

Identification of Resistance to the Corn Weevil (Sitophilus zeamais M.) in Mexican Maize Races (Zea mays L.)

 , , ,
, , ,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Bioassays for Resistance Studies
2.2.1. Obligate-Antibiosis Bioassays
2.2.2. Free-Choice Bioassays
2.3. Statistical Analysis
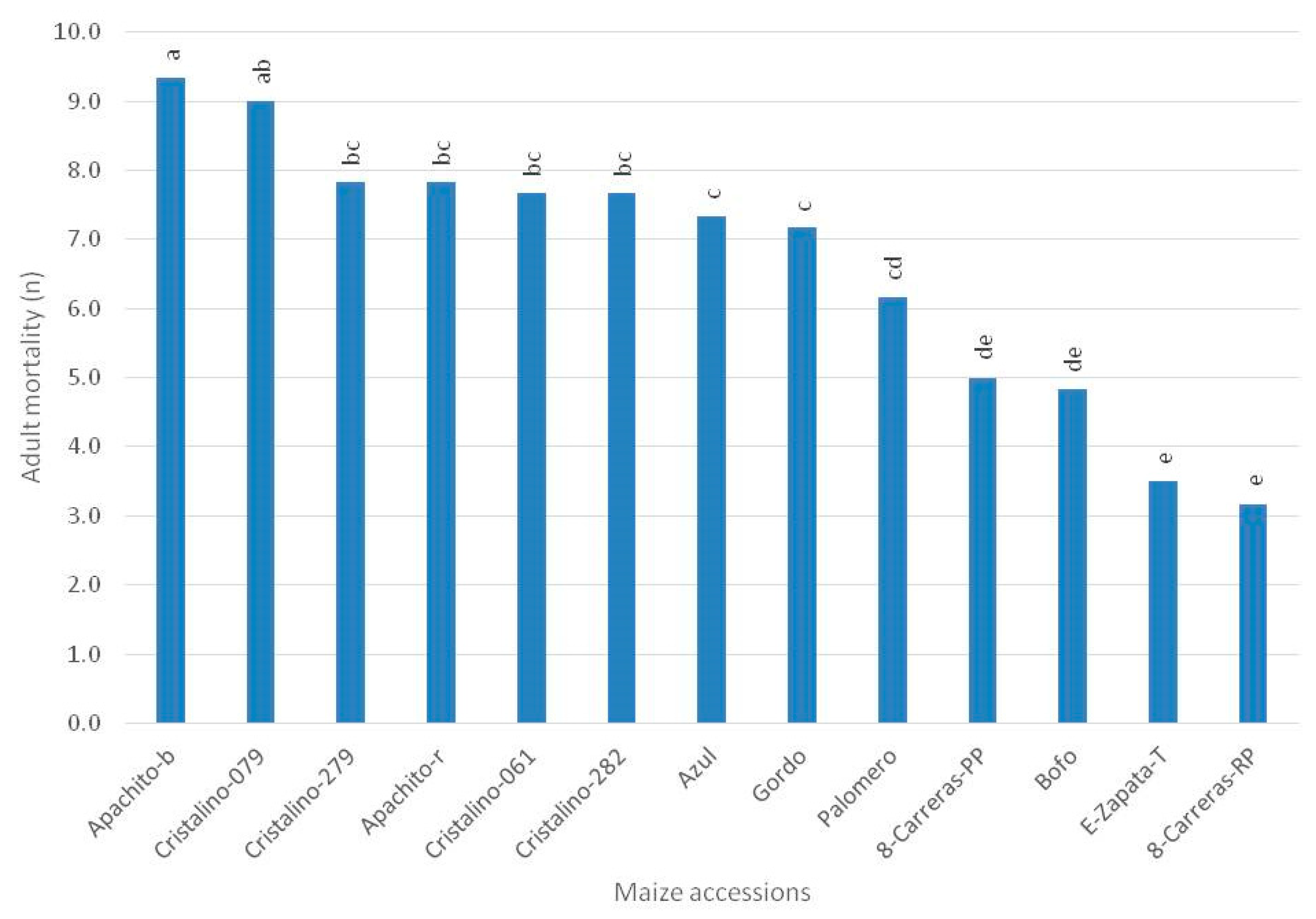
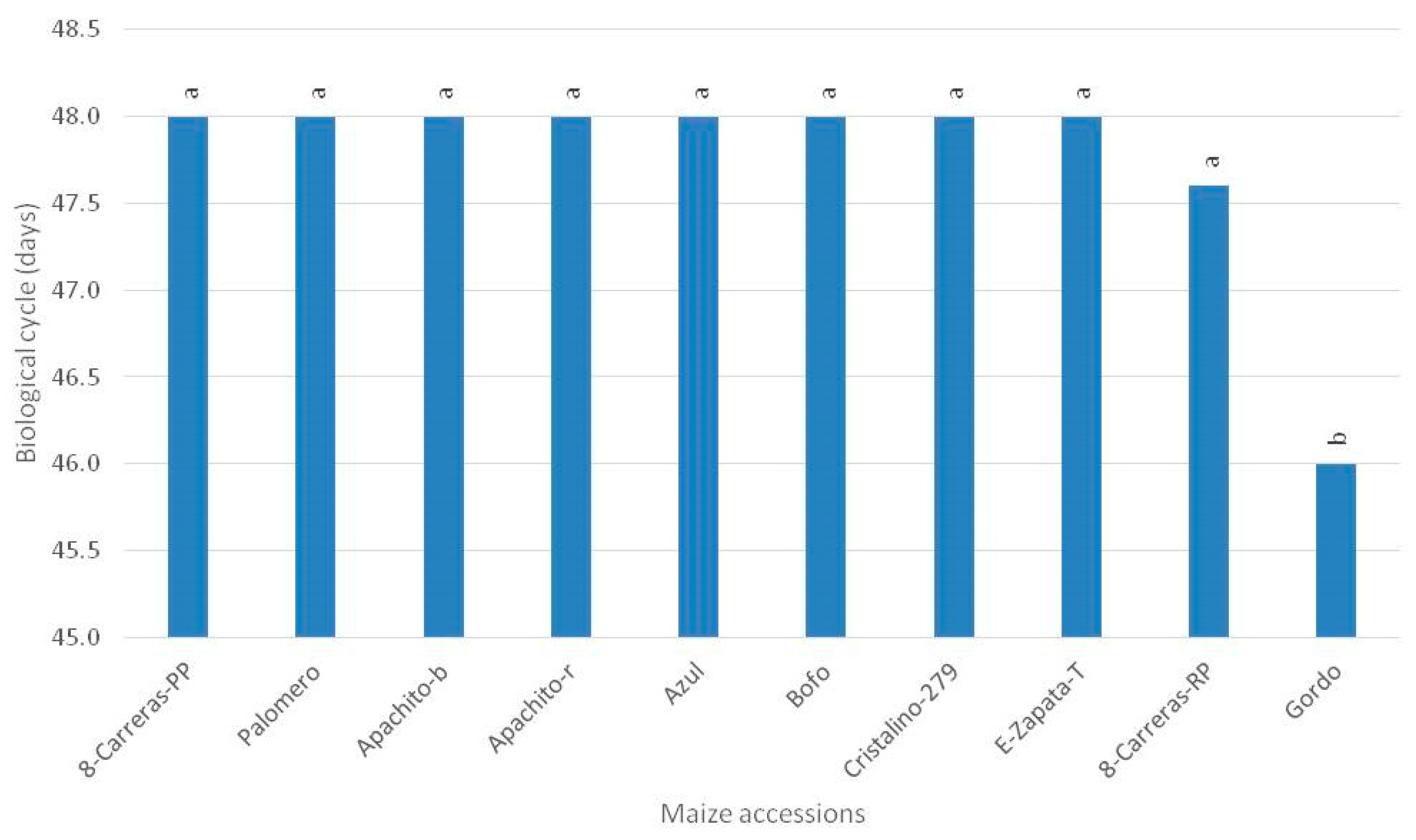
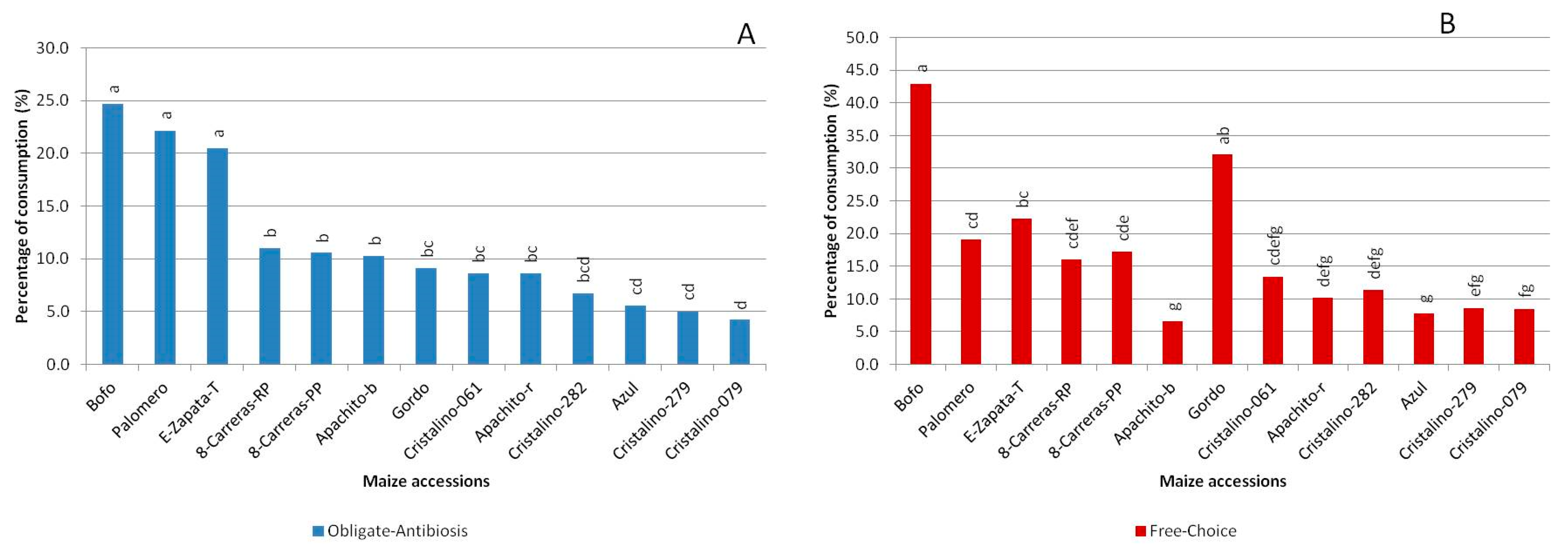
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. Statistical Database. In Food and Agriculture Organization of the United Nations; FAO: Rome, Italy, 2022; Available online: http://faostat.fao.org/ (accessed on 24 June 2022).
- Boxall, R.A. Damage and loss caused by the larger grain borer Prostephanus truncatus. Integr. Pest Manag. Rev. 2002, 7, 105–121. [Google Scholar] [CrossRef]
- Shah, J.; Chaturvedi, R.; Chowdhury, Z.; Venables, B.; Petros, R.A. Signaling by small metabolites in systemic acquired resistance. Plant J. 2014, 79, 645–658. [Google Scholar] [CrossRef]
- García-Lara, S.; Espinosa Carrillo, C.; Bergvinson, D.J. Manual de Plagas en Granos Almacenado y Tecnologías Alternas para su Manejo y Control; CIMMYT: El Batán, Mexico, 2007. [Google Scholar]
- Tefera, T.; Mugo, S.; Likhayo, P. Effects of insect population density and storage time on grain damage and weight loss in maize due to the maize weevil Sitophilus zeamais and the larger grain borer Prostephanus truncatus. Afr. J. Agric. Res. 2011, 6, 2249–2254. [Google Scholar]
- Tavares, W.S.; Costa, M.A.; Cruz, I.; Silveira, R.D.; Serrao, J.E.; Zanuncio, J.C. Selective effects of natural and synthetic insecticides on mortality of Spodoptera frugiperda (Lepidoptera: Noctuidae) and its predator Eriopis connexa (Coleoptera: Coccinellidae). J. Environ. Sci. Health Part B 2010, 45, 557–561. [Google Scholar] [CrossRef]
- Yoza, K.-i.; Imamura, T.; Kramer, K.J.; Morgan, T.D.; Nakamura, S.; Akiyama, K.; Kawasaki, S.; Takaiwa, F.; Ohtsubo, K.i. Avidin expressed in transgenic rice confers resistance to the stored-product insect pests Tribolium confusum and Sitotroga cerealella. Biosci. Biotechnol. Biochem. 2005, 69, 966–971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keneni, G.; Bekele, E.; Getu, E.; Imtiaz, M.; Damte, T.; Mulatu, B.; Dagne, K. Breeding Food Legumes for Resistance to Storage Insect Pests: Potential and Limitations. Sustainability 2011, 3, 1399–1415. [Google Scholar] [CrossRef] [Green Version]
- Jiménez, J.C.; de la Fuente, M.; Ordás, B.; Domínguez, L.E.G.; Malvar, R.A. Resistance categories to Acanthoscelides obtectus (Coleoptera: Bruchidae) in tepary bean (Phaseolus acutifolius), new sources of resistance for dry bean (Phaseolus vulgaris) breeding. Crop Prot. 2017, 98, 255–266. [Google Scholar] [CrossRef]
- Arnason, J.; Baum, B.; Gale, J.; Lambert, J.; Bergvinson, D.; Philogene, B.; Serratos, J.; Mihm, J.; Jewell, D. Variation in resistance of Mexican landraces of maize to maize weevil Sitophilus zeamais, in relation to taxonomic and biochemical parameters. Euphytica 1993, 74, 227–236. [Google Scholar] [CrossRef]
- Abebe, F.; Tefera, T.; Mugo, S.; Beyene, Y.; Vidal, S. Resistance of maize varieties to the maize weevil Sitophilus zeamais (Motsch.) (Coleoptera: Curculionidae). Afr. J. Biotechnol. 2009, 8, 5937–5943. [Google Scholar]
- Serratos, A.; Arnason, J.; Nozzolillo, C.; Lambert, J.; Philogene, B.; Fulcher, G.; Davidson, K.; Peacock, L.; Atkinson, J.; Morand, P. Factors contributing to resistance of exotic maize populations to maize weevil, Sitophilus zeamais. J. Chem. Ecol. 1987, 13, 751–762. [Google Scholar] [CrossRef]
- Kim, S.K.; Kossou, D.K. Responses and genetics of maize germplasm resistant to the maize weevil Sitophilus zeamais Motschulsky in West Africa. J. Stored Prod. Res. 2003, 39, 489–505. [Google Scholar] [CrossRef]
- Muzemu, S.; Chitamba, J.; Goto, S. Screening of stored maize (Zea mays L.) varieties grain for tolerance against maize weevil, Sitophilus zeamais (Motsch.). Int. J. Plant Res. 2013, 3, 17–22. [Google Scholar]
- Tefera, T.; Demissie, G.; Mugo, S.; Beyene, Y. Yield and agronomic performance of maize hybrids resistant to the maize weevil Sitophilus zeamais Motschulsky (Coleoptera: Curculionidae). Crop Prot. 2013, 46, 94–99. [Google Scholar] [CrossRef]
- Mwololo, J.; Mugo, S.; Okori, P.; Tefera, T.; Otim, M.; Munyiri, S. Sources of Resistance to the Maize Weevil Sitophilus zeamais in Tropical Maize; Canadian Center of Science and Education: Toronto, ON, Canada, 2012. [Google Scholar]
- Oloyede-Kamiyo, Q.; Adetumbi, J. Relationship between seed physical traits and maize weevil (Sitophilus zeamais) damage parameters in selected Quality Protein Maize (QPM) varieties. J. Stored Prod. Res. 2017, 73, 42–46. [Google Scholar] [CrossRef]
- Khakata, S.; Nzuve, F.; Chemining’wa, G.; Mwimali, M.; Karanja, J.; Harvey, J.; Mwololo, J. Post-harvest evaluation of selected hybrids to maize weevil Sitophilus zeamais resistance. J. Stored Prod. Postharvest Res. 2018, 9, 16–26. [Google Scholar]
- Nwosu, L.C. Chemical bases for maize grain resistance to infestation and damage by the maize weevil, Sitophilus zeamais Motschulsky. J. Stored Prod. Res. 2016, 69, 41–50. [Google Scholar] [CrossRef]
- Castro-Alvarez, F.F.; William, M.; Bergvinson, D.J.; García-Lara, S. Genetic mapping of QTL for maize weevil resistance in a RIL population of tropical maize. Theor. Appl. Genet. 2015, 128, 411–419. [Google Scholar] [CrossRef]
- García-Lara, S.; Burt, A.J.; Arnason, J.T.; Bergvinson, D.J. QTL mapping of tropical maize grain components associated with maize weevil resistance. Crop Sci. 2010, 50, 815–825. [Google Scholar] [CrossRef]
- Suleiman, R.K.; Rosentrater, K.A.; Bern, C. Evaluation of maize weevils Sitophilus zeamais Motschulsky infestation on seven varieties of maize. J. Stored Prod. Res. 2015, 64, 97–102. [Google Scholar] [CrossRef]
- Cardona, C.; Kornegay, J. Bean germplasm resources for insect resistance. In Global Plant Genetic Resources for Insect-Resistant Crops; CRC Press: Boca Raton, FL, USA, 1999; pp. 85–99. [Google Scholar]
- Sharma, H.C.; Ortiz, R. Host plant resistance to insects: An eco-friendly approach for pest management and environment conservation. J. Environ. Biol. 2002, 23, 111–135. [Google Scholar] [PubMed]
- SAS Institute. Base SAS 9.4 Procedures Guide: Statistical Procedures. Version 9.4; SAS Institute: Cary, NC, USA, 2016. [Google Scholar]
- Butrón, A.; Romay, M.; Ordás, A.; Malvar, R.; Revilla, P. Genetic and environmental factors reducing the incidence of the storage pest Sitotroga cerealella in maize. Entomol. Exp. Appl. 2008, 128, 421–428. [Google Scholar] [CrossRef] [Green Version]
- Ehrlich, P.R.; Raven, P.H. Butterflies and plants: A study in coevolution. Evolution 1964, 18, 586–608. [Google Scholar] [CrossRef]
- Hogenhout, S.A.; Bos, J.I. Effector proteins that modulate plant–insect interactions. Curr. Opin. Plant Biol. 2011, 14, 422–428. [Google Scholar] [CrossRef]
- Jermy, T. Evolution of insect/host plant relationships. Am. Nat. 1984, 124, 609–630. [Google Scholar] [CrossRef]
- Rausher, M.D. Co-evolution and plant resistance to natural enemies. Nature 2001, 411, 857–864. [Google Scholar] [CrossRef]
- Thompson, J.N. Specific hypotheses on the geographic mosaic of coevolution. Am. Nat. 1999, 153, S1–S14. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Casillas, J.M.; Esquivel, G.E. Rendimiento de grano y características agronómicas en germoplasma de maíz de valles altos de México. Rev. Fitotec. Mex. 2004, 27, 27–31. [Google Scholar]
- Suleiman, R.; Williams, D.; Nissen, A.; Bern, C.; Rosentrater, K. Is flint corn naturally resistant to Sitophilus zeamais infestation? J. Stored Prod. Res. 2015, 60, 19–24. [Google Scholar] [CrossRef]
- Ahmed, S.; Faridi, U.; Shahid, M.; Nisar, M. Effect of phenolics in maize grains of different varieties on biology of Angoumois grain moth, Sitotroga cerealella (Oliv.) (Lepidoptera: Gelechiidae). Cereal Res. Commun. 2013, 41, 636–646. [Google Scholar] [CrossRef]
- Jiménez-Galindo, J.C.; Tortosa, M.; Velasco, P.; De La Fuente, M.; Ordás, B.; Malvar, R.A. Inheritance and metabolomics of the resistance of two F 2 populations of Phaseolus spp. to Acanthoscelides obtectus. Arthropod-Plant Interact. 2020, 14, 641–651. [Google Scholar] [CrossRef]
- Nhamucho, E.; Mugo, S.; Gohole, L.; Tefera, T.; Kinyua, M.; Mulima, E. Resistance of selected Mozambican local and improved maize genotypes to maize weevil, Sitophilus zeamais (Motschulsky). J. Stored Prod. Res. 2017, 73, 115–124. [Google Scholar] [CrossRef]
- García-Lara, S.; Bergvinson, D.J.; Burt, A.J.; Ramputh, A.I.; Díaz-Pontones, D.M.; Arnason, J.T. The role of pericarp cell wall components in maize weevil resistance. Crop Sci. 2004, 44, 1546–1552. [Google Scholar] [CrossRef]
- Lale, N.E.S.; Kartay, M.O. Role of physical characteristics of the seed in the resistance of local cultivars of maize to Sitophilus zeamais infestation in storage. Trop. Sci. 2006, 46, 1–3. [Google Scholar] [CrossRef]
- Akpodiete, O.; Lale, N.; Umeozor, O.; Zakka, U. Role of physical characteristics of the seed on the stability of resistance of maize varieties to maize weevil (Sitophilus zeamais Motschulsky). IOSR J. Environ. Sci. Toxicol. Food Technol. Ver. II 2015, 9, 60–66. [Google Scholar]
- García-Lara, S.; Arnason, J.T.; Díaz-Pontones, D.; Gonzalez, E.; Bergvinson, D.J. Soluble peroxidase activity in maize endosperm associated with maize weevil resistance. Crop Sci. 2007, 47, 1125–1130. [Google Scholar] [CrossRef]
- López-Castillo, L.M.; Flores-Rivera, M.F.D.; Winkler, R.; García-Lara, S. Increase of peroxidase activity in tropical maize after recurrent selection to storage pest resistance. J. Stored Prod. Res. 2018, 75, 47–55. [Google Scholar] [CrossRef]
- Saulnier, L.; Thibault, J.F. Ferulic acid and diferulic acids as components of sugar-beet pectins and maize bran heteroxylans. J. Sci. Food Agric. 1999, 79, 396–402. [Google Scholar] [CrossRef]
- Sen, A.; Bergvinson, D.; Miller, S.S.; Atkinson, J.; Fulcher, R.G.; Arnason, J.T. Distribution and microchemical detection of phenolic acids, flavonoids, and phenolic acid amides in maize kernels. J. Agric. Food Chem. 1994, 42, 1879–1883. [Google Scholar] [CrossRef]
- Macías-Estrada, P.; Orozco-González, F.; Castellanos-Pérez, G.; Castillo-Rosales, A.; Ortega-Ortega, A.; Malvar, R.A.; Padilla-Chacón, D.; Jiménez-Galindo, J.C. Sitotroga cerealella-resistant mexican maize races (Zea mays L.), new sources of resistance for commercial maize breeding. Cereal Res. Commun. 2022, 1–12. [Google Scholar] [CrossRef]
- López-Castillo, L.M.; Silva-Fernández, S.E.; Winkler, R.; Bergvinson, D.J.; Arnason, J.T.; García-Lara, S. Postharvest insect resistance in maize. J. Stored Prod. Res. 2018, 77, 66–76. [Google Scholar] [CrossRef]
- Derera, J.; Pixley, K.V.; Giga, D.P.; Makanda, I. Resistance of maize to the maize weevil: III. Grain weight loss assessment and implications for breeding. J. Stored Prod. Res. 2014, 59, 24–35. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | Maize Race | Color | Endosperm Type a |
|---|---|---|---|
| 8-Carreras-PP | 8-Carreras | White | Dent |
| 8-Carreras-RP | 8-Carreras | White | Dent |
| Azul | Azul | Black | Floury |
| Cristalino-061 | Cristalino de Chihuahua | White | Flint |
| Bofo | Bofo | Purple | Floury |
| E-Zapata-T | – | White | Dent |
| Cristalino-279 | Cristalino de Chihuahua | White | Flint |
| Cristalino-282 | Cristalino de Chihuahua | Yellow | Flint |
| Apachito-r | Apachito | Pink | Flint |
| Cristalino-079 | Cristalino de Chihuahua | Yellow | Flint |
| Apachito-b | Apachito | White | Flint |
| Gordo | Gordo | White | Floury |
| Palomero | Palomero de Chihuahua | Yellow | Popcorn |
| Trait. Type of Bioassay | Endosperm Type | ||
|---|---|---|---|
| Dent | Flint | Floury | |
| Adult’s mortality. Obligate-antibiosis | 0.389 c | 0.793 a | 0.644 b |
| Emerged Adults. Obligate-antibiosis | 6.9 a | 2.1 b | 4.2 a |
| Net consumption (g). Free-choice | 0.612 a | 0.216 b | 0.525 a |
| Net consumption (g). Obligate-antibiosis | 0.517 a | 0.198 c | 0.346 b |
| Percentage of consumption (%). Free-choice | 19.9 a | 12.3 b | 24 a |
| Percentage of consumption (%). Obligate-antibiosis | 14.7 a | 9.5 b | 12.1 ab |
| Initial Weight (g) | Adult Mortality (n) | Biological Cycle (days) | Emerged Adult (n) | Net Consumption (g) | Percentage of Consumption (%) | |
|---|---|---|---|---|---|---|
| Initial weight (g) | 0.38 | 0.42 | −0.16 | |||
| Adult mortality (n) | −0.67 * | |||||
| First generation duration (days) | 0.29 | −0.01 | ||||
| Emerged adults (n) | 0.50 | −0.94 * | 0.15 | 0.58 | 0.40 | |
| Net consumption (g) | 0.50 | −0.85 * | 0.26 | 0.95 * | 0.81 * | |
| Percentage of consumption (%) | 0.02 | −0.61 * | 0.09 | 0.82 * | 0.86 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiménez-Galindo, J.C.; Castillo-Rosales, A.; Castellanos-Pérez, G.; Orozco-González, F.; Ortega-Ortega, A.; Padilla-Chacón, D.; Butrón, A.; Revilla, P.; Malvar, R.A. Identification of Resistance to the Corn Weevil (Sitophilus zeamais M.) in Mexican Maize Races (Zea mays L.). Agronomy 2023, 13, 312. https://doi.org/10.3390/agronomy13020312
Jiménez-Galindo JC, Castillo-Rosales A, Castellanos-Pérez G, Orozco-González F, Ortega-Ortega A, Padilla-Chacón D, Butrón A, Revilla P, Malvar RA. Identification of Resistance to the Corn Weevil (Sitophilus zeamais M.) in Mexican Maize Races (Zea mays L.). Agronomy. 2023; 13(2):312. https://doi.org/10.3390/agronomy13020312
Chicago/Turabian StyleJiménez-Galindo, José Cruz, Adán Castillo-Rosales, Gloria Castellanos-Pérez, Francisco Orozco-González, Adalberto Ortega-Ortega, Daniel Padilla-Chacón, Ana Butrón, Pedro Revilla, and Rosa Ana Malvar. 2023. "Identification of Resistance to the Corn Weevil (Sitophilus zeamais M.) in Mexican Maize Races (Zea mays L.)" Agronomy 13, no. 2: 312. https://doi.org/10.3390/agronomy13020312
APA StyleJiménez-Galindo, J. C., Castillo-Rosales, A., Castellanos-Pérez, G., Orozco-González, F., Ortega-Ortega, A., Padilla-Chacón, D., Butrón, A., Revilla, P., & Malvar, R. A. (2023). Identification of Resistance to the Corn Weevil (Sitophilus zeamais M.) in Mexican Maize Races (Zea mays L.). Agronomy, 13(2), 312. https://doi.org/10.3390/agronomy13020312