | C | 3.46 |
| TSNS | 1.29 × 10−5 | 6.51 | 0.78 | ||||
| SW | S7B_165529101 | 7B | 165529101 | 1.16 × 10−7 | 6.19 | C | 0.49 |
| SL | 1.06 × 10−5 | 5.78 | 0.22 | ||||
| TNKS | 7.8 × 10−6 | 6.50 | 3.52 | ||||
| TSNS | 4.68 × 10−6 | 7.12 | 0.79 | ||||
| TNKS | S7B_181032630 | 7B | 181032630 | 1.29 × 10−5 | 6.17 | A | 3.50 |
| TSNS | 4.8 × 10−6 | 6.84 | 0.79 | ||||
| SW | S7B_329792071 | 7B | 329792071 | 5.5 × 10−6 | 6.91 | A | 0.54 |
| SL | 1.29 × 10−5 | 6.56 | 0.24 | ||||
| TNKS | 4.68 × 10−6 | 6.87 | 3.80 | ||||
| TSNS | 3.77 × 10−6 | 7.79 | 0.87 | ||||
| SW | S7B_607427421 | 7B | 607427421 | 3.27 × 10−6 | 5.70 | G | 0.19 |
| TNKS | 1.29 × 10−6 | 5.70 | 2.90 | ||||
| SW | S7D_485517060 | 7D | 485517060 | 4.83 × 10−6 | 6.67 | T | 0.62 |
| SL | 3.77 × 10−6 | 6.61 | 0.28 | ||||
| TNKS | 3.77 × 10−6 | 7.01 | 4.43 | ||||
| TSNS | 7.8 × 10−6 | 6.67 | 0.92 |
| Trait | Marker ID | Chromosome | Position | Log 10 p Value | R2 | Target Allele | Allele Effect |
|---|---|---|---|---|---|---|---|
| SL | S3B_60737182 | 3B | 60737182 | 3.06 × 10−5 | 7.00 | A | 0.12 |
| TNKS | 2.11 × 10−5 | 7.00 | 1.85 | ||||
| SW | S3B_62315382 | 3B | 62315382 | 3.91 × 10−7 | 8.36 | C | 0.15 |
| SL | 1.96 × 10−6 | 8.26 | 0.40 | ||||
| TNKS | 3.91 × 10−7 | 13.36 | 1.85 | ||||
| TSNS | 3.3 × 10−6 | 8.03 | 1.55 | ||||
| SW | S3B_62315407 | 3B | 62315407 | 2.5 × 10−7 | 8.36 | T | 0.15 |
| SL | 2.02 × 10−6 | 8.26 | 0.40 | ||||
| TNKS | 2.5 × 10−6 | 9.23 | 2.23 | ||||
| TSNS | 1.96 × 10−6 | 8.03 | 0.55 | ||||
| SL | S3B_64172577 | 3B | 64172577 | 2.11 × 10−5 | 6.01 | T | 0.12 |
| TNKS | 1.61 × 10−5 | 8.63 | 1.90 | ||||
| TSNS | 2.02 × 10−5 | 6.31 | 0.48 | ||||
| SW | S5D_61792984 | 5D | 61792984 | 1.09 × 10−5 | 6.17 | C | 0.21 |
| SL | 3.72 × 10−7 | 7.96 | 0.65 | ||||
| TNKS | 1.09 × 10−5 | 9.09 | 3.06 | ||||
| TSNS | 3.72 × 10−7 | 6.85 | 0.83 | ||||
| SW | S5D_62479367 | 5D | 62479367 | 2.02 × 10−5 | 6.91 | G | 0.58 |
| TSNS | 2.52 × 10−5 | 5.45 | 0.71 | ||||
| SW | S5D_72377429 | 5D | 72377429 | 1.61 × 10−5 | 5.96 | C | 0.21 |
| SL | 3.3 × 10−6 | 7.77 | 0.64 | ||||
| TNKS | 1.61 × 10−5 | 8.63 | 3.06 | ||||
| TSNS | 2.02 × 10−5 | 6.18 | 0.80 | ||||
| SL | S6B_668517613 | 6B | 668517613 | 1.61 × 10−5 | 5.90 | A | 0.20 |
| TNKS | 2.11 × 10−5 | 7.56 | 3.05 | ||||
| TSNS | 2.09 × 10−5 | 5.75 | 0.77 | ||||
| SL | S7A_610993044 | 7A | 610993044 | 2.11 × 10−5 | 6.00 | G | 0.12 |
| TSNS | 2.67 × 10−5 | 5.87 | 0.46 | ||||
| SL | S7B_729441244 | 7B | 729441244 | 2.11 × 10−5 | 6.62 | A | 0.25 |
| TSNS | 1.87 × 10−5 | 6.60 | 0.94 |
| Grain Yield | Polygenic Marker | Chromosome | Yield-Related Traits |
|---|---|---|---|
| Lincoln | S5D_548379143 | 5D | (SL, SW, TNKS and TSNS) |
| S6D_469537865 | 6D | (SW, TNKS and TSNS) | |
| S7B_329792071 | 7B | (SL, SW, TNKS and TSNS) | |
| S7D_485517060 | 7D | (SL, SW, TNKS and TSNS) | |
| North Platt | S3B_60737182 | 3B | (SL and TNKS) |
| S3B_62315382 | 3B | (SL, SW, TNKS and TSNS) | |
| S3B_62315407 | 3B | (SL, SW, TNKS and TSNS) | |
| S3B_64172577 | 3B | (SL TNKS and TSNS) | |
| S5D_61792984 | 5D | (SL, SW, TNKS and TSNS) | |
| S5D_62479367 | 5D | (SW and TSNS) | |
| S5D_72377429 | 5D | (SL, SW, TNKS and TSNS) | |
| S7B_729441244 | 7B | (SL and TSNS) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eltaher, S.; Sallam, A.; Emara, H.A.; Nower, A.A.; Salem, K.F.M.; Börner, A.; Baenziger, P.S.; Mourad, A.M.I. Genome-Wide Association Mapping Revealed SNP Alleles Associated with Spike Traits in Wheat. Agronomy 2022, 12, 1469. https://doi.org/10.3390/agronomy12061469
Eltaher S, Sallam A, Emara HA, Nower AA, Salem KFM, Börner A, Baenziger PS, Mourad AMI. Genome-Wide Association Mapping Revealed SNP Alleles Associated with Spike Traits in Wheat. Agronomy. 2022; 12(6):1469. https://doi.org/10.3390/agronomy12061469
Chicago/Turabian StyleEltaher, Shamseldeen, Ahmed Sallam, Hamdy A. Emara, Ahmed A. Nower, Khaled F. M. Salem, Andreas Börner, P. Stephen Baenziger, and Amira M. I. Mourad. 2022. "Genome-Wide Association Mapping Revealed SNP Alleles Associated with Spike Traits in Wheat" Agronomy 12, no. 6: 1469. https://doi.org/10.3390/agronomy12061469
APA StyleEltaher, S., Sallam, A., Emara, H. A., Nower, A. A., Salem, K. F. M., Börner, A., Baenziger, P. S., & Mourad, A. M. I. (2022). Genome-Wide Association Mapping Revealed SNP Alleles Associated with Spike Traits in Wheat. Agronomy, 12(6), 1469. https://doi.org/10.3390/agronomy12061469