
Mechanism of Early Flowering in a Landrace Naked Barley eam8.l Mutant

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Lines
2.2. Western Blotting
2.3. RNA Extraction and Expression Analysis
2.4. Statistical Analysis
3. Results
3.1. eam8.l Encodes a Truncated Protein
3.2. Spatial Expression of the EAM8 Gene at Different Developmental Stages
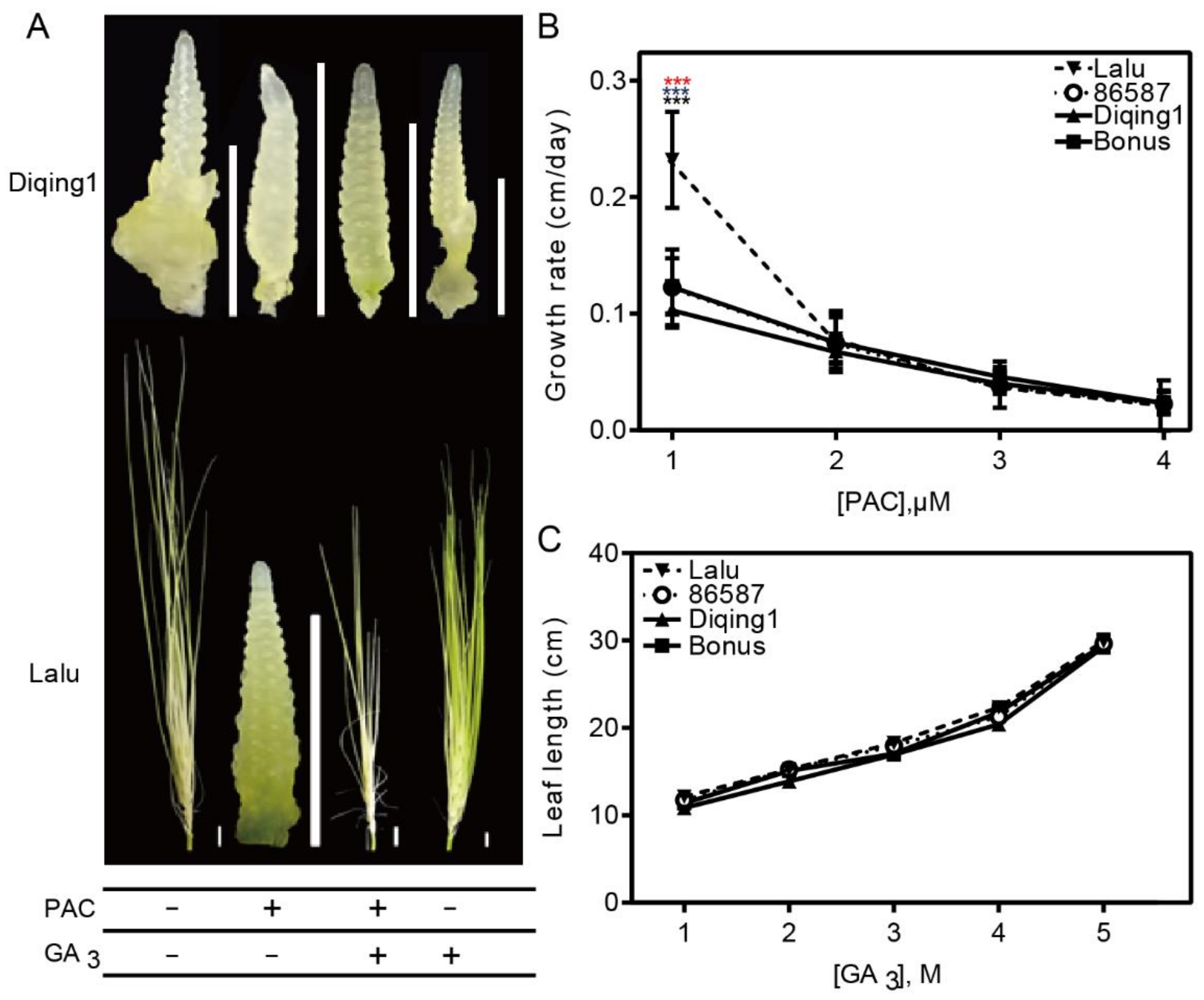
3.3. The Early Flowering Phenotype in Lalu Depends on Increasing GA Production
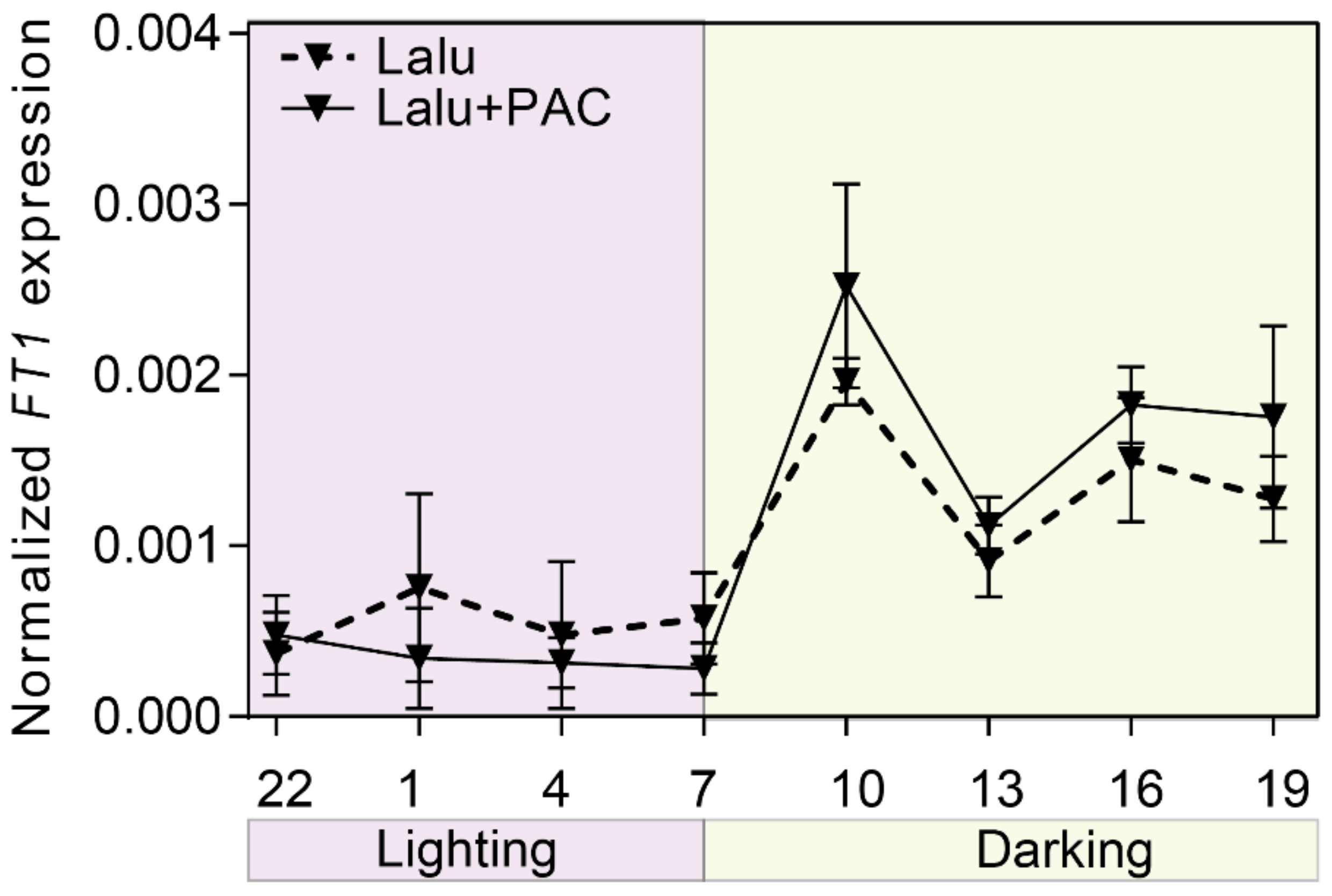
3.4. The GA-Dependent Pathway and FT1 Gene Represent Two Independent Pathways That Promote Flowering in Lalu
3.5. Mutation of the EAM8 Gene (eam8.l) Suppresses GA Synthesis in Lalu
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Simpson, G.G. The autonomous pathway: Epigenetic and post-transcriptional gene regulation in the control of Arabidopsis flowering time. Curr. Opin. Plant Biol. 2004, 7, 570–574. [Google Scholar] [CrossRef] [PubMed]
- Michaels, S.D.; Himelblau, E.; Kim, S.Y.; Schomburg, F.M.; Amasino, R.M. Integration of Flowering Signals in Winter-Annual Arabidopsis. Plant Physiol. 2005, 137, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Kardailsky, I.; Shukla, V.K.; Ahn, J.H.; Dagenais, N.; Christensen, S.K.; Nguyen, J.T.; Chory, J.; Harrison, M.J.; Weigel, D. Activation Tagging of the Floral Inducer FT. Science 1999, 286, 1962–1965. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Xu, D.; Tian, T.; Kong, F.; Lin, K.; Gan, S.; Zhang, H.; Li, G. Molecular and functional dissection of EARLY-FLOWERING 3 (ELF3) and ELF4 in Arabidopsis. Plant Sci. 2021, 303, 110786. [Google Scholar] [CrossRef] [PubMed]
- Zakhrabekova, S.; Gough, S.P.; Braumann, I.; Müller, A.H.; Lundqvist, J.; Ahmann, K.; Dockter, C.; Matyszczak, I.; Kurowska, M.; Druka, A. Induced mutations in circadian clock regulator Mat-a facilitated short-season adaptation and range extension in cultivated barley. Proc. Natl. Acad. Sci. USA 2012, 109, 4326–4331. [Google Scholar] [CrossRef] [PubMed]
- Lundqvist, U. Eighty Years of Scandinavian Barley Mutation Genetics and Breeding; Food and Agriculture Organization of the United Nations: Rome, Italy, 2009; pp. 39–43. [Google Scholar]
- Faure, S.; Turner, A.S.; Gruszka, D.; Christodoulou, V.; Davis, S.J.; von Korff, M.; Laurie, D.A. Mutation at the circadian clock gene 13 EARLY MATURITY 8 adapts domesticated barley (Hordeum vulgare) to short growing seasons. Proc. Natl. Acad. Sci. USA 2012, 109, 8328–8333. [Google Scholar] [CrossRef] [PubMed]
- Pugsley, A.T. The impact of plant physiology on Australian wheat breeding. Euphytica 1983, 32, 743–748. [Google Scholar] [CrossRef]
- Beales, J.; Turner, A.; Griffiths, S.; Snape, J.W.; Laurie, D.A. A Pseudo-Response Regulator is misexpressed in the photoperiod insensitive Ppd-D1a mutant of wheat (Triticum aestivum L.). Theor. Appl. Genet. 2007, 115, 721–733. [Google Scholar] [CrossRef]
- Boden, S.A.; Weiss, D.; Ross, J.J.; Davies, N.W.; Trevaskis, B.; Chandler, P.M.; Swain, S.M. EARLY FLOWERING3 Regulates Flowering in Spring Barley by Mediating Gibberellin Production and FLOWERING LOCUS T Expression. Plant Cell 2014, 26, 1557–1569. [Google Scholar] [CrossRef]
- Xia, T.; Zhang, L.; Xu, J.; Wang, L.; Liu, B.; Hao, M.; Chang, X.; Zhang, T.; Li, S.; Zhang, H. The alternative splicing of EAM8 contributes to early flowering and short-season adaptation in a landrace barley from the Qinghai-Tibetan Plateau. Theor. Appl. Genet. 2017, 130, 757–766. [Google Scholar] [CrossRef]
- Wolf, F.T.; Haber, A.H.; Wolf, A.H.H.F.T. Chlorophyll Content of Gibberellin-Treated Wheat Seedlings. Nature 1960, 186, 217–218. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Raman, A.S.; Ream, J.E.; Fujiwara, H.; Cerny, R.E.; Brown, S.M. Overexpression of 20-Oxidase Confers a Gibberellin-Overproduction Phenotype in Arabidopsis. Plant Physiol. 1998, 118, 773–781. [Google Scholar] [CrossRef] [PubMed]
- Hauvermale, A.L.; Ariizumi, T.; Steber, C.M. Gibberellin Signaling: A Theme and Variations on DELLA Repression. Plant Physiol. 2012, 160, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Davies, L.J.; Rappaport, L. Metabolism of Tritiated Gibberellins in d-5 Dward Maize. Plant Physiol. 1975, 56, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.L.; Li, L.; Wu, K.; Peeters, A.J.; Gage, A.D.; Zeevaart, A.J. The GA5 locus of Arabidopsis thaliana encodes a multifunctional gibberellin 20-oxidase: Molecular cloning and functional expression. Proc. Natl. Acad. Sci. USA 1995, 92, 6640–6644. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.; Beales, J.; Faure, S.; Dunford, R.P.; Laurie, D.A. The Pseudo-Response Regulator Ppd-H1 Provides Adaptation to Photoperiod in Barley. Science 2005, 310, 1031–1034. [Google Scholar] [CrossRef] [PubMed]
- Lang, A. The Effect of Gibberellin upon Flower Formation. Proc. Natl. Acad. Sci. USA 1957, 43, 709–717. [Google Scholar] [CrossRef]
- Chailakhyan, M.K. Hormonale faktoren des pflanzenblühens. Biol. Zent. Bl. 1958, 77, 641–662. [Google Scholar]
- Bernier, G.; Havelange, A.; Houssa, C.; Petitjean, A.; Lejeune, P. Physiological Signals That Induce Flowering. Plant Cell 1993, 5, 1147–1155. [Google Scholar] [CrossRef]
- Blázquez, M.A.; Trénor, M.; Weigel, D. Independent Control of Gibberellin Biosynthesis and Flowering Time by the Circadian Clock in Arabidopsis. Plant Physiol. 2002, 130, 1770–1775. [Google Scholar] [CrossRef]
- Yan, L.; Loukoianov, A.; Tranquilli, G.; Helguera, M.; Fahima, T.; Dubcovsky, J. Positional cloning of the wheat vernalization gene VRN1. Proc. Natl. Acad. Sci. USA 2003, 100, 6263–6268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixon, L.E.; Knox, K.; Kozma-Bognar, L.; Southern, M.M.; Pokhilko, A.; Millar, A.J. Temporal Repression of Core Circadian Genes Is Mediated through EARLY FLOWERING 3 in Arabidopsis. Curr. Biol. 2011, 21, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Fowler, S.; Lee, K.; Onouchi, H.; Samach, A.; Richardson, K.; Morris, B.; Coupland, G.; Putterill, J. GIGANTEA: A circadian clock-controlled gene that regulates photoperiodic flowering in Arabidopsis and encodes a protein with several possible membrane-spanning domains. EMBO J. 1999, 18, 4679–4688. [Google Scholar] [CrossRef] [PubMed]
- King, R.W.; Moritz, T.; Evans, L.T.; Martin, J.; Andersen, C.H.; Blundell, C.; Kardailsky, I.; Chandler, P.M. Regulation of Flowering in the Long-Day Grass Lolium temulentum by Gibberellins and the FLOWERING LOCUS T Gene. Plant Physiol. 2006, 141, 498–507. [Google Scholar] [CrossRef]
- Hisamatsu, T.; King, R.W. The nature of floral signals in Arabidopsis. II. Roles for FLOWERING LOCUS T (FT) and gibberellin. J. Exp. Bot. 2008, 59, 3821–3829. [Google Scholar] [CrossRef]
- Porri, A.; Torti, S.; Romera-Branchat, M.; Coupland, G. Spatially distinct regulatory roles for gibberellins in the promotion of flowering of Arabidopsis under long photoperiods. Development 2012, 139, 2198–2209. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mao, C.; Xu, J.; Wang, H.; Chen, T.; Bian, H.; You, E.; Shen, Y.; Wang, L. Mechanism of Early Flowering in a Landrace Naked Barley eam8.l Mutant. Agronomy 2022, 12, 2137. https://doi.org/10.3390/agronomy12092137
Mao C, Xu J, Wang H, Chen T, Bian H, You E, Shen Y, Wang L. Mechanism of Early Flowering in a Landrace Naked Barley eam8.l Mutant. Agronomy. 2022; 12(9):2137. https://doi.org/10.3390/agronomy12092137
Chicago/Turabian StyleMao, Chengzhi, Jinqing Xu, Handong Wang, Tongrui Chen, Haiyan Bian, En You, Yuhu Shen, and Lei Wang. 2022. "Mechanism of Early Flowering in a Landrace Naked Barley eam8.l Mutant" Agronomy 12, no. 9: 2137. https://doi.org/10.3390/agronomy12092137
APA StyleMao, C., Xu, J., Wang, H., Chen, T., Bian, H., You, E., Shen, Y., & Wang, L. (2022). Mechanism of Early Flowering in a Landrace Naked Barley eam8.l Mutant. Agronomy, 12(9), 2137. https://doi.org/10.3390/agronomy12092137