
Free-Grazing versus Enclosure Lead to an Increase in the Germination of the Leymus chinensis Seed Bank in the Hulunbuir Grassland





Abstract
:1. Introduction
2. Materials and Methods
2.1. Overview of the Experimental Site
2.2. Soil Seed Bank Sampling
2.3. Seed Bank Germination Test
2.4. Data Analysis
- L. chinensis germination data were collected over 45 days. Additionally, the numbers of germinable L. chinensis seeds in the enclosed and free-grazing areas in the 0–2 cm, 2–5 cm, and 5–10 cm soil layers were compared.
- The data were summarized and averaged.
- A curve chart was drawn in SigmaPlot 14.0 to compare the germination characteristics of L. chinensis in the 0–2 cm, 2–5 cm, 5–10 cm, and 0–10 cm soil layers in the free-grazing areas and enclosed areas.
3. Results
3.1. Effects of the Grazing Area and Enclosed Area on the Soil Germinable Seed Bank
3.2. Effects of Grazing Area and Enclosed Area on the Germinable Seed Bank of Leymus chinensis in Soil
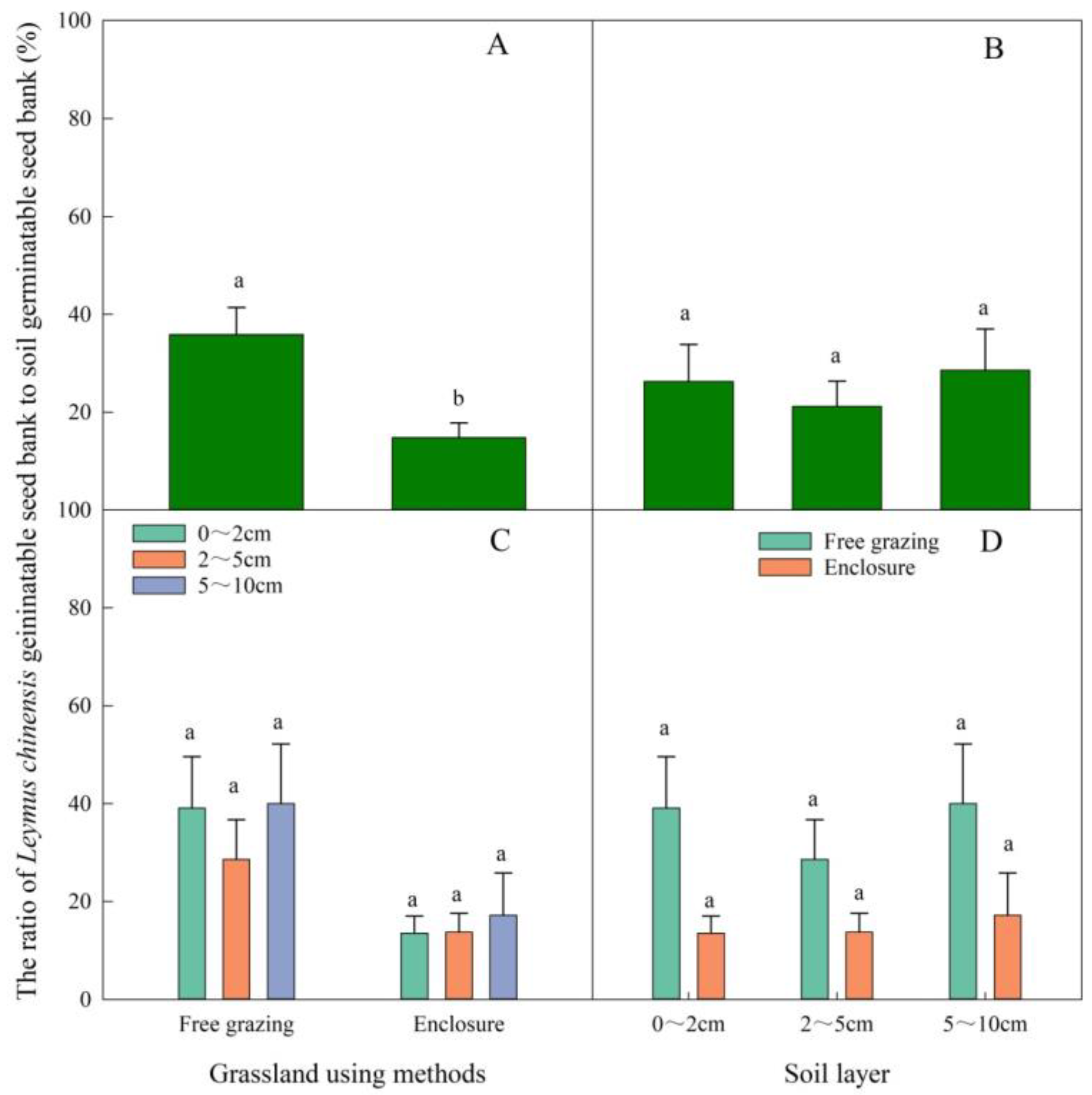
3.3. Effects of Grazing and Enclosed Areas on the Ratio of the Germinable Leymus chinensis Seed Bank to Overall Soil Germinable Seed Bank
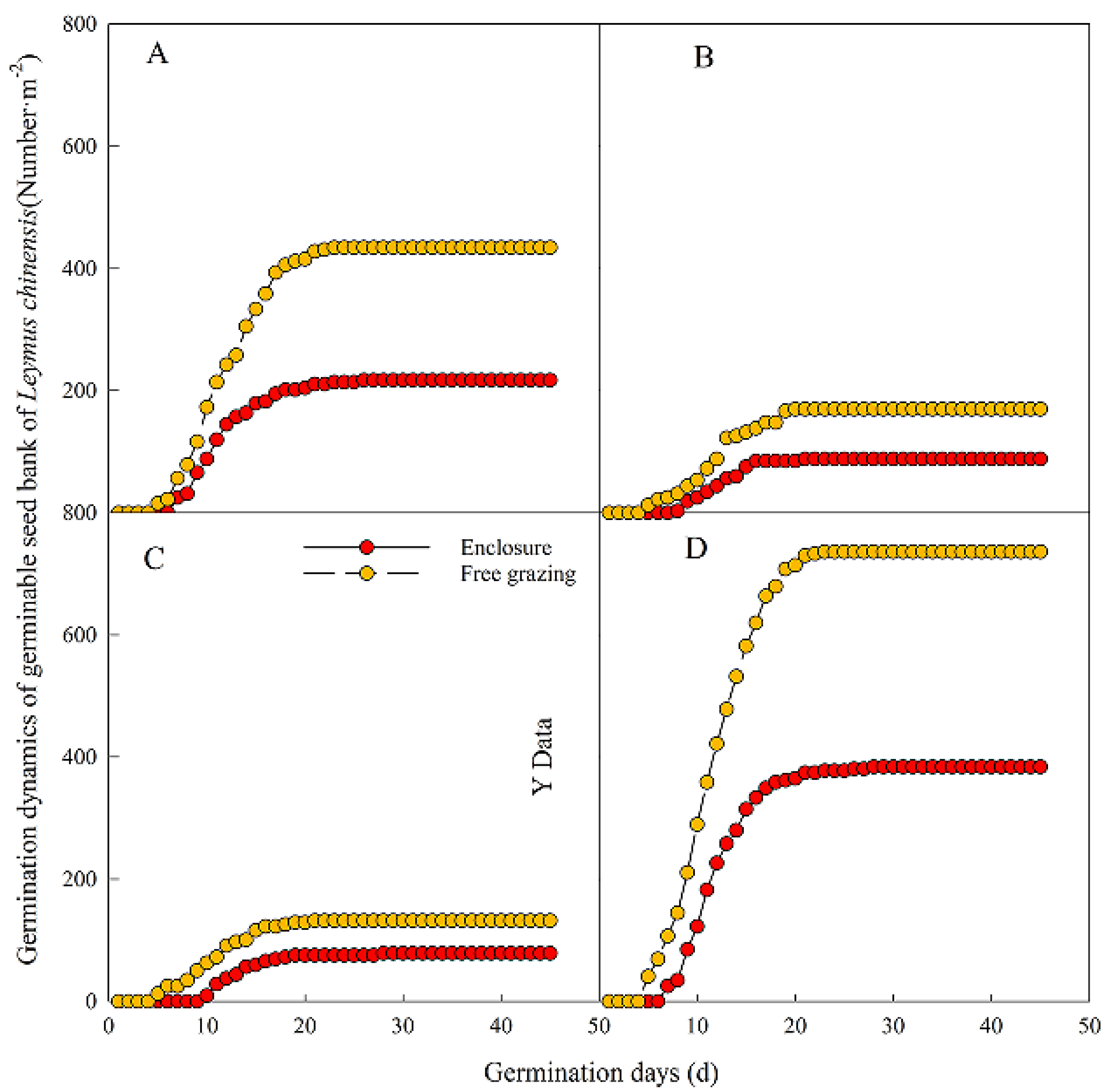
3.4. Effects of the Grazing and Enclosed Areas on the Germination Dynamics of the Leymus chinensis Seed Bank
4. Discussion
4.1. Effect of Grazing on the Germinable Leymus chinensis Seed Bank
4.2. Effect of Grazing on Seed Source of Germinating Seed Bank of Leymus chinensis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kinucan, R.J.; Smeins, F.E. Soil seed bank of a semiarid Texas grassland under three long-term (36 years) grazing regimes. Am. Midl. Nat. 1992, 128, 11–21. [Google Scholar] [CrossRef]
- Gasparino, D.; Malavasi, U.C.; Malavasi, M.M.; Souza, I. Quantificação do banco de sementes sob diferentes usos do solo em área de domínio ciliar. Rev. Arvore 2006, 30, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Lezama, F.; Baeza, S.; Altesor, A.; Cesa, A.; Chaneton, E.J.; Paruelo, J.M. Variation of grazing-induced vegetation changes across a large-scale productivity gradient. J. Veg. Sci. 2014, 24, 8–21. [Google Scholar] [CrossRef]
- Loydi, A. Effects of grazing exclusion on vegetation and seed bank composition in a mesic mountain grassland in Argentina. Plant Ecol. Divers. 2019, 12, 127–138. [Google Scholar] [CrossRef]
- Gao, Y.; An, Y.; Qi, B.; Liu, J.; Yu, H.Z.; Wang, D.J. Grazing exclusion mediates the trade-off between plant diversity and productivity in Leymus Chinensis meadows along a chronosequence on the Songnen Plain, China. Ecol. Indic. 2021, 126, 107655. [Google Scholar] [CrossRef]
- Bekker, R.M.; Verweij, G.L.; Bakker, J.P.; Fresco, L.F.M. Soil seed bank dynamics in hayfield succession. J. Ecol. 2000, 4, 594–607. [Google Scholar] [CrossRef]
- Royo, A.A.; Ristau, T.E. Stochastic and deterministic processes regulate spatio-temporal variation in seed bank diversity. J. Veg. Sci. 2013, 24, 724–734. [Google Scholar] [CrossRef] [Green Version]
- Harrod, R.J.; Halpem, C.B. The seed ecology of Iliamna longisepala (Torr.) Wiggins, an east Cascade endemic. Nat. Area J. 2005, 25, 246–256. [Google Scholar]
- Thompson, K. The functional ecology of soil seed banks. In Seeds the Ecology of Regeneration in Plant Communities; CABI Publishing: Wallingford, UK, 2000; pp. 215–236. [Google Scholar]
- Grime, J.P.; Hillier, S.H. The contribution of seedling regeneration to the structure and dynamics of plant communities, ecosystems and larger units of the landscape. In The Ecology of Regeneration in Plant Communities; CABI Publishing: Wallingford, UK, 2000; pp. 361–374. [Google Scholar]
- Szabó, A.; Ruprecht, E. Restoration possibilities of dry grasslands afforested by pine: The role of seed bank and remnant vegetation. Tuexenia 2018, 38, 405–418. [Google Scholar] [CrossRef]
- Hao, L.; Sun, G.; Liu, Y.Q.; Gao, Z.Q.; He, J.J.; Shi, T.T.; Wu, B.J. Effects of precipitation on grassland ecosystem restoration under grazing exclusion in Inner Mongolia, China. Landsc. Ecol. 2014, 29, 1657–1673. [Google Scholar] [CrossRef]
- Dong, S.K.; Sherman, R. Enhancing the resilience of coupled human and natural systems of alpine rangelands on the Qinghai-Tibetan Plateau. Rangel. J. 2015, 37, i–iii. [Google Scholar] [CrossRef]
- Wang, R.Z.; Yuan, Y.Q. Seed productions and seed banks in Leymus Chinensis in response to environment changes in the Songnen plain; northeastern China. Int. J. Environ. Sci. Technol. 2001, 27, 163–169. [Google Scholar]
- Liu, G.X.; Han, J.G. Seedling establishment of wild and cultivated Leymus chinensis (Trin.) Tzvel. under different seeding depths. J. Arid Environ. 2008, 72, 279–284. [Google Scholar] [CrossRef]
- Lin, J.X.; Shi, Y.J.; Tao, S.; Yu, X.Y.; Yu, D.F.; Yan, X.F. Seed-germination response of Leymus chinensis to cold stratification in a range of temperatures; light and low water potentials under salt and drought stresses. Crop Pasture Sci. 2017, 68, 188–194. [Google Scholar] [CrossRef]
- Chen, S.Y.; Huang, X.; Yan, X.Q.; Liang, Y.; Wang, Y.Z.; Li, X.F.; Peng, X.J. Transcriptome Analysis in Sheepgrass (Leymus chinensis): A Dominant Perennial Grass of the Eurasian Steppe. PLoS ONE 2013, 8, e67974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Deng, Y.K.; Yu, X.H.; Wu, G.H.; Gao, Y.B.; Ren, A.Z. Epichloë Endophyte infection changes the root endosphere microbial community composition of Leymus chinensis under both potted and field growth conditions. Microb. Ecol. 2022, 2, 1–13. [Google Scholar] [CrossRef]
- Birhanu, L.; Tamrat, B.; Binyam, T.; Sebsebe, D. Soil seed bank composition and aboveground vegetation in dry afromontane forest patches of northwestern Ethiopia. Trees For. People 2022, 9, 100292. [Google Scholar] [CrossRef]
- Albrecht, H.; Eder, E.; Langbehn, T.; Tschiersch, C. The soil seed bank and its relationship to the established vegetation in urban wastelands. Landsc. Urban Plan 2011, 100, 87–97. [Google Scholar] [CrossRef]
- Loydi, A.; Zalba, S.M.; Distel, R.A. Viable seed banks under grazing and exclosure conditions in montane mesic grasslands of argentina. Acta Oecol. 2021, 43, 8–15. [Google Scholar] [CrossRef]
- Cao, L.; Guo, C.; Chen, J. Fluctuation in seed abundance has contrasting effects on the fate of seeds from two rapidly germinating tree species in an Asian tropical forest. Integr. Zool. 2017, 12, 2–11. [Google Scholar] [CrossRef]
- Gonzalez, S.L.; Ghermandi, L. Overgrazing causes a reduction in the vegetation cover and seed bank of Patagonian grasslands. Plant Soil 2021, 464, 75–87. [Google Scholar] [CrossRef]
- Gongbuzeren, Y.; Li, W. China’s rangeland management policy debates: What have we learned. Rangel. Ecol. Manag. 2015, 68, 305–314. [Google Scholar] [CrossRef]
- Hamilton, E.W.; Frank, D.A. Can plants stimulate soil microbes and their own nutrient supply? Evidence from a grazing tolerant grass. Ecology 2001, 9, 2397–2402. [Google Scholar] [CrossRef]
- Schönbach, H.W.; Wan, M.; Gierus, Y.F.; Bai, K.; Müller, L.J.; Lin, A.; Susenbeth, F. Grassland responses to grazing: Effects of grazing intensity and management system in an Inner Mongolian steppe ecosystem. Plant Soil Environ. 2011, 340, 103–115. [Google Scholar] [CrossRef]
- Onatibia, G.R.; Aguiar, M.R. Grasses and grazers in arid rangelands: Impact of sheep management on forage and non-forage grass populations. J. Environ. Manag. 2019, 235, 42–50. [Google Scholar] [CrossRef]
- Ma, H.Y.; Liang, Z.W.; Wang, Z.C.; Chen, Y.; Huang, L.H.; Yang, F. Lemmas and endosperms significantly inhibited germination of Leymus chinensis (Trin.) Tzvel. (Poaceae). J. Arid Environ. 2008, 72, 573–578. [Google Scholar] [CrossRef]
- Zhang, W.D.; Chen, S.Y.; Liu, G.S.; Jan, C.C. Seed-set and pollen-stigma compatibility in Leymus chinensis. Grass Forage Sci. 2004, 59, 180–185. [Google Scholar] [CrossRef]
- Yang, Y.F.; Liu, G.C.; Zhang, B.T. Ananalysis of age structure and the strategy for asexual propagation of Aneurolepidium chinense (Trin.) Kitag population. Acta Oceanol. Sin. 1995, 37, 147–153. [Google Scholar]
- Liu, G.S.; Qi, D.M. Research progress on the biology of Leymus chinensis. Acta Pratacult. Sin. 2004, 13, 6–11. [Google Scholar]
- Zhang, W.D.; Bi, J.J.; Ning, T.Y.; Liu, X.L.; He, M.R. Effects of temperature; light and other treatments on seed germination of Leymus chinensis. Can. J. Plant Sci. 2006, 86, 143–148. [Google Scholar] [CrossRef]
- Badgery, W.B.; Kemp, D.R.; Zhang, Y.J.; Wang, Z.W.; Han, G.D.; Hou, F.J.; Liu, N.; David, M.; Karl, B. Optimising grazing for livestock production and environmental benefits in Chinese grasslands. Rangel. J. 2020, 42, 347–358. [Google Scholar] [CrossRef]
- Ma, H.Y.; Li, J.P.; Yang, F.; Lü, X.T.; Pan, Y.P.; Liang, Z.W. Regenerative role of soil seed banks of different successional stages in a saline-alkaline grassland in northeast China. Chin. Geogr. Sci. 2018, 28, 154–166. [Google Scholar] [CrossRef] [Green Version]
- Silva, G.; Overbeck, G.E. Soil seed bank in a subtropical grassland under different grazing intensities. Acta Bot. Bras. 2020, 34, 360–370. [Google Scholar] [CrossRef]
- Chen, T.; Michael, C.; Nan, Z.B.; Hou, F.J. Effects of grazing intensity on seed size; germination and fungal colonization of Lespedeza davurica in a semi-arid grassland of northwest China. J. Arid Environ. 2017, 144, 91–97. [Google Scholar] [CrossRef]
- Verdoodt, A.; Mureithi, S.M.; Ranst, E.V. Impacts of management and enclosure age on recovery of the herbaceous rangeland vegetation in semi-arid Kenya. J. Arid Environ. 2010, 74, 1066–1073. [Google Scholar] [CrossRef]
- Wang, R.Z.; Ripley, E.A. Effects of grazing on a Leymus chinensis grassland on the Songnen plain of northeastern China. J. Arid Environ. 1997, 36, 307–318. [Google Scholar] [CrossRef]
- Qing, X.L. Studies on Reproductive Characteristics of Plants in Typical Steppe Community. Ph.D Thesis, Inner Mongolia Agricultural University, Huhhot, Mongolia.
- Wang, R.Z.; Ripley, E.A. The effects of grazing and mowing on a Leymus chinensis grassland in the Songnen plain of northeastern China. Int. J. Ecol. Environ. 1998, 24, 383–391. [Google Scholar]
- Blumenthal, M.J.; Ison, R.L. Plant population dynamics in subterranean clover and murex medic swards. 3. Effect of pod burial; summer grazing and autumn cultivation on emergence. Anim. Prod. Sci. 1996, 36, 533–538. [Google Scholar] [CrossRef]
- Smith, S.E.; Fendenheim, M.D. Seed Production in Sideoats Grama Populations with Different Grazing Histories. J. Oper. Manag. 2000, 53, 550–555. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Location | ||
|---|---|---|---|
| Team 6 | Team 11 | Team 12 | |
| Longitude | 120°2′47″–3′36″ E | 120°6′50″–7′28″ E | 120°2′47″–3′36″ E |
| Latitude | 49°19′32″–19′51″ N | 49°20′52″–21′16″ N | 49°19′32″–19′51″ N |
| Annual average temperature | −2.2 °C | −2.4 °C | −2.2 °C |
| Annual precipitation | 351 mm | 350 mm | 351 mm |
| Soil category | chestnut calcareous soil | chestnut calcareous soil | chestnut calcareous soil |
| Main plant populations | Leymus chinensis, Stipa baicalensis, Artemisia tanacetifolia, Pulsatilla turczaninovii, Cleistogenes squarrosa | Leymus chinensis, Stipa baicalensis, Artemisia tanacetifolia, Pulsatilla turczaninovii, Cleistogenes squarrosa | Leymus chinensis, Stipa baicalensis,Artemisia tanacetifolia, Pulsatilla turczaninovii, Cleistogenes squarrosa |
| Dominant species | Leymus chinensis | Stipa baicalensis and Leymus chinensis | Leymus chinensis |
| Treatment | Free-grazing area, enclosed area | Free-grazing area, enclosed area | Free-grazing area, enclosed area |
| Soil Layer | Grassland Use Method | Initial Germination Days | Germination Termination Days | Germination Duration Days | Germination Index |
|---|---|---|---|---|---|
| 0–10 cm | Enclosed area | 8.63 ± 0.42 a | 20.38 ± 1.76 | 12.75 ± 1.92 | 35.44 ± 11.78 |
| Free-grazing area | 7.00 ± 0.55 b | 19.11 ± 1.02 | 13.11 ± 1.20 | 34.59 ± 6.95 | |
| 0–2 cm | Enclosed area | 7.67 ± 0.67 | 22.00 ± 2.65 | 15.33 ± 3.28 | 72.98 ± 22.02 |
| Free-grazing area | 6.33 ± 0.67 | 22.00 ± 0.58 | 16.67 ± 0.88 | 43.4 ± 18.32 | |
| 2–5 cm | Enclosed area | 8.67 ± 0.33 | 17.00 ± 2.08 | 9.33 ± 2.40 | 17.61 ± 4.91 |
| Free-grazing area | 7.33 ± 1.20 | 18.00 ± 1.53 | 11.67 ± 2.33 | 33.97 ± 8.65 | |
| 5–10 cm | Enclosed area | 10.00 ± 0.00 | 23.00 ± 5.00 | 14.00 ± 5.00 | 15.74 ± 9.89 |
| Free-grazing area | 7.33 ± 1.20 | 17.33 ± 1.86 | 11.00 ± 1.15 | 26.41 ± 9.81 |
| Grassland Use Method | Soil Layer | Initial Germination Days | Germination Termination Days | Germination Duration Days | Germination Index |
|---|---|---|---|---|---|
| Total | 0–2 cm | 7.00 ± 0.52 | 22.00 ± 1.21 | 16.00 ± 1.55 | 58.19 ± 14.42 a |
| 2–5 cm | 8.00 ± 0.63 | 17.50 ± 1.18 | 10.50 ± 1.59 | 25.79 ± 5.76 b | |
| 5–10 cm | 8.40 ± 0.93 | 19.60 ± 2.34 | 12.20 ± 1.85 | 21.08 ± 6.67 b | |
| Enclosed area | 0–2 cm | 7.67 ± 0.67 b | 22.00 ± 2.65 | 15.33 ± 3.28 | 72.98 ± 22.02 a |
| 2–5 cm | 8.67 ± 0.33 ab | 17.00 ± 2.08 | 9.33 ± 2.40 | 17.61 ± 4.91 b | |
| 5–10 cm | 10.00 ± 0.00 a | 23.00 ± 5.00 | 14.00 ± 5.00 | 15.74 ± 9.89 b | |
| Free-grazing area | 0–2 cm | 6.33 ± 0.67 | 22.00 ± 0.58 | 16.67 ± 0.88 | 43.4 ± 18.32 |
| 2–5 cm | 7.33 ± 1.20 | 18.00 ± 1.53 | 11.67 ± 2.33 | 33.97 ± 8.65 | |
| 5–10 cm | 7.33 ± 1.20 | 17.33 ± 1.86 | 11.00 ± 1.15 | 26.41 ± 9.81 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Wu, Y.; Li, Y.; Lv, S.; Wei, Z.; Chen, B.; Xu, L.; Yang, G.; Xin, X.; Yan, R. Free-Grazing versus Enclosure Lead to an Increase in the Germination of the Leymus chinensis Seed Bank in the Hulunbuir Grassland. Agronomy 2023, 13, 22. https://doi.org/10.3390/agronomy13010022
Liu H, Wu Y, Li Y, Lv S, Wei Z, Chen B, Xu L, Yang G, Xin X, Yan R. Free-Grazing versus Enclosure Lead to an Increase in the Germination of the Leymus chinensis Seed Bank in the Hulunbuir Grassland. Agronomy. 2023; 13(1):22. https://doi.org/10.3390/agronomy13010022
Chicago/Turabian StyleLiu, Hongmei, Yanling Wu, Yingxin Li, Shijie Lv, Zhijun Wei, Baorui Chen, Lijun Xu, Guixia Yang, Xiaoping Xin, and Ruirui Yan. 2023. "Free-Grazing versus Enclosure Lead to an Increase in the Germination of the Leymus chinensis Seed Bank in the Hulunbuir Grassland" Agronomy 13, no. 1: 22. https://doi.org/10.3390/agronomy13010022
APA StyleLiu, H., Wu, Y., Li, Y., Lv, S., Wei, Z., Chen, B., Xu, L., Yang, G., Xin, X., & Yan, R. (2023). Free-Grazing versus Enclosure Lead to an Increase in the Germination of the Leymus chinensis Seed Bank in the Hulunbuir Grassland. Agronomy, 13(1), 22. https://doi.org/10.3390/agronomy13010022