
The Increase in the Karstification–Photosynthesis Coupled Carbon Sink and Its Implication for Carbon Neutrality



Abstract
:1. Introduction
2. Uncoupled Karstification Limits Photosynthetic Carbon Sequestration
2.1. Karst Drought
2.2. High pH, and High Bicarbonate and Calcium Contents
2.3. Deficiency of Available Nitrogen, Phosphorus, and Nutrient Elements
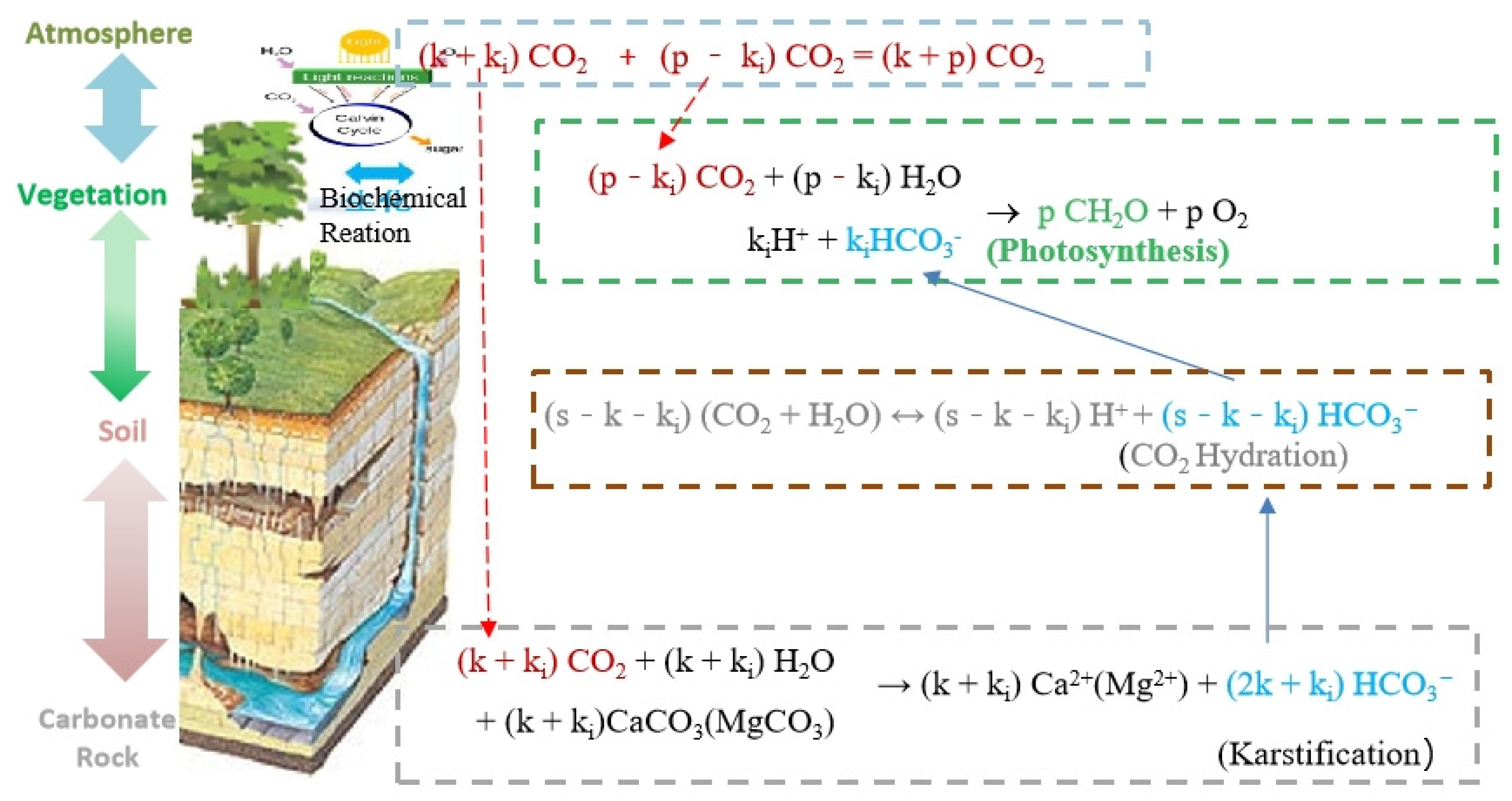
3. Karstification–Photosynthesis Coupling Increases Karst Carbon Sinks
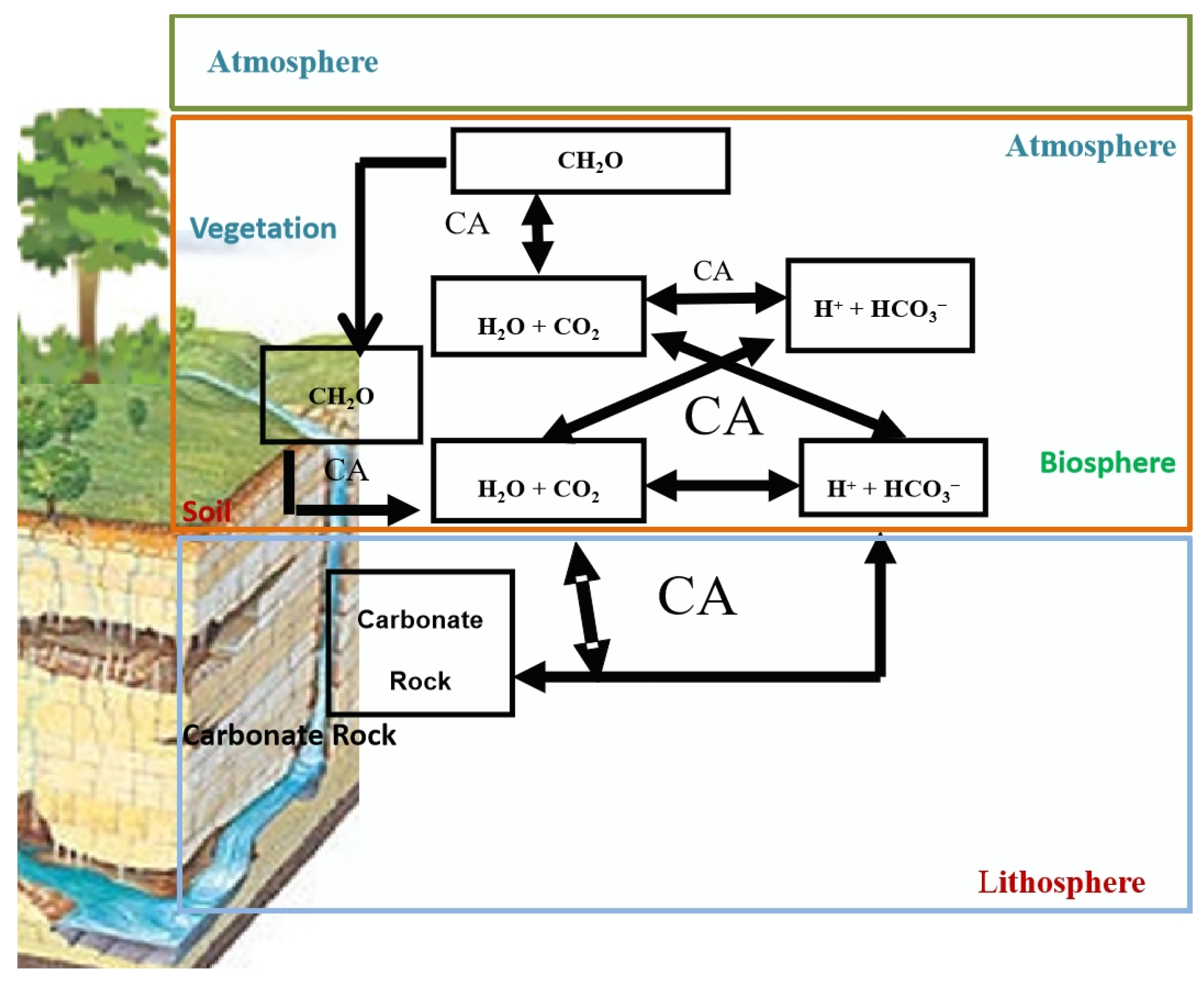
4. Carbonic Anhydrase, a Key Pivot of Karstification–Photosynthesis Coupling
4.1. Carbonic Anhydrase Controls the Transport of Water and Inorganic Carbon at the Carbonate Rocks–Soil–Atmosphere Interface by Catalyzing Carbonate Dissolution and Carbon Dioxide Hydration
4.2. Carbonic Anhydrase Promotes Inorganic Carbon Assimilation in Plants
5. Carbon Sequestration and the Enhancement Effect of Karst-Adaptable Plants during Karstification–Photosynthesis Coupling
6. Conclusions and Outlook
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC. IPCC Sixth Assessment Report. Available online: https://www.ipcc.ch/report/ar6/wg1/, (accessed on 13 July 2022).
- NOAA. Global Monitoring Laboratory. Available online: https://gml.noaa.gov/ccgg/trends/mlo.html, (accessed on 13 July 2022).
- Jia, Z.; Li, Z. Carbonate Sedimentary Facies and Sedimentary Environment; China University of Geosciences Press: Wuhan, China, 1989; pp. 1–6. [Google Scholar]
- Wang, S. The most serious ecological and geological environmental problem in southwest China-Karst rock desertification. Bull Miner. Pet. Geochem 2003, 22, 120–126. [Google Scholar]
- Jiang, Z.; Lian, Y.; Qin, X. Rocky desertification in Southwest China: Impacts, causes, and restoration. Earth Sci. Rev. 2014, 132, 1–12. [Google Scholar] [CrossRef]
- Xiong, P.; Yuan, D.; Xie, S. Progress of research on rocky desertification in South China Karst Mountain. Carsol Sin 2010, 29, 355–356. [Google Scholar]
- Oelkers, E.H.; Declercq, J.; Saldi, G.D.; Gislason, S.R.; Schott, J. Olivine dissolution rates: A critical review. Chem. Geol. 2018, 500, 1–19. [Google Scholar] [CrossRef]
- Gaillardet, J.; Galy, A. Himalaya-Carbon Sink or Source. Science 2008, 320, 1727–1728. [Google Scholar] [CrossRef] [PubMed]
- Yuan, D. Progress in the study on “Karst process and carbon cycle”. Adv. Earth Sci. 1999, 14, 425–432. [Google Scholar]
- Larson, C. An unsung carbon sink. Science 2011, 334, 886–887. [Google Scholar] [CrossRef]
- Liu, Z. “Method of maximum potential dissolution” to calculate the intensity of karst process and the relevent carbon sink: With discussions on methods of solute load carbonate-rock-tablet test. Carsol. Sin. 2011, 30, 379–382. [Google Scholar]
- Liu, Z.H. New progress and prospects in the study of rock-weathering-related carbon sinks. Chin. Sci. Bull. 2012, 57, 95–102. [Google Scholar] [CrossRef]
- Chen, C.Y.; Liu, Z.H. The role of biological carbon pump in the carbon sink and water environment improvement in karst surface aquatic ecosystems. Chin. Sci. Bull. 2017, 62, 3440–3450. [Google Scholar] [CrossRef]
- Berner, R.A. Atmospheric carbon dioxide levels over Phanerozoic time. Science 1990, 249, 1382–1386. [Google Scholar] [CrossRef] [PubMed]
- Berner, R.A.; Kothavala, Z. GEOCARB III: A revised model of atmospheric CO2 over Phanerozoic time. Am. J. Sci. 2001, 301, 182–204. [Google Scholar] [CrossRef]
- Curl, R.L. Carbon shifted but not sequestered. Science 2012, 335, 655. [Google Scholar] [CrossRef]
- Yuan, D. Rock desertification in the subtropical karst of south China. Z. Geomorph. (Suppl.) 1997, 108, 81–90. [Google Scholar]
- Cao, J.; Yuan, D.; Pan, G. Sme soil features in karst ecosystem. Adv. Earth Sci. 2003, 18, 37–44. [Google Scholar]
- Legrand, H.E.; Stringfield, V.T. Karst hydrology:A review. J. Hydrol. 1973, 20, 97–120. [Google Scholar] [CrossRef]
- Qin, X. Characteristics and comprehensive treatment strategy of karst drought in Central Guangxi. J. Guilin Univ. Technol. 2005, 25, 278–283. [Google Scholar]
- Yuan, D. Carbon cycle in Earth system and its effects on environment and resources. Quatern. Sci. 2001, 21, 223–232. [Google Scholar]
- Guo, F.; Jiang, G.; Yuan, D. Major ions in typical subterranean rivers and their anthropogenic impacts in southwest karst areas, China. Environ. Geol. 2007, 53, 533–541. [Google Scholar] [CrossRef]
- Song, T.; Peng, W.; Du, H.; Wang, K.; Zeng, F. Occurrence, spatial-temporal dynamics and regulation strategies of karst rocky desertification in southwest China. Chin. J. Ecol. 2014, 34, 5328–5341. [Google Scholar]
- Liu, L.; Zeng, F.; Song, T.; Peng, W.; Wang, K.; Qin, W.; Tan, W. Spatial heterogeneity of soil nutrients in Karst area’s Mulun National Nature Reserve. Chin. J. Appl. Ecol. 2010, 21, 1667–1673. [Google Scholar]
- Dong, L.; He, T.; Liu, Y.; Shu, Y.; Luo, H.; Liu, F. Changes of soil physical- chemical properties derived from different parent materials/rocks in karst mountain. Chin. J. Soil Sci. 2008, 39, 471–474. [Google Scholar]
- Ji, F.; Li, N.; Deng, X. Calcium contents and high calcium adaptation of plants in karst area of China. Chin. J. Plant Ecol. 2009, 33, 926–935. [Google Scholar]
- Xiong, D.; Cai, H.; Zhang, R.; Li, C.; QIan, X. Distribution of soil medium and micro-elements in Fujian tobacco-growing soils. Chin. J. Eco Agric. 2007, 15, 36–38. [Google Scholar]
- Li, Z.; Zou, B.; Cao, Y.; Ren, H.; Liu, J. nutrient characteristics of soils in typical degraded hilly land in Southern China. Acta. Ecol. Sin. 2003, 23, 1648–1656. [Google Scholar]
- Paramasivam, S.; Alva, A.K. Nitrogen recovery from controlled-release fertilizers under intermittent leaching and dry cycles. Soil Sci. 1997, 162, 447–453. [Google Scholar] [CrossRef]
- Wei, X.; Shao, M. The distribution of soil nutrients on sloping land in the gully regiion watersheds of the Loess Plateau. Acta. Ecol. Sin. 2007, 27, 603–613. [Google Scholar]
- Liu, C. Biogeochemical Processes and Cycling of Nutrients in the Earth’s Surface: Cycling of Nutrients in Soil-Plant Systems of Karstic Environments, Southwest China; Science Press of China: Beijing, China, 2009; pp. 308–353, 401–443. [Google Scholar]
- National Soil Census Office of China. Soil in China; China Agriculture Press: Beijing, China, 1998; pp. 843–984. [Google Scholar]
- Jiang, D.; Cheng, J. Changes of pH and available pool of B, Cu, Zn, Mn in surface rice soils in Yixing, Jiangsu province over the last decade. J. Nanjing Agric. Univ. 1997, 20, 111–113. [Google Scholar]
- Wu, Y.Y.; Xing, D.K. Effect of bicarbonate treatment on photosynthetic assimilation of inorganic carbon in two plant species of Moraceae. Photosynth. 2012, 50, 587–594. [Google Scholar] [CrossRef]
- Wu, Y.; Li, H.; Xie, T. Biogeochemical Action of Microalgal Carbonic Anhydrase; Science Press of China: Beijing, China, 2015; pp. 75–110. [Google Scholar]
- Hang, H.T.; Wu, Y.Y. Quantification of photosynthetic inorganic carbon utilisation via a bidirectional stable carbon isotope tracer. Acta. Geochim. 2016, 35, 130–137. [Google Scholar] [CrossRef]
- Rao, S.; Wu, Y.Y. Root-derived bicarbonate assimilation in response to variable water deficit in Camptotheca acuminate seedlings. Photosyn. Res. 2017, 134, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Wu, Y.; Xing, D.; Hang, H.; Xie, X.; Yang, X.; Zhang, K.; Rao, S. Biomass production of three biofuel energy plants’ use of a new carbon resource by carbonic anhydrase in simulated karst soils: Mechanism and capacity. Energies 2017, 10, 1370. [Google Scholar] [CrossRef]
- Xie, T.; Wu, Y. The biokarst system and its carbon sinks in response to pH changes: A simulation experiment with microalgae. Geochem. Geophys. Geosyst. 2017, 18, 827–843. [Google Scholar] [CrossRef]
- Wu, Y.; Xing, D.; Hang, H.; Zhao, K. Principles and Techniques of Determination on Plants’ Adaptation to Karst Environment; Science Press of China: Beijing, China, 2018; pp. 1–188. [Google Scholar]
- Fang, L.; Wu, Y.Y. Bicarbonate uptake experiment show potential karst carbon sinks transformation into carbon sequestration by terrestrial higher plants. J. Plant Interact. 2022, 17, 419–426. [Google Scholar] [CrossRef]
- Wu, Y. Combined effect of bicarbonate and water in photosynthetic oxygen evolution and its implication for carbon neutrality. Sci. Bull. 2022. submitted and revised. [Google Scholar]
- He, S.; Yu, S.; Pu, J.; Yuan, Y.; Zhang, C. Dynamics in Riverine Inorganic and Organic Carbon Based on Carbonate Weathering Coupled with Aquatic Photosynthesis in a Karst Catchment, Southwest China. Water Res. 2021, 189, 116658. [Google Scholar]
- Yang, R.; Sun, H.; Chen, B.; Yang, M.; Zeng, Q.; Zeng, C.; Huang, J.; Luo, H.; Lin, D. Temporal variations in riverine hydrochemistry and estimation of the carbon sink produced by coupled carbonate weathering with aquatic photosynthesis on land: An example from the Xijiang River, a large subtropical karst-dominated river in China. Environ. Sci. Pollut. Res. 2020, 27, 13142–13154. [Google Scholar] [CrossRef]
- Cao, J.H.; Wu, X.; Huang, F.; Hu, B.; Groves, C.; Yang, H.; Zhang, C.L. Global significance of the carbon cycle in the karst dynamic system: Evidence from geological and ecological processes. China Geol. 2018, 1, 17–27. [Google Scholar] [CrossRef]
- Berner, R.A. The carbon cycle and carbon dioxide over Phanerozoic time: The role of land plants. Philos. Trans. R. Soc. B 1998, 353, 75–82. [Google Scholar] [CrossRef]
- Moulton, K.L.; Berner, R.A. Quantification of the effect of plants on weathering: Studies in Iceland. Geology 1998, 26, 895–898. [Google Scholar] [CrossRef]
- Knoll, A.H.; Nowak, M.A. The timetable of evolution. Sci. Adv. 2017, 3, e1603076. [Google Scholar] [CrossRef] [PubMed]
- Hart, M.H. The evolution of the atmosphere of the Earth. Icarus 1978, 33, 23–39. [Google Scholar] [CrossRef]
- Des Marais, D.J. When did photosynthesis emerge on Earth? Science 2000, 289, 1703–1705. [Google Scholar] [CrossRef]
- Dismukes, G.C.; Klimov, V.V.; Baranov, S.V.; Kozlov, Y.N.; DasGupta, J.; Tyryshkin, A. The origin of atmospheric oxygen on Earth: The innovation of oxygenic photosynthesis. Proc. Natl. Acad. Sci. USA 2001, 98, 2170–2175. [Google Scholar] [CrossRef] [PubMed]
- Pagani, M.; Caldeira, K.; Berner, R.A.; Beerling, D.J. The role of terrestrial plants in limiting atmospheric CO2 decline over the past 24 million years. Nature 2009, 460, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Kasting, J.F. Earth’s early atmosphere. Science 1993, 259, 920–926. [Google Scholar] [CrossRef]
- Halevy, I.; Bachan, A. The geologic history of seawater pH. Science 2017, 355, 1069–1071. [Google Scholar] [CrossRef]
- Teng, F.Z.; Ma, L. Deciphering isotope signatures of Earth Surface and Critical Zone processes. Chem. Geol. 2016, 445, 1–3. [Google Scholar] [CrossRef]
- Schwender, J.; Goffman, F.; Ohlrogge, J.B.; Shachar-Hill, Y. Rubisco without the Calvin cycle improves the carbon efficiency of developing green seeds. Nature 2004, 432, 779–782. [Google Scholar] [CrossRef]
- Tcherkez, G.; Mahe, A.; Gauthier, P.; Mauve, C.; Gout, E.; Bligny, R.; Cornic, G.; Hodges, M. In folio respiratory fluxomics revealed by 13C isotopic labeling and H/D isotope effects highlight the noncyclic nature of the tricarboxylic acid “Cycle” in illuminated leaves. Plant Physiol. 2009, 151, 620–630. [Google Scholar] [CrossRef]
- Lin, G. Stable isotope ecology: A new branch of ecology resulted from technology advances. Chin. J. Plant Ecol. 2010, 34, 119–122. [Google Scholar]
- Nichols, J.; Booth, R.K.; Jackson, S.T.; Pendall, E.G.; Huang, Y.S. Differential hydrogen isotopic ratios of Sphagnum and vascular plant biomarkers in ombrotrophic peatlands as a quantitative proxy for precipitation-evaporation balance. Geochim. Cosmochim. Acta 2010, 74, 1407–1416. [Google Scholar] [CrossRef]
- Broeckx, L.S.; Fichot, R.; Verlinden, M.S.; Ceulemans, R. Seasonal variations in photosynthesis, intrinsic water-use efficiency and stable isotope composition of poplar leaves in a short-rotation plantation. Tree Physiol. 2014, 34, 701–715. [Google Scholar] [CrossRef] [PubMed]
- Xie, T.; Wu, Y. The role of microalgae and their carbonic anhydrase on the biological dissolution of limestone. Environ. Earth Sci. 2014, 71, 5231–5239. [Google Scholar] [CrossRef]
- Hang, H. Expression Encoding Carbonic Anhydrase Gene and Inorganic Carbon Utilization of the Two Plants Species under Karst Environment. Ph.D. Dissertation, The University of Chinese Academy of Sciences, Beijing, China, 2015. [Google Scholar]
- Yan, W.; Zeng, C.; Xiao, S.; Lan, J.; Dai, L.; Tai, Z.; He, J.; He, C.; Di, Y. Dissolution rate and karst carbon sink of different land use in typical dolomite watershed with humid subtropical weather. Earth Environ. 2021, 49, 529–538. [Google Scholar]
- Zeng, C.; Zhao, M.; Yang, R.; Liu, Z. Comparison of karst processes-related carbon sink intensity calculated by carbonate rock tablet test and solute load method: A case study in the Chenqi karst spring system. Hydrogeo Eng. Geol. 2014, 41, 106–111. [Google Scholar]
- Lan, J.C.; Fu, W.L.; Peng, J.T.; Zhou, X.P.; Xiao, S.Z.; Yuan, B. Dissolution rate under soil in karst areas and the influencing factors of different land use patterns. Acta Ecol. Sin. 2013, 33, 3205–3212. [Google Scholar]
- Zhang, C. Carbonate rock dissolution rates in different land uses and their carbon sink effect. Chin. Sci. Bull. 2011, 56, 3759–3765. [Google Scholar] [CrossRef]
- Zeng, C.; Liu, Z.; Zhao, M.; Yang, R. Hydrologically-driven variations in the karst-related carbon sink fluxes: Insights from high-resolution monitoring of three karst catchments in Southwest China. J. Hydrol. 2016, 533, 74–90. [Google Scholar] [CrossRef]
- Zeng, C.; He, C.; Xiao, S.; Liu, Z.; Chen, W.; He, J. Karst related inorganic carbon sink flux in a typical humid subtropical dolomite basin. Earth Sci. Front. 2022, 29, 179–188. [Google Scholar]
- Li, S.; Liu, C.; Li, J.; Lang, Y.; Ding, H.; Li, L. Geochemistry of dissolved inorganic carbon and carbonate weathering in a small typical karstic catchment of Southwest China: Isotopic and chemical constraints. Chem. Geol. 2010, 277, 301–309. [Google Scholar] [CrossRef]
- Yu, W.; Dong, D.; Ni, J. Comparison of biomass and net primary productivity of Karst and non-karst forests in mountainous area, Southwestern China. J. Subtrop. Resour. Environ. 2010, 5, 25–30. [Google Scholar]
- Wu, Y.; Rao, S.; Wu, Y.; Fang, L.; Su, Y.; Li, H.; Wang, R.; Wang, S.; Liu, C. A Method for Obtaining the Share of Bicarbonate Utilization by Plants in Natural Habitats; ZL2018103132903, Beijing; CN108319820B; State Intellectual Property Office of China: Beijing, China, 2018. [Google Scholar]
- Xia, H. Biomass and net primary production in different successional stages of karst vegetation in Maolan, SW China, Guizhou. For. Sci. Technol. 2010, 38, 1–7. [Google Scholar]
- Veitch, F.P.; Blankenship, L.C. Carbonic anhydrase in bacteria. Nature 1963, 197, 76–77. [Google Scholar] [CrossRef] [PubMed]
- Badger, M.R.; Price, G.D. The role of carbonic anhydrase in photosynthesis. Annu. Rev. Plant Biol. 1994, 45, 369–392. [Google Scholar] [CrossRef]
- Smith, K.S.; Ferry, J.G. Prokaryotic carbonic anhydrases. FEMS Microbiol. Rev. 2000, 24, 335–366. [Google Scholar] [CrossRef] [PubMed]
- Gilmour, K.M.; Perry, S.F. Carbonic anhydrase and acid-base regulation in fish. J. Exp. Biol. 2009, 212, 1647–1661. [Google Scholar] [CrossRef] [PubMed]
- Price, G.D.; Long, B.M.; Förster, B. DABs accumulate bicarbonate. Nat. Microbiol. 2019, 4, 2029–2030. [Google Scholar] [CrossRef]
- Ivanov, B.N.; Ignatova, L.K.; Romanova, A.K. Diversity in forms and functions of carbonic anhydrase in terrestrial higher plants. Russ. J. Plant Physl. 2007, 54, 143–162. [Google Scholar] [CrossRef]
- Sun, W.; Wu, Y.; Wen, X.; Xiong, S.; He, H.; Wang, Y.; Lu, G. Different mechanisms of photosynthetic response to drought stress in tomato and violet orychophragmus. Photosynthetica 2016, 54, 226–233. [Google Scholar] [CrossRef]
- DiMario, R.J.; Machingura, M.C.; Waldrop, G.L.; Moroney, J.V. The many types of carbonic anhydrases in photosynthetic organisms. Plant Sci. 2018, 268, 11–17. [Google Scholar] [CrossRef]
- Hang, H.; Wu, Y. Effect of bicarbonate stress on carbonic anhydrase gene expressions from Orychophragmus violaceus and Brassica juncea seedlings. Pol. J. Environ. Stud. 2019, 28, 1135–1143. [Google Scholar] [CrossRef]
- Liu, Z.; Dreybrod, W. Dissolution kinetics of calcium carbonate minerals in H2O--CO2 solutions in turbulent flow: The role of the diffusion boundary layer and the slow reaction H2O + CO2 → H+ + HCO3−. Geochim. Cosmochim. Acta 1997, 61, 2879–2889. [Google Scholar] [CrossRef]
- Li, W.; Yu, L.J.; Wu, Y.; Jia, L.P.; Yuan, D.X. Enhancement of Ca2+ release from limestone by microbial extracellular carbonic anhydrase. Bioresour. Technol. 2007, 98, 950–953. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z. Role of carbonic anhydrase as an activator in carbonate rock dissolution and its implication for atmospheric CO2 sink. Acta Geol. Sin. (Engl. Ed.) 2001, 75, 275–278. [Google Scholar]
- Papamichael, E.M.; Economou, E.D.; Vaimakis, T.C. Dissolution of the carbonate minerals of phosphate ores: Catalysis by carbonic anhydrase II, from bovine erythrocytes, in acid solutions. J. Colloid Interf. Sci. 2002, 251, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Seibt, U.; Wingate, L.; Lloyd, J.; Berry, J.A. Diurnally variable Δ18O signatures of soil CO2 fluxes indicate carbonic anhydrase activity in a forest soil. J. Geophy. Res. Biogeo. 2006, 111, G04005. [Google Scholar]
- Wingate, L.; Seibt, U.; Maseyk, K.; Ogee, J.; Almeida, P.; Yakir, D.; Pereira, J.; Mencuccini, M. Evaporation and carbonic anhydrase activity recorded in oxygen isotope signatures of net CO2 fluxes from a Mediterranean soil. Glob. Change Biol. 2008, 14, 2178–2193. [Google Scholar] [CrossRef]
- Von Sperber, C.; Weiler, M.; Brüggemann, N. The effect of soil moisture, soil particle size, litter layer and carbonic anhydrase on the oxygen isotopic composition of soil-released CO2. Eur. J. Soil Sci. 2015, 66, 566–576. [Google Scholar] [CrossRef]
- Hu, H.; Boisson-Dernier, A.; Israelsson-Nordström, M.; Böhmer, M.; Xue, S.; Ries, A.; Godoski, J.; Kuhn, J.M.; Schroeder, J.I. Carbonic anhydrases are upstream regulators of CO2-controlled stomatal movements in guard cells. Nat. Cell Biol. 2010, 12, 87–93. [Google Scholar] [CrossRef]
- Hu, H.; Rappel, W.J.; Occhipinti, R.; Ries, A.; Böhmer, M.; You, L.; Xiao, C.; Engineer, C.B.; Boron, W.F.; Schroeder, J.I. Distinct cellular locations of carbonic anhydrases mediate carbon dioxide control of stomatal movements. Plant Physiol. 2015, 169, 1168–1178. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.Y.; Liu, C.Q.; Li, P.P.; Wang, J.Z.; Xing, D.; Wang, B.L. Photosynthetic characteristics involved in adaptability to Karst soil and alien invasion of paper mulberry (Broussonetia papyrifera (L.) Vent.) in comparison with mulberry (Morus alba L.). Photosynthetica 2009, 47, 155–160. [Google Scholar] [CrossRef]
- Nathan, V.K.; Ammini, P. Carbon dioxide sequestering ability of bacterial carbonic anhydrase in a mangrove soil microcosm and its bio-mineralization properties. Water Air Soil Pollut. 2019, 230, 192. [Google Scholar] [CrossRef]
- Wu, Y.; Liu, C.; Wang, S. Adaptability to Karst of Orychophragmus violaceus; Guizhou Science and Technology Press: Guiyang, China, 2014; pp. 1–153. [Google Scholar]
- Xia, A.; Wu, Y. Joint interactions of carbon and nitrogen metabolism dominated by bicarbonate and nitrogen in Orychophragmus violaceus and Brassica napus under simulated karst habitats. BMC Plant Biol. 2022, 22, 264. [Google Scholar] [CrossRef]
- Wu, Y. Strategies to increase carbon fixation and sequestration by Karst-adaptable plants. Carsol. Sin. 2011, 30, 461–465. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Land Use | Average Dissolution Rate mg cm−2 y−1 | Karst Carbon Sink Intensity tCO2 km−2 y−1 | Depth of the Underlying Carbonate-Rock-Tablet cm | Reference |
|---|---|---|---|---|
| Woodland | 1.01 | 4.69 | 10 | [63] |
| Dry land | 4.25 | 19.72 | 60 | |
| Paddy land | 0.14 | 0.65 | 46 | |
| Shrub | 0.61 | 2.83 | 50 | [64] |
| Dry land | 5.27 | 24.45 | 50 | |
| Paddy land | 3.05 | 14.15 | 50 | |
| Woodland | 7.61 | 35.31 | 50 | [65] |
| Grassland | 8.89 | 41.25 | 50 | |
| Vegetable-planted land | 6.21 | 28.81 | 50 | |
| Tilled land | 11.0 | 51.04 | 20 | [66] |
| Tilled land | 14.9 | 69.14 | 50 | |
| Shrub | 0.5 | 2.32 | 20 | |
| Shrub | 2.6 | 12.06 | 50 | |
| Woodland | 68.7 | 318.77 | 20 | |
| Woodland | 18.7 | 86.77 | 50 | |
| Orchard | 87.7 | 406.93 | 20 | |
| Orchard | 120.1 | 557.26 | 50 |
| Catchment Name (Location) | CSFs | Reference | Catchment Name (Location) | CSFs | Reference |
|---|---|---|---|---|---|
| Banzhai (Libo) | 28.84 ± 3.04 | [67] | Huangzhou River (Shibing) | 36.43 | [68] |
| Huanghou (Libo) | 32.81 ± 4.70 | Chenqi (Puding) | 55.07 | [64] | |
| Houzhai (Puding) | 39.13 ± 7.56 | Houzhai (Puding) | 25.70 | [69] |
| Succession Stage | Arbor Layer | Shrub Layer | Herbal Layer | Coarse, Medium Root | Fine Roots | Total | Carbon Sinks 1 |
|---|---|---|---|---|---|---|---|
| Climax community of evergreen-deciduous broadleaved mixed forests | 7.01 | 0.25 | 0.04 | 1.51 | 4.72 | 13.58 | 2240.70 |
| Sub-climax community of evergreen-deciduous broadleaved mixed forests | 5.09 | 0.32 | 0.09 | 0.71 | 2.52 | 8.73 | 1440.45 |
| Scrub–shrub community | 1.39 | 0.79 | 0.06 | 0.57 | 3.27 | 6.08 | 1003.20 |
| Herb–scrub community | 1.03 | 0.60 | 0.85 | 2.10 | 4.58 | 755.70 | |
| Herb community | 1.19 | 0.12 | 1.42 | 2.73 | 450.45 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.; Wu, Y. The Increase in the Karstification–Photosynthesis Coupled Carbon Sink and Its Implication for Carbon Neutrality. Agronomy 2022, 12, 2147. https://doi.org/10.3390/agronomy12092147
Wu Y, Wu Y. The Increase in the Karstification–Photosynthesis Coupled Carbon Sink and Its Implication for Carbon Neutrality. Agronomy. 2022; 12(9):2147. https://doi.org/10.3390/agronomy12092147
Chicago/Turabian StyleWu, Yanyou, and Yansheng Wu. 2022. "The Increase in the Karstification–Photosynthesis Coupled Carbon Sink and Its Implication for Carbon Neutrality" Agronomy 12, no. 9: 2147. https://doi.org/10.3390/agronomy12092147
APA StyleWu, Y., & Wu, Y. (2022). The Increase in the Karstification–Photosynthesis Coupled Carbon Sink and Its Implication for Carbon Neutrality. Agronomy, 12(9), 2147. https://doi.org/10.3390/agronomy12092147