
Effects of Arbuscular Mycorrhizal Fungi on Growth and Nutrient Accumulation of Oat under Drought Conditions

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Materials and Experimental Design
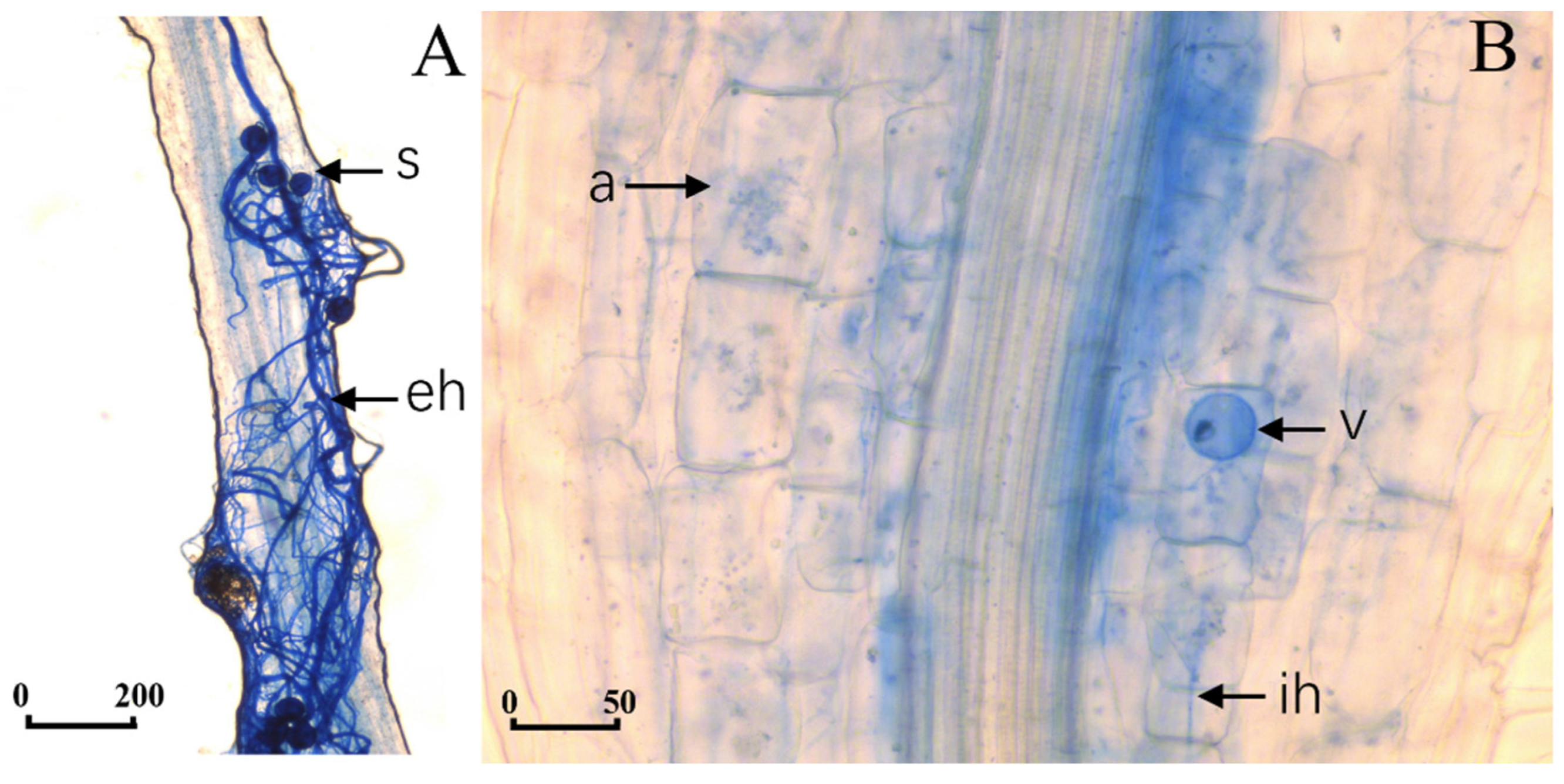
2.2. Mycorrhizal Colonization and Plant Growth Parameters
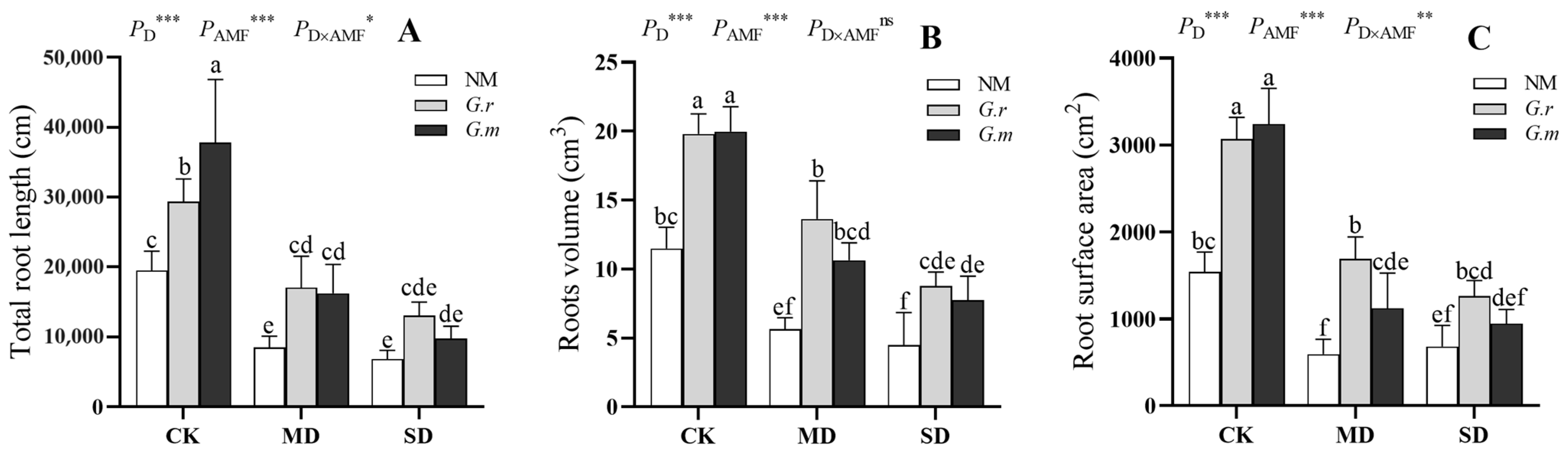
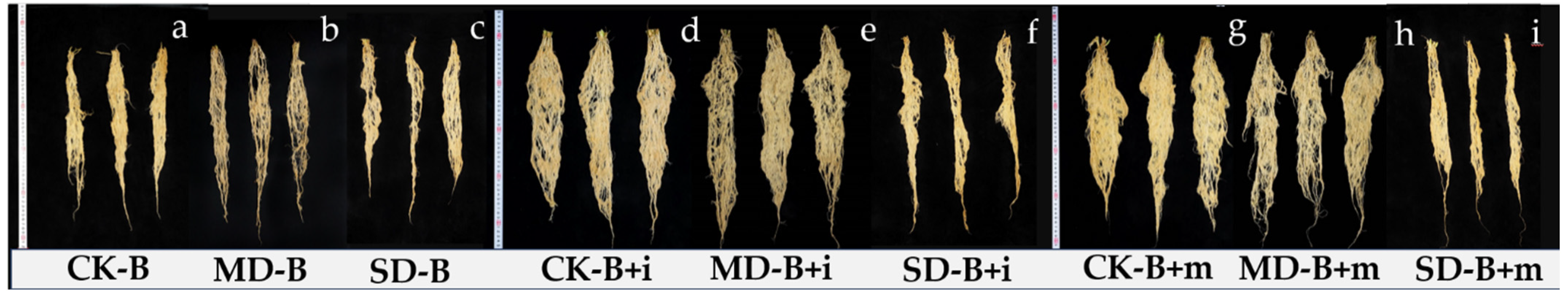
2.3. Method for Determination of Root Architecture
2.4. TSS, MDA, and Protein Concentrations
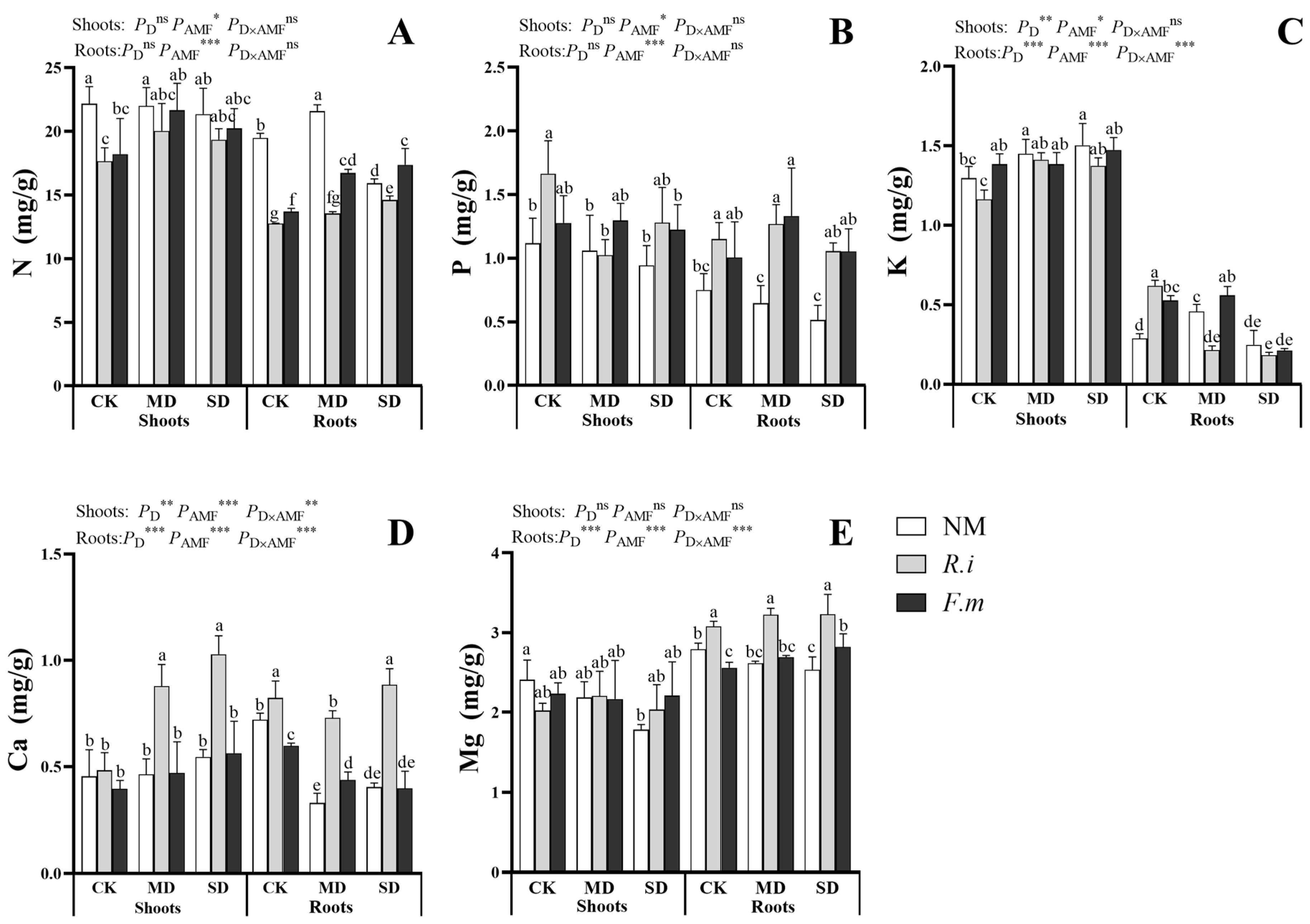
2.5. Macronutrient Concentrations in Oats
2.6. Statistical Analysis
3. Results
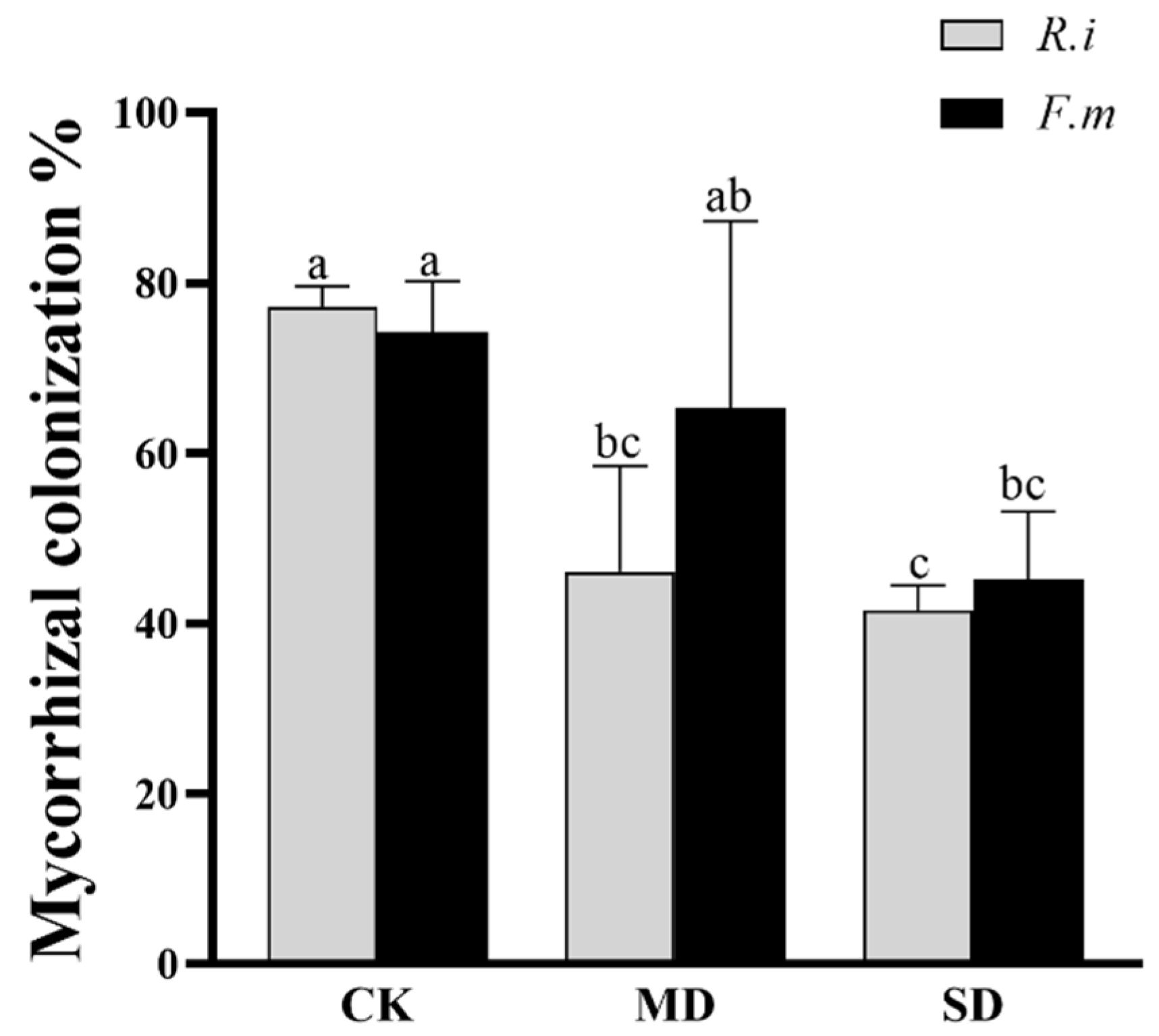
3.1. Mycorrhizal Colonization
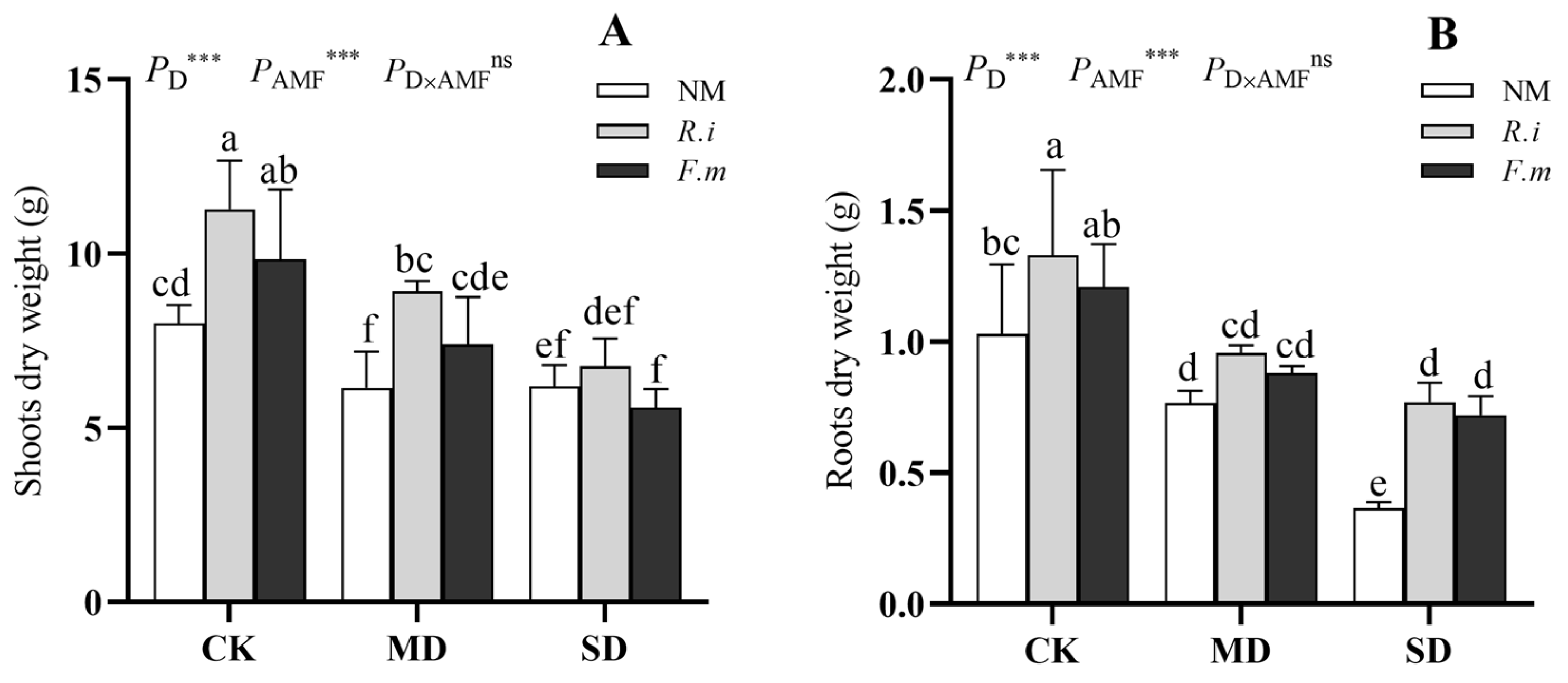
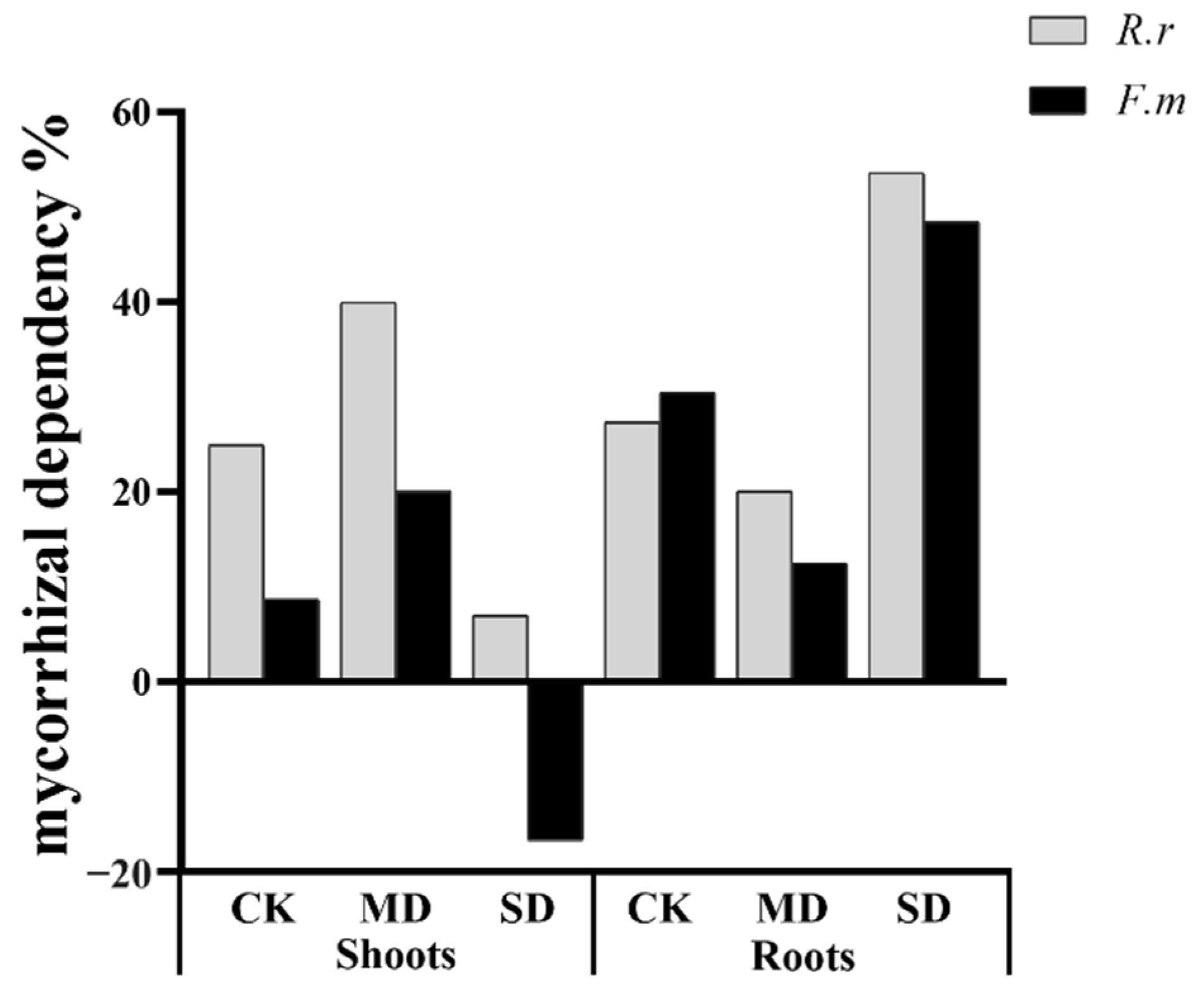
3.2. Plant Growth of Shoots and Roots
3.3. TSS, MDA, and Protein Contents
3.4. Macronutrient Concentrations in Oats
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mushtaq, A.; Zaffar, G.; Dar, Z.A.; Mehfuza, H. A review on oat (Avena sativa L.) as a dual-purpose crop. Sci. Res. Essays 2014, 9, 52–59. [Google Scholar] [CrossRef]
- Robert, W.W. The Oat Crop: Production and Utilization; Springer: Dordrecht, The Netherlands, 1995; 584p. [Google Scholar] [CrossRef]
- Zhou, Q.; Gou, X.; Tian, L.; Chen, Y.; Gao, S.; Bai, W.; Zhang, W. Performances of early and late maturing oat varieties in cold regions. Chin. Sci. Bull. 2018, 63, 1722–1730. [Google Scholar] [CrossRef]
- Diao, X. Production and genetic improvement of minor cereals in China. Crop J. 2017, 5, 103–114. [Google Scholar] [CrossRef]
- USDA. World Oats Production, Consumption, and Stocks; US Department of Agriculture: Washington, DC, USA, 2014.
- Li, Y.; Ye, W.; Wang, M.; Yan, X. Climate change and drought: A risk assessment of crop-yield impacts. Clim. Res. 2009, 39, 31–46. [Google Scholar] [CrossRef]
- Stevens, E.J.; Armstrong, K.W.; Bezar, H.J.; Griffin, W.B.; Hampton, J.G. Fodder Oats: A World Overview; Food and Agricultural Organization of the United Nations: Roma, Italy, 2004; pp. 11–18. [Google Scholar]
- Hakala, K.; Jauhiainen, L.; Rajala, A.A.; Jalli, M.; Kujala, M.; Laine, A. Different responses to weather events may change the cultivation balance of spring barley and oats in the future. Field Crop. Res. 2020, 259, 107956. [Google Scholar] [CrossRef]
- Hussain, H.A.; Men, S.; Hussain, S.; Chen, Y.; Ali, S.; Zhang, S.; Liao, C.; Wang, L. Interactive effects of drought and heat stresses on morpho-physiological attributes, yield, nutrient uptake and oxidative status in maize hybrids. Sci. Rep. 2019, 9, 3890. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Ordiz, M.I.; Jaworski, J.G.; Beachy, R.N. Induced accumulation of cuticular waxes enhances drought tolerance in Arabidopsis by changes in development of stomata. Plant Physiol. Bioch. 2011, 49, 1448–1455. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.; Battaglia, M.L. Drought stress impacts on plants and different approaches to alleviate its adverse effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef]
- Yang, Z.; Chi, X.; Guo, F.; Jin, X.; Luo, H.; Hawar, A.; Chen, Y.; Feng, K.; Wang, B.; Qi, J.; et al. SbWRKY30 enhances the drought tolerance of plants and regulates a drought stress-responsive gene, SbRD19, in sorghum. J. Plant Physiol. 2020, 246, 153142. [Google Scholar] [CrossRef]
- Lenoir, I.; Fontaine, J.; Sahraoui, A.L.H. Arbuscular mycorrhizal fungal responses to abiotic stresses: A review. Phytochemistry 2016, 123, 4–15. [Google Scholar] [CrossRef]
- Begum, N.; Qin, C.; Ahanger, M.A.; Raza, S.; Khan, M.I.; Ashraf, M.; Ahmed, N.; Zhang, L. Role of arbuscular mycorrhizal fungi in plant growth regulation: Implications in abiotic stress tolerance. Front. Plant Sci. 2019, 10, 1068. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E. Mycorrhizal Symbiosis. Soil Sci. 2008, 3, 273–281. [Google Scholar] [CrossRef]
- Atul-Nayyar, A.; Hamel, C.; Hanson, K.; Germida, J. The arbuscular mycorrhizal symbiosis links N mineralization to plant demand. Mycorrhiza 2009, 19, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Kakouridis, A.; Hagen, J.A.; Kan, M.P.; Mambelli, S.; Feldman, L.J.; Herman, D.J.; Weber, P.K.; Pett-ridge, J.; Firestone, M.K. Routes to roots: Direct evidence of water transport by arbuscular mycorrhizal fungi to host plants. New Phytol. 2022, 236, 210–221. [Google Scholar] [CrossRef]
- Wang, W.; Shi, J.; Xie, Q.; Jiang, Y.; Yu, N.; Wang, E. Nutrient exchange and regulation in arbuscular mycorrhizal symbiosis. Mol. Plant 2017, 10, 1147–1158. [Google Scholar] [CrossRef]
- An, J.; Zeng, T.; Ji, C.; de Graaf, S.; Zheng, Z.; Xiao, T.T.; Limpens, E.; Pan, Z.A. Medicago truncatula SWEET transporter implicated in arbuscule maintenance during arbuscular mycorrhizal symbiosis. New Phytol. 2019, 224, 396–408. [Google Scholar] [CrossRef]
- Tang, H.; Hassan, M.U.; Feng, L.; Nawaz, M.; Shah, A.N.; Qari, S.H.; Liu, Y.; Miao, J. The critical role of arbuscular mycorrhizal fungi to improve drought tolerance and nitrogen use efficiency in crops. Front. Plant Sci. 2022, 13, 919166. [Google Scholar] [CrossRef]
- Li, L.; Zhang, H.; Tang, M.; Chen, H. Nutrient uptake and distribution in mycorrhizal cuttings of Populus× canadensis ‘Neva’under drought stress. J. Soil Sci. Plant Nutr. 2021, 21, 2310–2324. [Google Scholar] [CrossRef]
- Zhou, Q.; Ravnskov, S.; Jiang, D.; Wollenweber, B. Changes in carbon and nitrogen allocation, growth and grain yield induced by arbuscular mycorrhizal fungi in wheat (Triticum aestivum L.) subjected to a period of water deficit. Plant Growth Regul. 2015, 75, 751–760. [Google Scholar] [CrossRef]
- Metwally, A.; Azooz, M.; Nafady, N.; El-Enany, A. Arbuscual mycorrhizal symbiosis alleviates drought stress imposed on wheat plants (Triticum aestivum L.). Appl. Ecol. Environ. Res. 2019, 17, 13713–13727. [Google Scholar] [CrossRef]
- Ratajczak, K.; Sulewska, H.; Błaszczyk, L.; Basińska-Barczak, A.; Mikołajczak, K.; Salamon, S.; Szymańska, G.; Dryjański, L. Growth and Photosynthetic Activity of Selected Spelt Varieties (Triticum aestivum ssp. spelta L.) Cultivated under Drought Conditions with Different Endophytic Core Microbiomes. Int. J. Mol. Sci. 2020, 21, 7987. [Google Scholar] [CrossRef]
- Chen, Y.Y.; Cheng, Y.H.; Jia, X.X. Effects of arbuscular mycorrhizal fungi on the growth and zinc uptake of trifoliate orange (Poncirus trifoliata) seedlings grown in low-zinc soil. J. Plant Nutr. 2017, 40, 324–331. [Google Scholar] [CrossRef]
- Abdel, L.; Arafat, A.H.; He, C. Arbuscular mycorrhizal influence on growth, photosynthetic pigments, osmotic adjustment and oxidative stress in tomato plants subjected to low temperature stress. Acta Physiol. Plant. 2011, 33, 1217–1225. [Google Scholar] [CrossRef]
- Noshad, M.H.; Chavoshi, E.; Mosaddeghi, M.R.; Dorostkar, V.; Hosseini, F. Growth and nutrient uptake of haloxylon and atriplex as affected by mycorrhizal symbiosis under combined drought and salinity stresses. Can. J. Soil Sci. 2022, 103, 305–317. [Google Scholar] [CrossRef]
- Duc, N.H.; Csintalan, Z.; Posta, K. Arbuscular mycorrhizal fungi mitigate negative effects of combined drought and heat stress on tomato plants. Plant Physiol. Bioch. 2018, 132, 297–307. [Google Scholar] [CrossRef]
- Asadollahi, M.; Iranbakhsh, A.; Ahmadvand, R.; Ebadi, M.; Mehregan, I. Synergetic effect of water deficit and arbuscular mycorrhizal symbiosis on the expression of aquaporins in wheat (Triticum aestivum L.) roots: Insights from NGS RNA-sequencing. Physiol. Mol. Biol. Plants 2023, 29, 195–208. [Google Scholar] [CrossRef]
- Tisarum, R.; Theerawitaya, C.; Samphumphuang, T.; Phisalaphong, M.; Singh, H.P.; Cha-Um, S. Promoting water deficit tolerance and anthocyanin fortification in pigmented rice cultivar (Oryza sativa L. subsp. indica) using arbuscular mycorrhizal fungi inoculation. Physiol. Mol. Biol. Plants 2019, 25, 821–835. [Google Scholar] [CrossRef]
- Phillips, J.M. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- McGonigle, T.P.; Miller, M.H.; Evans, D.G.; Fairchild, G.L.; Swan, J.A. A new method which gives an objective measure of colonization of roots by vesicular-arbuscular mycorrhizal fung. New Phytol. 1990, 115, 495–501. [Google Scholar] [CrossRef]
- Van Der Heijden, M.G.A. Arbuscular mycorrhizal fungi as a determinant of plant diversity: In search of underlying mechanisms and general principles. In Mycorrhizal Ecology; Springer: Berlin/Heidelberg, Germany, 2002; pp. 243–265. [Google Scholar] [CrossRef]
- Kong, Z.; Glick, B.R.; Duan, J.; Ding, S.; Tian, J.; McConkey, B.J.; Wei, G. Effects of 1-aminocyclopropane-1-carboxylate (ACC) deaminase-overproducing Sinorhizobium meliloti on plant growth and copper tolerance of Medicago lupulina. Plant Soil 2015, 391, 383–398. [Google Scholar] [CrossRef]
- Sterges, A.J.; Hardin, L.J.; MacIntire, W.H. A modification of the official micro method for the determination of phosphorus content of plant tissue. J. Assoc. Off. Agric. Chem. 1950, 33, 114–121. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, H.G. Determination of Nine Mineral Elements in Hulless Barley by Ultraviolet Spectrophotometry and Flame Atomic Absorption Spectrometry. Spectrosc. Spect. Anal. 2010, 30, 1126–1129. [Google Scholar] [CrossRef]
- Moradi Tarnabi, Z.; Iranbakhsh, A.; Mehregan, I.; Ahmadvand, R. Impact of arbuscular mycorrhizal fungi (AMF) on gene expression of some cell wall and membrane elements of wheat (Triticum aestivum L.) under water deficit using transcriptome analysis. Physiol. Mol. Biol. Plants 2020, 26, 143–162. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Lv, Y.F.; Li, Y.; Li, L.; Jia, J.Q.; Feng, M.C.; Fahad, S.; Zhang, M.J. Inoculation with Rhizophagus intraradices Confers Drought Stress Tolerance in Oat by Improving Nitrogen and Phosphorus Nutrition. J. Soil Sci. Plant Nutr. 2023, 23, 2039–2052. [Google Scholar] [CrossRef]
- Nacoon, S.; Ekprasert, J.; Riddech, N.; Mongkolthanaruk, W.; Jogloy, S.; Vorasoot, N.; Cooper Julia Boonlue, S. Growth enhancement of sunchoke by arbuscular mycorrhizal fungi under drought condition. Rhizosphere 2021, 17, 100308. [Google Scholar] [CrossRef]
- Baum, C.; Toljander, Y.K.; Eckhardt, K.U.; Weih, M. The significance of host-fungus combinations in ectomycorrhizal symbioses for the chemical quality of willow foliage. Plant Soil 2009, 323, 213–224. [Google Scholar] [CrossRef]
- Dąbrowska, G.; Baum, C.; Trejgell, A.; Hrynkiewicz, K. Impact of arbuscular mycorrhizal fungi on the growth and expression of gene encoding stress protein-metallothionein BnMT2 in the non-host crop Brassica napus L. J. Plant Nutr. Soil Sci. 2014, 177, 459–467. [Google Scholar] [CrossRef]
- Liu, C.Y.; Zhang, F.; Zhang, D.J.; Srivastava, A.K.; Wu, Q.S.; Zou, Y.N. Mycorrhiza stimulates root-hair growth and IAA synthesis and transport in trifoliate orange under drought stress. Sci. Rep. 2018, 8, 1978. [Google Scholar] [CrossRef]
- Eltigani, A.; Müller, A.; Ngwene, B.; George, E. Physiological and Morphological Responses of Okra (Abelmoschus esculentus L.) to Rhizoglomus irregulare Inoculation under Ample Water and Drought Stress Conditions Are Cultivar Dependent. Plants 2021, 11, 89. [Google Scholar] [CrossRef]
- Begum, N.; Ahanger, M.A.; Zhang, L. AMF inoculation and phosphorus supplementation alleviates drought induced growth and photosynthetic decline in Nicotiana tabacum by up-regulating antioxidant metabolism and osmolyte accumulation Science Direct. Environ. Exp. Bot. 2020, 176, 104088. [Google Scholar] [CrossRef]
- Ruiz-Lozano, J.M.; Azcón, R. Mycorrhizal colonization and drought stress as factors affecting nitrate reductase activity in lettuce plants. Agric. Ecosyst. Environ. 1996, 60, 175–181. [Google Scholar] [CrossRef]
- Hoang, D.T.T.; Rashtbari, M.; Anh, L.T.; Wang, S.; Tu, D.T.; Hiep, N.V.; Razavi, B.S. Mutualistic interaction between arbuscular mycorrhiza fungi and soybean roots enhances drought resistant through regulating glucose exudation and rhizosphere expansion. Soil Biol. Biochem. 2022, 171, 108728. [Google Scholar] [CrossRef]
- Dong, H.; Kong, X.; Li, W.; Tang, W.; Zhang, D. Effects of plant density and nitrogen and potassium fertilization on cotton yield and uptake of major nutrients in two fields with varying fertility. Field Crop. Res. 2010, 119, 106–113. [Google Scholar] [CrossRef]
- Aksu, G.; Altay, H. The Effects of potassium applications on srought stress in sugar beet. part ii. plant nutrition content. J. Sci. Perspect. 2020, 4, 203–216. [Google Scholar] [CrossRef]
- Markus, K. Water Relations and Nutrient Uptake. In The Science of Grapevines; Academic Press: Cambridge, MA, USA, 2010; pp. 85–105. [Google Scholar] [CrossRef]
- Zhang, H.; Wei, S.; Hu, W.; Xiao, L.; Tang, M. Arbuscular mycorrhizal fungus Rhizophagus irregularis increased potassium content and expression of genes encoding potassium channels in Lycium barbarum. Front. Plant Sci. 2017, 8, 440. [Google Scholar] [CrossRef]
- Lorella, N.; Moscatiello, R.; Genre, A.; Novero, M.; Baldan, B.; Bonfante, P.; Mariani, P. The arbuscular mycorrhizal fungus Glomus intraradices induces intracellular calcium changes in soybean cells. Caryologia 2007, 60, 137–140. [Google Scholar] [CrossRef]
- Mbengue, M.D.; Hervé, C.; Debellé, F. Nod factor signaling in symbiotic nodulation. Adv. Bot. Res. 2020, 94, 1–39. [Google Scholar] [CrossRef]
- Khan, A.; Anwar, Y.; Hasan, M.M.; Iqbal, A.; Ali, M.; Alharby, H.F.; Hakeem, K.R.; Hasanuzzaman, M. Attenuation of Drought Stress in Brassica Seedlings with Exogenous Application of Ca2+ and H2O2. Plants 2017, 6, 20. [Google Scholar] [CrossRef]
- Liu, J.; Fang, L.; Pei, W.; Li, F.; Zhao, J. Effects of magnesium application on the arbuscular mycorrhizal symbiosis in tomato. Symbiosis 2023, 89, 73–82. [Google Scholar] [CrossRef]
- Lopes, J.I.; Arrobas, M.; Brito, C.; Gonçalves, A.; Silva, E.; Martins, S.; Raimundo, S.; Rodrigues, M.Â.; Correia, C.M. Mycorrhizal Fungi were More Effective than Zeolites in Increasing the Growth of Non-Irrigated Young Olive Trees. Sustainability 2020, 12, 10630. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Part | Treatment | Soluble Sugar | Soluble Protein | MDA |
|---|---|---|---|---|
| Shoots | Drought | 10.63 ** | 6.46 ** | 12.13 *** |
| AMF | 110.97 *** | 19.08 *** | 8.33 ** | |
| Drought × AMF | 16.26 *** | 10.45 *** | 2.28 ns | |
| Roots | Drought | 1.94 ns | 2.89 ns | 0.54 ns |
| AMF | 6.34 ** | 0.13 ns | 2.24 ns | |
| Drought × AMF | 11.3 *** | 9.09 *** | 0.558 ns |
| Part | Treatment | Soluble Sugar (mg/g) | Soluble Protein (mg/g) | MDA (nmol/g) |
|---|---|---|---|---|
| Leaves | CK-NM | 10.78 ± 0.72 cd | 17.69 ± 0.52 cd | 8.9 ± 0.36 c |
| CK-R.i | 12.88 ± 1.59 bc | 18.06 ± 1.2 cd | 8.67 ± 0.34 c | |
| CK-F.m | 8.43 ± 0.7 de | 16.33 ± 0.35 de | 8.95 ± 0.37 c | |
| MD-NM | 14.58 ± 0.65 b | 18.49 ± 0.78 bc | 11.26 ± 0.54 ab | |
| MD-R.i | 20.26 ± 0.42 a | 20.49 ± 0.68 ab | 9.95 ± 0.91 bc | |
| MD-F.m | 6.24 ± 0.28 e | 16.67 ± 0.2 cde | 8.71 ± 0.51 c | |
| SD-NM | 8.88 ± 0.62 d | 15.35 ± 0.76 e | 12.22 ± 0.35 a | |
| SD-R.i | 20.06 ± 1.27 a | 22.17 ± 0.44 a | 10.27 ± 0.51 bc | |
| SD-F.m | 9.27 ± 0.47 d | 20.18 ± 0.23 ab | 9.97 ± 0.18 bc | |
| Roots | CK-NM | 5.95 ± 0.78 ab | 8.36 ± 0.91 ab | 13.5 ± 0.3 a |
| CK-R.i | 3.83 ± 0.16 c | 4.77 ± 0.85 c | 8.27 ± 0.56 ef | |
| CK-F.m | 4.65 ± 0.35 abc | 8.89 ± 1.07 ab | 11.96 ± 0.84 abc | |
| MD-NM | 4.46 ± 0.21 bc | 6.53 ± 0.37 bc | 12.66 ± 0.3 ab | |
| MD-R.i | 6.09 ± 0.57 a | 11.7 ± 1.94 a | 10.84 ± 1.04 bcd | |
| MD-F.m | 1.69 ± 0.1 d | 4.17 ± 0.57 c | 7.8 ± 0.79 f | |
| SD-NM | 4.34 ± 0.72 c | 9.01 ± 1.3 ab | 10.17 ± 0.61 cde | |
| SD-R.i | 4.63 ± 0.49 abc | 8.28 ± 0.65 ab | 9.63 ± 0.46 def | |
| SD-F.m | 4.83 ± 0.16 abc | 10.38 ± 1.1 a | 10.3 ± 0.9 cde |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, H.; Jia, Z.; Liu, W.; Wei, X.; Wang, H.; Bao, G.; Li, J.; Zhou, Q. Effects of Arbuscular Mycorrhizal Fungi on Growth and Nutrient Accumulation of Oat under Drought Conditions. Agronomy 2023, 13, 2580. https://doi.org/10.3390/agronomy13102580
Tian H, Jia Z, Liu W, Wei X, Wang H, Bao G, Li J, Zhou Q. Effects of Arbuscular Mycorrhizal Fungi on Growth and Nutrient Accumulation of Oat under Drought Conditions. Agronomy. 2023; 13(10):2580. https://doi.org/10.3390/agronomy13102580
Chicago/Turabian StyleTian, Haoqi, Zhifeng Jia, Wenhui Liu, Xiaoxin Wei, Hui Wang, Gensheng Bao, Jin Li, and Qingping Zhou. 2023. "Effects of Arbuscular Mycorrhizal Fungi on Growth and Nutrient Accumulation of Oat under Drought Conditions" Agronomy 13, no. 10: 2580. https://doi.org/10.3390/agronomy13102580
APA StyleTian, H., Jia, Z., Liu, W., Wei, X., Wang, H., Bao, G., Li, J., & Zhou, Q. (2023). Effects of Arbuscular Mycorrhizal Fungi on Growth and Nutrient Accumulation of Oat under Drought Conditions. Agronomy, 13(10), 2580. https://doi.org/10.3390/agronomy13102580