


Thymol Induces Cell Death of Fusarium oxysporum f. sp. niveum via Triggering Superoxide Radical Accumulation and Oxidative Injury In Vitro


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Culturing Strains
2.2. Testing the Sensitivity of FON to Thymol In Vitro
2.3. Microscopic Observation of Mycelia
2.4. Fluorescent Detection of Superoxide Radical, Lipid Peroxidation, and Cell Death in Mycelia and Conidia
2.5. Determining Electrical Conductivity of Myclia
2.6. Determining TBARS (Thiobarbituric Acid Reactive Substances) Content in Mycelia
2.7. Determining Glycerol Content in Mycelia
2.8. Assays of Antioxidative Enzyme Activities in Mycelia
2.9. Statistical Analysis
3. Results
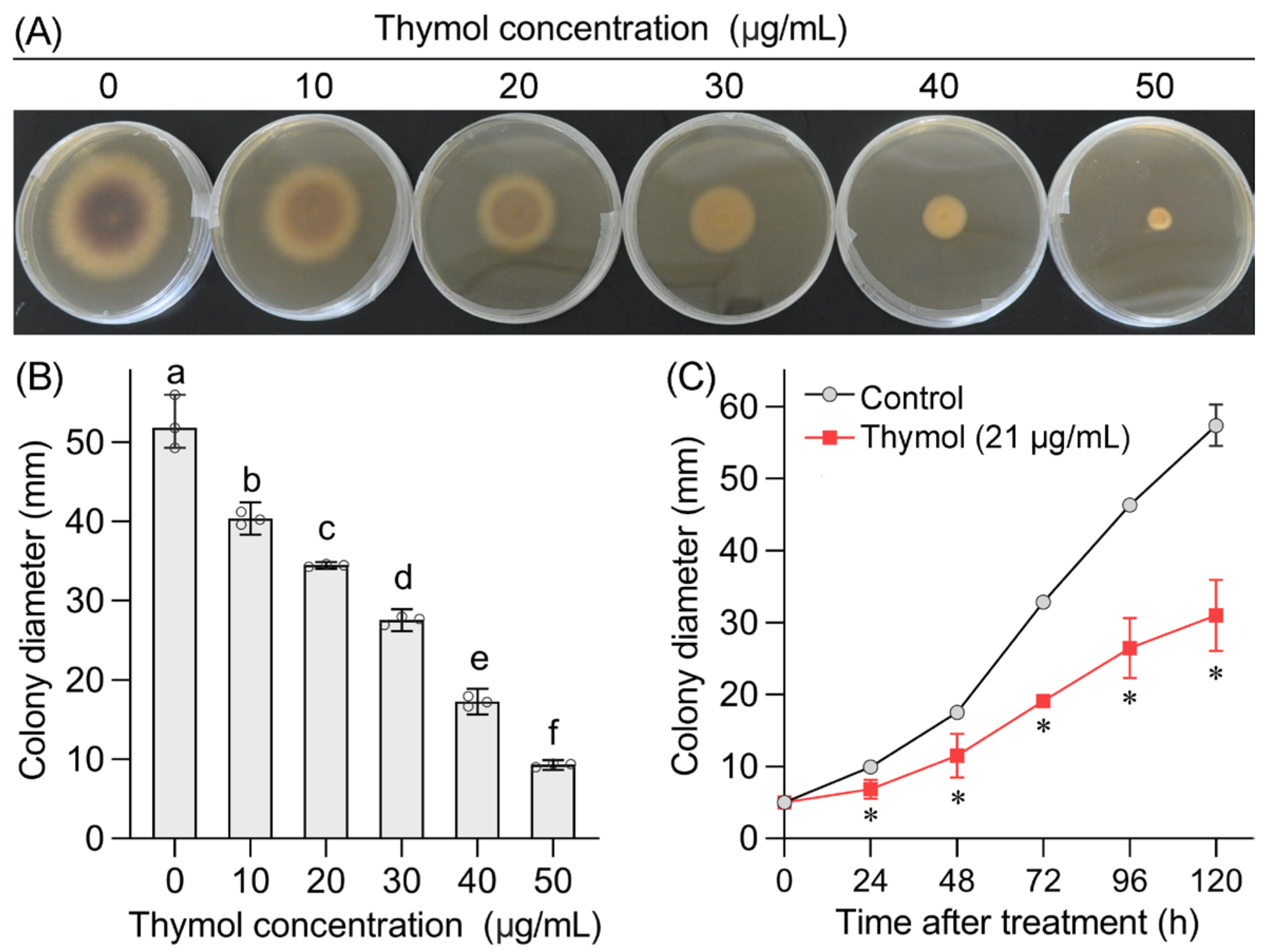
3.1. Thymol Inhibited Mycelial Growth of FON
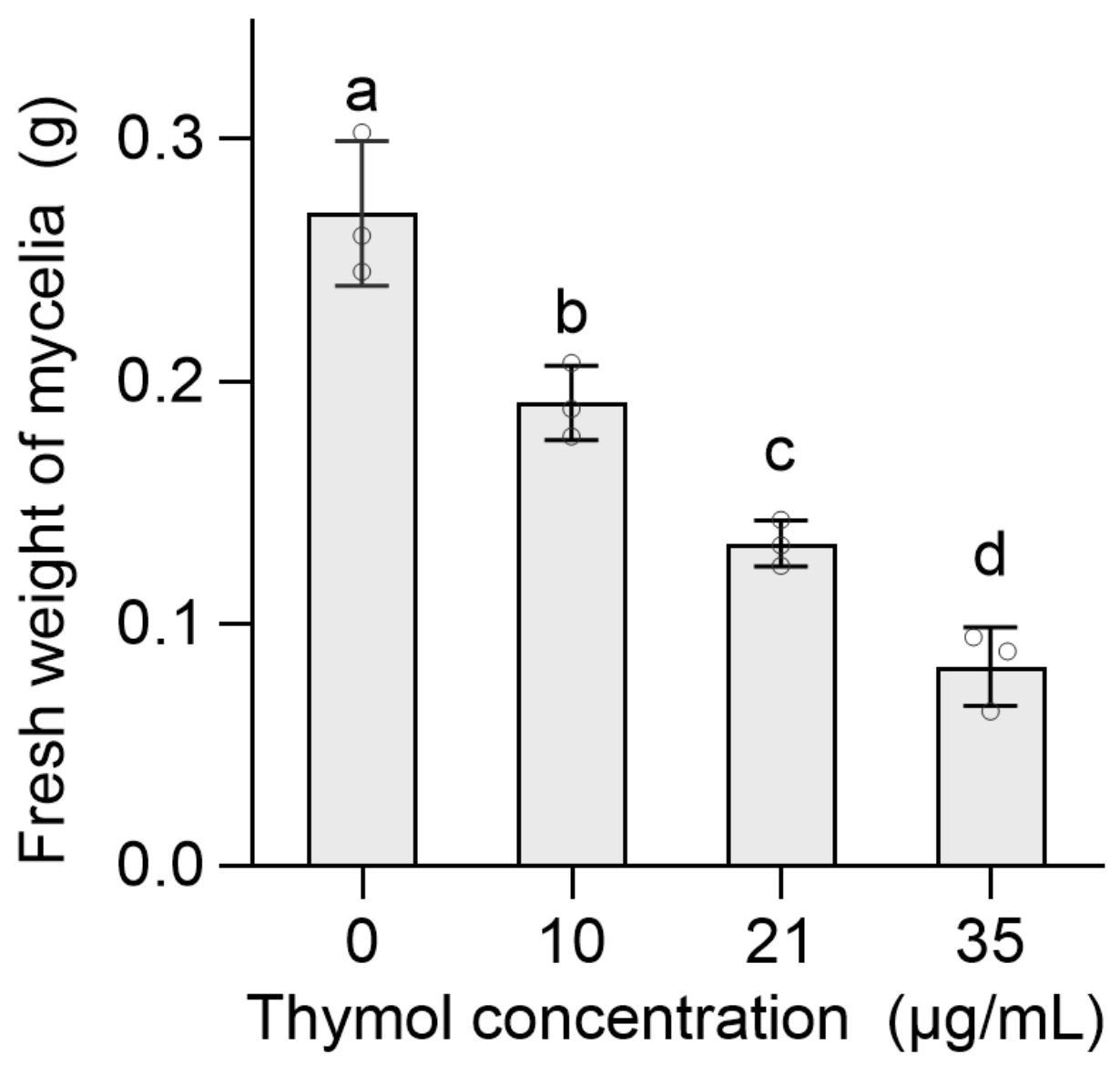
3.2. Thymol Damaged Mycelial Structure of FON
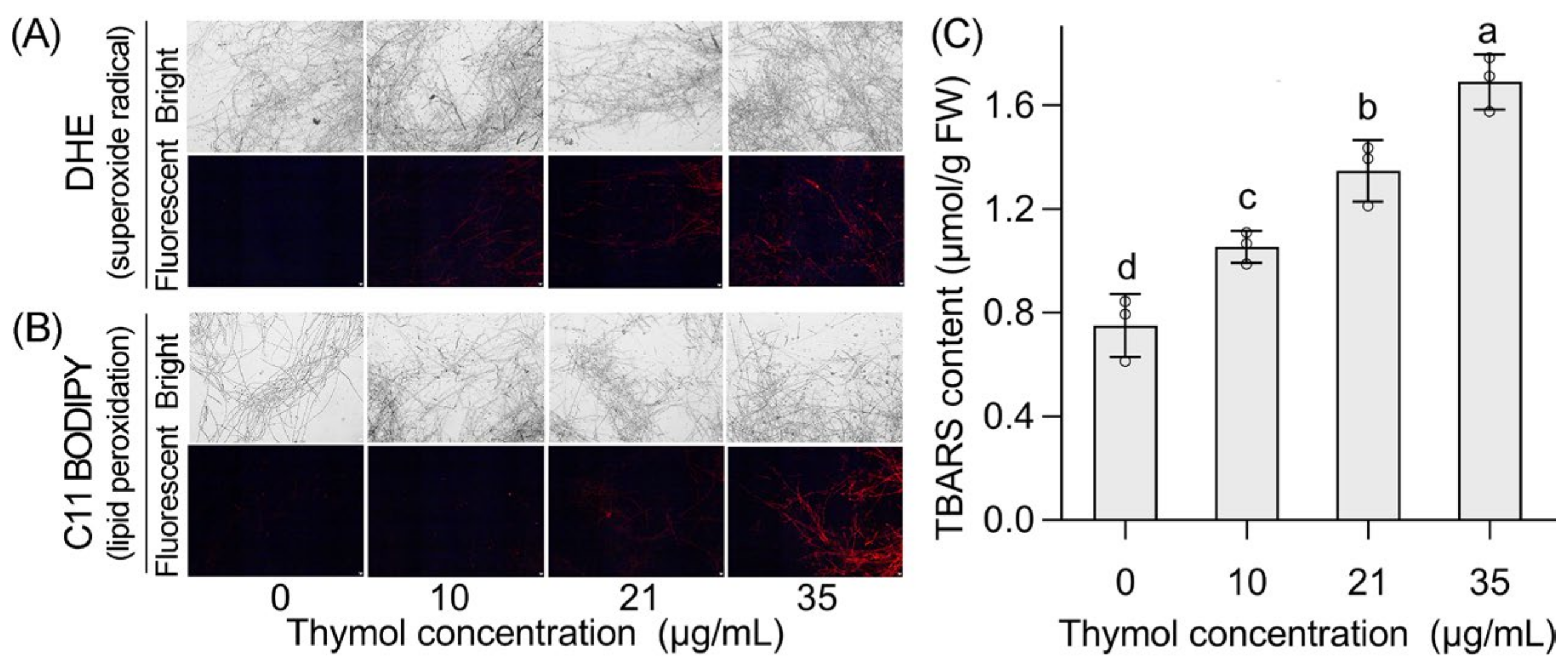
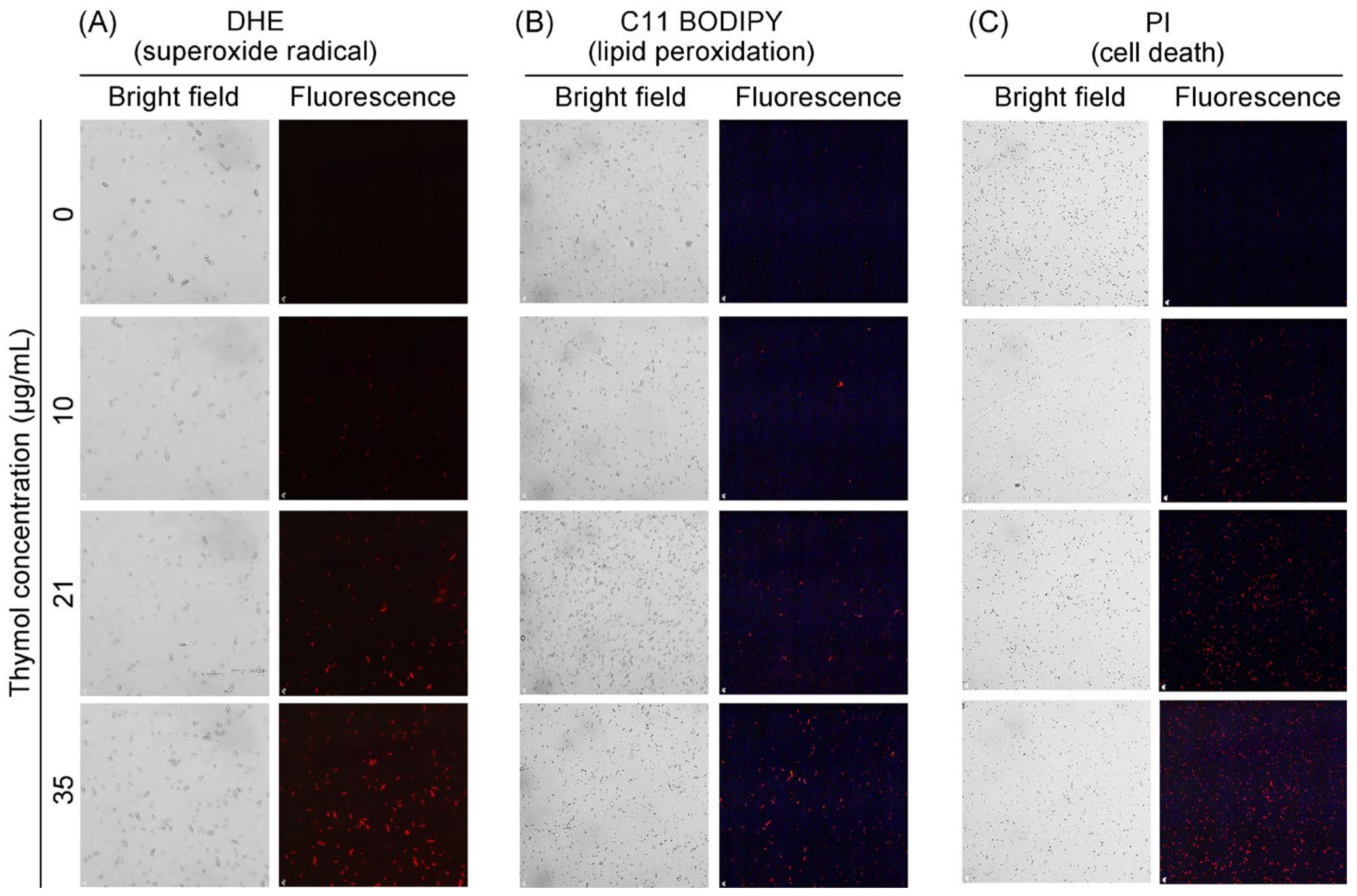
3.3. Thymol Induced Cell Death and Oxidative Injury in the Mycelia of FON
3.4. Thymol Induced Electrolyte Leakage and Osmotic Stress in the Mycelia of FON
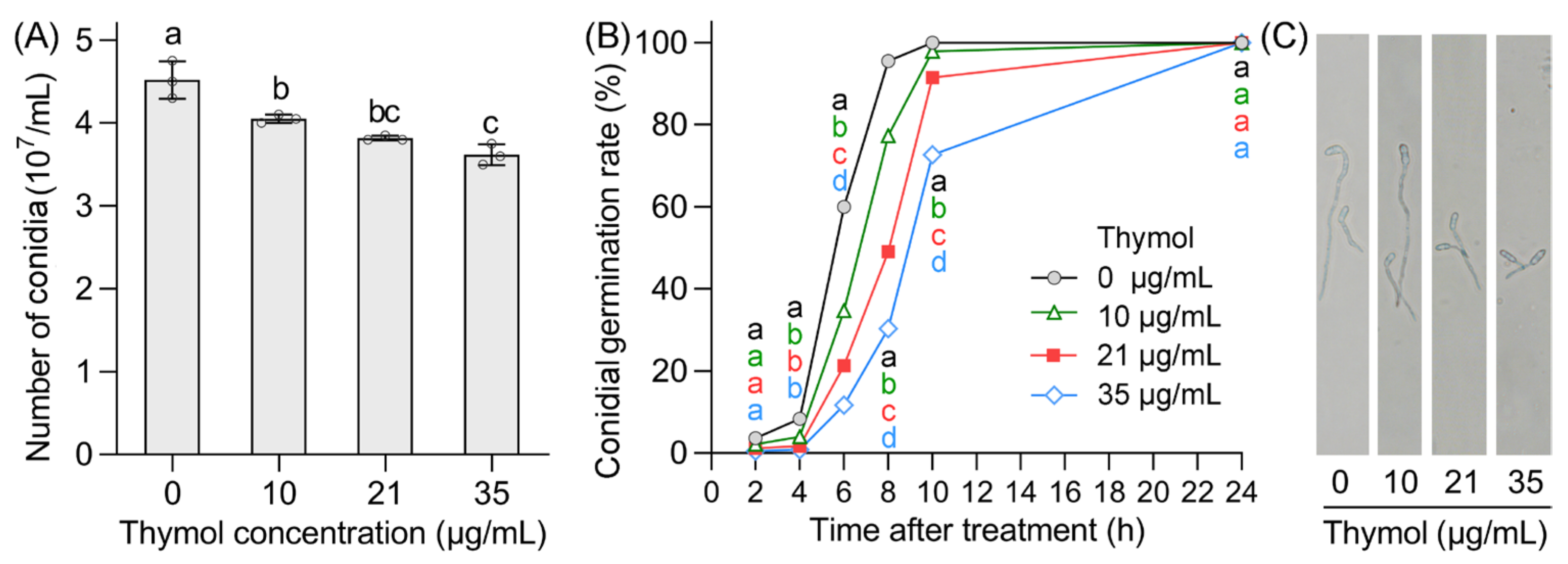
3.5. Thymol Inhibited Conidial Production and Conidial Germination of FON
3.6. Thymol Induced Oxidative Injury and Cell Death in the Conidia of FON
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gordon, T.R. Fusarium oxysporum and the Fusarium wilt syndrome. Annu. Rev. Phytopathol. 2017, 55, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Srinivas, C.; Nirmala, D.D.; Narasimha, M.K.; Mohan, C.D.; Lakshmeesha, T.R.; Singh, B.; Kalagatur, N.K.; Niranjana, S.R.; Hashem, A.; Alqarawi, A.A.; et al. Fusarium oxysporum f. sp. lycopersici causal agent of vascular wilt disease of tomato: Biology to diversity—A review. Saudi J. Biol. Sci. 2019, 26, 1315–1324. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Hsiang, T.; Luo, M.; Xiang, M. Draft genome sequence of an isolate of Fusarium oxysporum f. sp. melongenae, the causal agent of Fusarium Wilt of eggplant. Genome Announc. 2017, 5, e01597-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.; Zou, W.; Lin, S.; Onofua, D.; Yang, Z.; Chen, H.; Wang, S.; Chen, X. Transcriptome profiling and digital gene expression analysis of sweet potato for the identification of putative genes involved in the defense response against Fusarium oxysporum f. sp. batatas. PLoS ONE 2017, 12, e0187838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fulton, J.C.; Amaradasa, B.S.; Ertek, T.S.; Iriarte, F.B.; Sanchez, T.; Ji, P.; Paret, M.L.; Hudson, O.; Ali, M.E.; Dufault, N.S. Phylogenetic and phenotypic characterization of Fusarium oxysporum f. sp. niveum isolates from Florida-grown watermelon. PLoS ONE 2021, 16, e0248364. [Google Scholar] [CrossRef]
- Rebouças, T.A.; de Jesus Rocha, A.; Cerqueira, T.S.; Adorno, P.R.; Barreto, R.Q.; dos Santos Ferreira, M.; Lino, L.S.M.; de Oliveira Amorim, V.B.; dos Santos-Serejo, J.A.; Haddad, F.; et al. Pre-selection of banana somaclones resistant to Fusarium oxysporum f. sp. cubense, subtropical race 4. Crop Prot. 2021, 147, 105692. [Google Scholar] [CrossRef]
- Joshi, R. A review of Fusarium oxysporum on its plant interaction and industrial use. Crit. Rev. Microbiol. 2018, 6, 112–115. [Google Scholar] [CrossRef]
- Martyn, R.D. Fusarium wilt of watermelon: 120 years of research. Hortic. Rev. 2014, 42, 349–442. [Google Scholar]
- Kleczewski, N.M.; Egel, D.S. A diagnostic guide for Fusarium wilt of watermelon. Plant Health Prog. 2011, 12, 27. [Google Scholar] [CrossRef] [Green Version]
- Petkar, A.; Ji, P. Infection courts in watermelon plants leading to seed infestation by Fusarium oxysporum f. sp. niveum. Phytopathology 2017, 107, 828–833. [Google Scholar] [CrossRef] [Green Version]
- Hudson, O.; Hudson, D.; Ji, P.; Ali, M.E. Draft genome sequences of three Fusarium oxysporum f. sp. niveum isolates used in designing markers for race differentiation. Microbiol. Resour. Announc. 2020, 9, e01004-20. [Google Scholar] [CrossRef]
- Costa, A.E.D.S.; Cunha, F.S.D.; Araújo, K.M.G.; Lima Neto, I.S.; Capucho, A.S.; Borel, J.C.; Ishikawa, F.H. Morph-agronomic characterization of watermelon accessions with resistance to Fusarium Wilt. An. Acad. Bras. Cienc. 2021, 93, e20191359. [Google Scholar] [CrossRef]
- Hudson, O.; Fulton, J.C.; Dong, A.K.; Dufault, N.S.; Ali, M.E. Fusarium oxysporum f. sp. niveum molecular diagnostics past, present and future. Int. J. Mol. Sci. 2021, 22, 9735. [Google Scholar] [CrossRef]
- Bubici, G.; Kaushal, M.; Prigigallo, M.I.; Gómez-Lama Cabanás, C.; Mercado-Blanco, J. Biological control agents against Fusarium Wilt of banana. Front. Microbiol. 2019, 10, 616. [Google Scholar] [CrossRef] [Green Version]
- Niu, X.; Zhao, X.; Ling, K.S.; Levi, A.; Sun, Y.; Fan, M. The FonSIX6 gene acts as an avirulence effector in the Fusarium oxysporum f. sp. niveum-watermelon pathosystem. Sci. Rep. 2016, 6, 28146. [Google Scholar] [CrossRef] [Green Version]
- Keinath, A.P.; Hassell, R.L. Suppression of Fusarium Wilt caused by Fusarium oxysporum f. sp. niveum Race 2 on grafted triploid watermelon. Plant Dis. 2014, 98, 1326–1332. [Google Scholar] [CrossRef] [Green Version]
- Cháves-Gómez, J.L.; Becerra-Mutis, L.M.; Chávez-Arias, C.C.; Restrepo-Díaz, H.; Gómez-Caro, S. Screening of different Physalis genotypes as potential rootstocks or parents against vascular wilt using physiological markers. Front. Plant Sci. 2020, 11, 806. [Google Scholar] [CrossRef]
- An, M.; Zhou, X.; Wu, F.; Ma, Y.; Yang, P. Rhizosphere soil microorganism populations and community structures of different watermelon cultivars with differing resistance to Fusarium oxysporum f. sp. niveum. Can. J. Microbiol. 2011, 57, 355–365. [Google Scholar] [CrossRef]
- Zhou, X.; Li, C.; Liu, L.; Zhao, J.; Zhang, J.; Cai, Z.; Huang, X. Control of Fusarium wilt of lisianthus by reassembling the microbial community in infested soil through reductive soil disinfestation. Microbiol. Res. 2019, 220, 1–11. [Google Scholar] [CrossRef]
- Lv, H.; Cao, H.; Nawaz, M.A.; Sohail, H.; Huang, Y.; Cheng, F.; Kong, Q.; Bie, Z. Wheat intercropping enhances the resistance of watermelon to Fusarium Wilt. Front. Plant Sci. 2018, 9, 696. [Google Scholar] [CrossRef] [Green Version]
- Elmer, W.H.; Zuverza-Mena, N.; Triplett, L.R.; Roberts, E.L.; Silady, R.A.; White, J.C. Foliar application of copper oxide nanoparticles suppresses Fusarium wilt development on chrysanthemum. Environ. Sci. Technol. 2021, 55, 10805–10810. [Google Scholar] [CrossRef] [PubMed]
- Acosta-González, U.; Silva-Rojas, H.V.; Fuentes-Aragón, D.; Hernández-Castrejón, J.; Romero-Bautista, A.; Rebollar-Alviter, A. Comparative performance of fungicides and biocontrol products in the management of Fusarium Wilt of blackberry. Plant Dis. 2022, 106, 1419–1427. [Google Scholar] [CrossRef] [PubMed]
- Hudson, O.; Waliullah, S.; Ji, P.; Ali, M.E. Molecular characterization of laboratory mutants of Fusarium oxysporum f. sp. niveum resistant to prothioconazole, a demethylation inhibitor (DMI) fungicide. J. Fungi 2021, 7, 704. [Google Scholar] [CrossRef] [PubMed]
- Raza, W.; Ling, N.; Zhang, R.; Huang, Q.; Xu, Y.; Shen, Q. Success evaluation of the biological control of Fusarium wilts of cucumber, banana, and tomato since 2000 and future research strategies. Crit. Rev. Biotechnol. 2017, 37, 202–212. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Cai, X.Y.; Xu, M.; Tian, F. Enhanced biocontrol of cucumber Fusarium wilt by combined application of new antagonistic Bacteria Bacillus amyloliquefaciens B2 and phenolic acid-degrading fungus Pleurotus ostreatus P5. Front. Microbiol. 2021, 12, 700142. [Google Scholar] [CrossRef]
- Luo, W.; Liu, L.; Qi, G.; Yang, F.; Shi, X.; Zhao, X. Embedding Bacillus velezensis NH-1 in microcapsules for biocontrol of cucumber Fusarium Wilt. Appl. Environ. Microbiol. 2019, 85, e03128-18. [Google Scholar] [CrossRef] [Green Version]
- Zhai, Y.; Zhu, J.X.; Tan, T.M.; Xu, J.P.; Shen, A.R.; Yang, X.B.; Li, J.L.; Zeng, L.B.; Wei, L. Isolation and characterization of antagonistic Paenibacillus polymyxa HX-140 and its biocontrol potential against Fusarium wilt of cucumber seedlings. BMC Microbiol. 2021, 21, 75. [Google Scholar] [CrossRef]
- Khalil, M.M.R.; Fierro-Coronado, R.A.; Peñuelas-Rubio, O.; Villa-Lerma, A.G.; Plascencia-Jatomea, R.; Félix-Gastélum, R.; Maldonado-Mendoza, I.E. Rhizospheric bacteria as potential biocontrol agents against Fusarium wilt and crown and root rot diseases in tomato. Saudi J. Biol. Sci. 2021, 28, 7460–7471. [Google Scholar] [CrossRef]
- Marchese, A.; Orhan, I.E.; Daglia, M.; Barbieri, R.; Lorenzo, A.D.; Nabavi, S.F.; Gortzi, O.; Izadi, M.; Nabavi, S.M. Antibacterial and antifungal activities of thymol: A brief review of the literature. Food Chem. 2016, 210, 402–414. [Google Scholar] [CrossRef]
- Ahmad, A.; Vuuren, S.V.; Viljoen, A. Unravelling the complex antimicrobial interactions of essential oils—The case of Thymus vulgaris (thyme). Molecules 2014, 19, 2896–2910. [Google Scholar] [CrossRef] [Green Version]
- Palaniappan, K.; Holley, R.A. Use of natural antimicrobials to increase antibiotic susceptibility of drug resistant bacteria. Int. J. Food Microbiol. 2010, 140, 164–168. [Google Scholar] [CrossRef]
- Eloy, M.A.; Ribeiro, R.; Meireles, L.M.; Cutrim, T.A.S.; Francisco, C.S.; Javarini, C.L.; Borges, W.S.; Costa, A.V.; Queiroz, V.T.; Scherer, R.; et al. Thymol as an interesting building block for promising fungicides against Fusarium solani. J. Agric. Food Chem. 2021, 69, 6958–6967. [Google Scholar] [CrossRef]
- Gao, T.; Zhou, H.; Zhou, W.; Hu, L.B.; Chen, J.; Shi, Z.Q. The fungicidal activity of thymol against Fusarium graminearum via inducing lipid peroxidation and disrupting ergosterol biosynthesis. Molecules 2016, 21, 770. [Google Scholar] [CrossRef] [Green Version]
- Thomulka, K.W.; Abbas, C.G.; Young, D.A.; Lange, J.H. Evaluating median effective concentrations of chemicals with bioluminescent bacteria. Bull. Environ. Contam. Toxicol. 1996, 56, 446–452. [Google Scholar] [CrossRef]
- Yu, J.; Wang, W.N.; Matei, N.; Li, X.; Pang, J.W.; Mo, J.; Chen, S.P.; Tang, J.P.; Yan, M.; Zhang, J.H. Ezetimibe attenuates oxidative stress and neuroinflammation via the AMPK/Nrf2/TXNIP pathway after MCAO in rats. Oxid. Med. Cell. Longev. 2020, 2020, 4717258. [Google Scholar] [CrossRef]
- Domínguez-Rebolledo, A.E.; Martínez-Pastor, F.; Fernández-Santos, M.R.; del Olmo, E.; Bisbal, A.; Ros-Santaella, J.L.; Garde, J.J. Comparison of the TBARS assay and BODIPY C11 probes for assessing lipid peroxidation in red deer spermatozoa. Reprod. Domest. Anim. 2010, 45, e362. [Google Scholar]
- Kellermeier, F.; Chardon, F.; Amtmann, A. Natural variation of Arabidopsis root architecture reveals complementing adaptive strategies to potassium starvation. Plant. Physiol. 2013, 161, 1421–1432. [Google Scholar] [CrossRef] [Green Version]
- Duan, Y.; Ge, C.; Liu, S.; Chen, C.; Zhou, M. Effect of phenylpyrrole fungicide fludioxonil on morphological and physiological characteristics of Sclerotinia sclerotiorum. Pestic. Biochem. Physiol. 2013, 106, 61–67. [Google Scholar] [CrossRef]
- Yang, Y.; Fan, F.; Zhuo, R.; Ma, F.; Gong, Y.; Wan, X.; Jiang, M.; Zhang, X. Expression of the laccase gene from a white rot fungus in Pichia pastoris can enhance the resistance of this yeast to H2O2-mediated oxidative stress by stimulating the glutathione-based antioxidative system. Appl. Environ. Microbiol. 2012, 78, 5845–5854. [Google Scholar] [CrossRef] [Green Version]
- McCord, J.M.; Fridovich, I. Superoxide dismutase an enzymic function for erythrocuprein(hemocuprein). J. Biol. Chem. 1969, 244, 6049–6055. [Google Scholar] [CrossRef]
- Gao, C.; Fei, X.; Wang, M.; Chen, Q.; Zhao, N. Cardamomin protects from diabetes-induced kidney damage through modulating PI3K/AKT and JAK/STAT signaling pathways in rats. Int. Immunopharmacol. 2022, 107, 108610. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Wang, D.; Li, H.; Tang, Y.Q.; Ma, X.; Tang, H.Q.; Lin, M.; Liu, Z. Antifungal activity of an artificial peptide aptamer SNP-D4 against Fusarium oxysporum. Peer J. 2022, 10, e12756. [Google Scholar] [CrossRef] [PubMed]
- Du, R.; Liu, J.; Sun, P.; Li, H.; Wang, J. Inhibitory effect and mechanism of Tagetes erecta L. fungicide on Fusarium oxysporum f. sp. niveum. Sci. Rep. 2017, 7, 14442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dryden, M. Reactive oxygen species: A novel antimicrobial. Int. J. Antimicrob. Agents 2018, 51, 299–303. [Google Scholar] [CrossRef]
- Lee, J.E.; Seo, S.M.; Huh, M.J.; Lee, S.C.; Park, I.K. Reactive oxygen species mediated-antifungal activity of cinnamon bark (Cinnamomum verum) and lemongrass (Cymbopogon citratus) essential oils and their constituents against two phytopathogenic fungi. Pestic. Biochem. Physiol. 2020, 168, 104644. [Google Scholar] [CrossRef]
- Segal, L.M.; Wilson, R.A. Reactive oxygen species metabolism and plant-fungal interactions. Fungal Genet. Biol. 2018, 110, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Oiki, S.; Nasuno, R.; Urayama, S.I.; Takagi, H.; Hagiwara, D. Intracellular production of reactive oxygen species and a DAF-FM-related compound in Aspergillus fumigatus in response to antifungal agent exposure. Sci. Rep. 2022, 12, 13516. [Google Scholar] [CrossRef]
- Shen, Q.; Zhou, W.; Li, H.; Hu, L.; Mo, H. ROS involves the fungicidal actions of thymol against spores of Aspergillus flavus via the induction of nitric oxide. PLoS ONE 2016, 11, e0155647. [Google Scholar] [CrossRef] [Green Version]
- Larosa, V.; Remacle, C. Insights into the respiratory chain and oxidative stress. Biosci. Rep. 2018, 38, BSR20171492. [Google Scholar] [CrossRef] [Green Version]
- Yaakoub, H.; Mina, S.; Calenda, A.; Bouchara, J.P.; Papon, N. Oxidative stress response pathways in fungi. Cell Mol. Life Sci. 2022, 79, 333. [Google Scholar] [CrossRef]
- Dirschnabel, D.E.; Nowrousian, M.; Cano-Domínguez, N.; Aguirre, J.; Teichert, I.; Kück, U. New insights into the roles of NADPH oxidases in sexual development and ascospore germination in Sordaria macrospora. Genetics 2014, 196, 729–744. [Google Scholar] [CrossRef]
- Derr, A.M.; Faustoferri, R.C.; Betzenhauser, M.J.; Gonzalez, K.; Marquis, R.E.; Quivey, R.G., Jr. Mutation of the NADH oxidase gene (nox) reveals an overlap of the oxygen- and acid-mediated stress responses in Streptococcus mutans. Appl. Environ. Microbiol. 2012, 78, 1215–1227. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Zhang, X.; Ren, A.; Shi, D.K.; Shi, L.; Zhu, J.; Yu, H.S.; Zhao, M.W. Heat stress-induced reactive oxygen species participate in the regulation of HSP expression, hyphal branching and ganoderic acid biosynthesis in Ganoderma lucidum. Microbiol. Res. 2018, 209, 43–54. [Google Scholar] [CrossRef]
- Balan, D.J.; Rajavel, T.; Das, M.; Sathya, S.; Jeyakumar, M.; Devi, K.P. Thymol induces mitochondrial pathway-mediated apoptosis via ROS generation, macromolecular damage and SOD diminution in A549 cells. Pharmacol. Rep. 2021, 73, 240–254. [Google Scholar] [CrossRef]
- Yaakoub, H.; Sanchez, N.S.; Ongay-Larios, L.; Courdavault, V.; Calenda, A.; Bouchara, J.P.; Coria, R.; Papon, N. The high osmolarity glycerol (HOG) pathway in fungi†. Crit. Rev. Microbiol. 2022, 8, 657–695. [Google Scholar] [CrossRef]
- Jung, K.W.; Chung, M.S.; Bai, H.W.; Chung, B.Y.; Lee, S. Investigation of antifungal mechanisms of thymol in the human fungal pathogen, Cryptococcus neoformans. Molecules 2021, 26, 3476. [Google Scholar] [CrossRef]
- Huang, X.; Yi, Y.; Yong, J.; Sun, J.; Song, Z.; Li, D.; Li, Y. Inhibitory effect of berberine hydrochloride against Candida albicans and the role of the HOG-MAPK pathway. J. Antibiot. 2021, 74, 807–816. [Google Scholar] [CrossRef]
- Sephton-Clark, P.C.S.; Voelz, K. Spore germination of pathogenic filamentous fungi. Adv. Appl. Microbiol. 2018, 102, 117–157. [Google Scholar]
- Ortiz, S.C.; Huang, M.; Hull, C.M. Discovery of fungus-specific targets and inhibitors using chemical phenotyping of pathogenic spore germination. mBio. 2021, 12, e0167221. [Google Scholar] [CrossRef]
- Tudzynski, P.; Heller, J.; Siegmund, U. Reactive oxygen species generation in fungal development and pathogenesis. Curr. Opin. Microbiol. 2012, 15, 653–659. [Google Scholar] [CrossRef] [PubMed]
- Dou, S.; Liu, S.; Xu, X.; OuYang, Q.; Tao, N. Octanal inhibits spore germination of Penicillium digitatum involving membrane peroxidation. Protoplasma 2017, 254, 1539–1545. [Google Scholar] [CrossRef] [PubMed]
- Lai, T.; Li, B.; Qin, G.; Tian, S. Oxidative damage involves in the inhibitory effect of nitric oxide on spore germination of Penicillium expansum. Curr. Microbiol. 2011, 62, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, M.; Pereira, C.; Bessa, C.; Araujo, R.; Saraiva, L. Hydrogen peroxide-induced secondary necrosis in conidia of Aspergillus fumigatus. Can. J. Microbiol. 2016, 62, 95–101. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, M.; Stolp, Z.D.; Hardwick, J.M. Targeting intrinsic cell death pathways to control fungal pathogens. Biochem. Pharmacol. 2019, 162, 71–78. [Google Scholar] [CrossRef]
- Hu, L.B.; Ban, F.F.; Li, H.B.; Qian, P.P.; Shen, Q.S.; Zhao, Y.Y.; Mo, H.Z.; Zhou, X. Thymol induces conidial apoptosis in Aspergillus flavus via stimulating K+ eruption. J. Agric. Food Chem. 2018, 66, 8530–8536. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hao, Y.; Zhang, J.; Sun, C.; Chen, X.; Wang, Y.; Lu, H.; Chen, J.; Shi, Z.; Zhang, L.; Yang, L.; et al. Thymol Induces Cell Death of Fusarium oxysporum f. sp. niveum via Triggering Superoxide Radical Accumulation and Oxidative Injury In Vitro. Agronomy 2023, 13, 189. https://doi.org/10.3390/agronomy13010189
Hao Y, Zhang J, Sun C, Chen X, Wang Y, Lu H, Chen J, Shi Z, Zhang L, Yang L, et al. Thymol Induces Cell Death of Fusarium oxysporum f. sp. niveum via Triggering Superoxide Radical Accumulation and Oxidative Injury In Vitro. Agronomy. 2023; 13(1):189. https://doi.org/10.3390/agronomy13010189
Chicago/Turabian StyleHao, Yini, Jiao Zhang, Changwei Sun, Xuenai Chen, Yuxiao Wang, Haiyan Lu, Jian Chen, Zhiqi Shi, Li Zhang, Lifei Yang, and et al. 2023. "Thymol Induces Cell Death of Fusarium oxysporum f. sp. niveum via Triggering Superoxide Radical Accumulation and Oxidative Injury In Vitro" Agronomy 13, no. 1: 189. https://doi.org/10.3390/agronomy13010189
APA StyleHao, Y., Zhang, J., Sun, C., Chen, X., Wang, Y., Lu, H., Chen, J., Shi, Z., Zhang, L., Yang, L., & Huang, S. (2023). Thymol Induces Cell Death of Fusarium oxysporum f. sp. niveum via Triggering Superoxide Radical Accumulation and Oxidative Injury In Vitro. Agronomy, 13(1), 189. https://doi.org/10.3390/agronomy13010189