

Assessment of Resistance Mechanisms to Fall Armyworm, Spodoptera frugiperda in Tropical Maize Inbred Lines

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. FAW Larvae Colony
2.3. Plants Used in the Laboratory Experiment
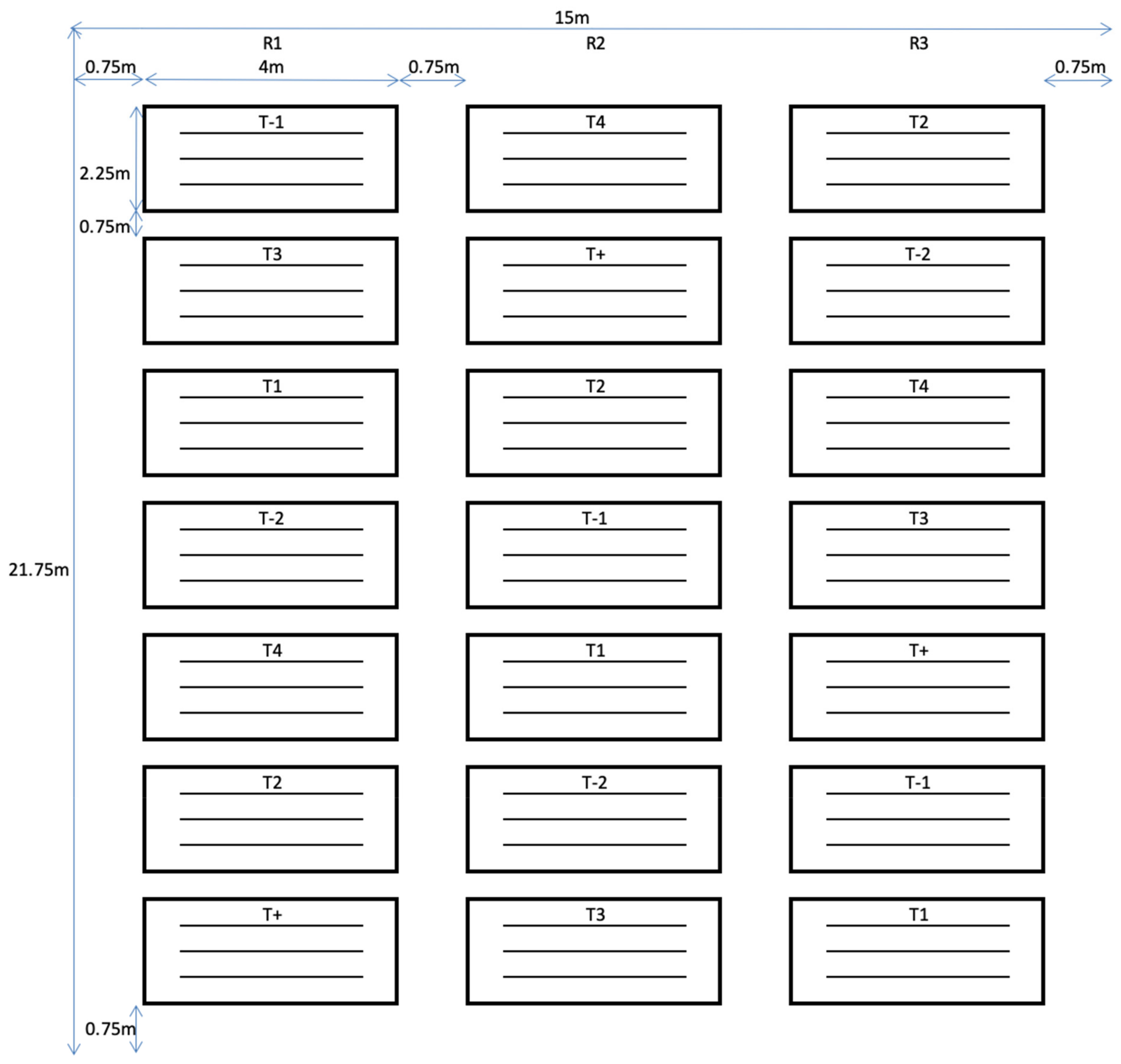
2.4. Plants Used in the Net House Experiment
2.5. Antibiosis Assessment under Laboratory Conditions
2.6. Antibiosis Assessment under Net House Conditions
2.7. Antixenosis Assessment under Laboratory Conditions
2.8. Data Analyses
- − For antibiosis assessments
- − For antixenosis assessments
3. Results
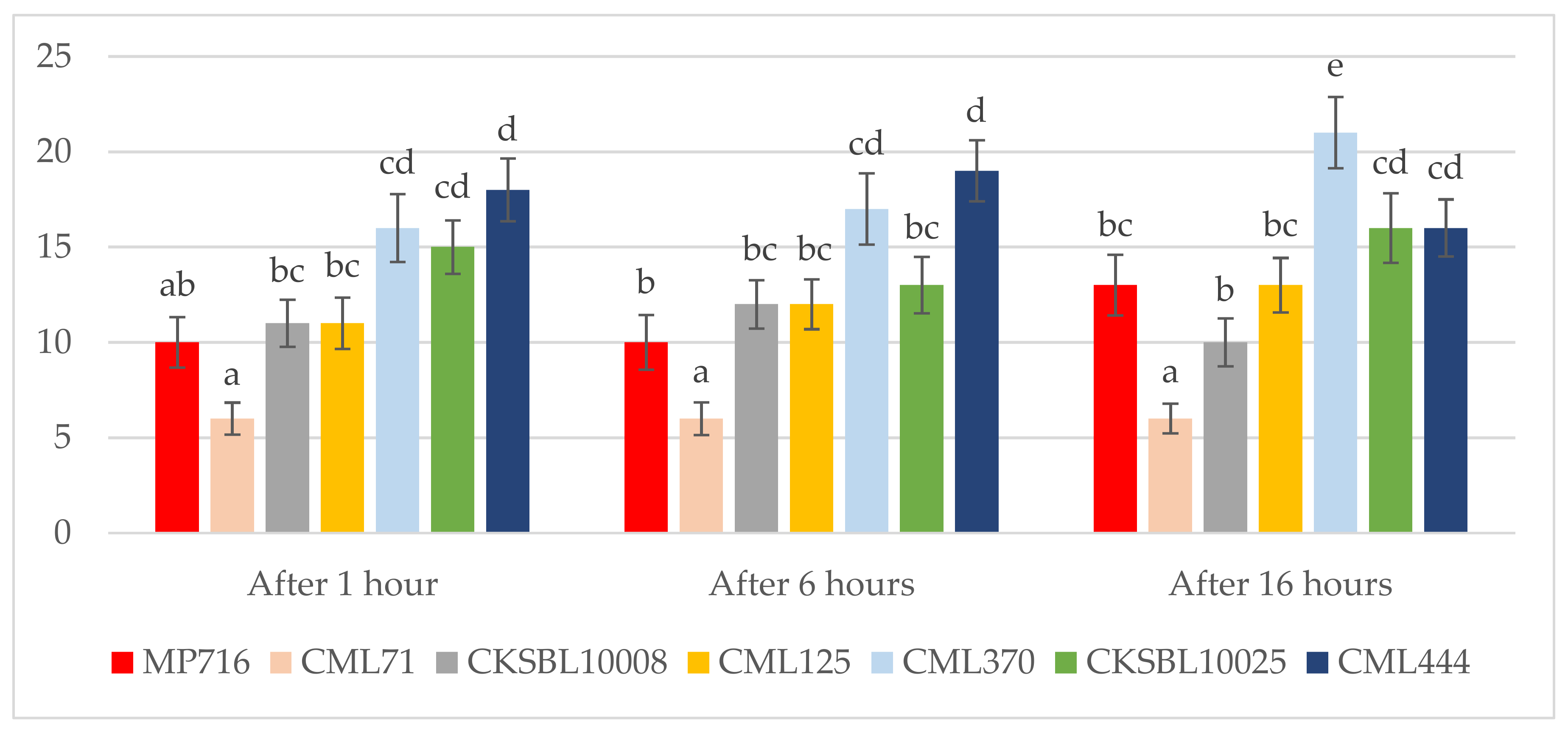
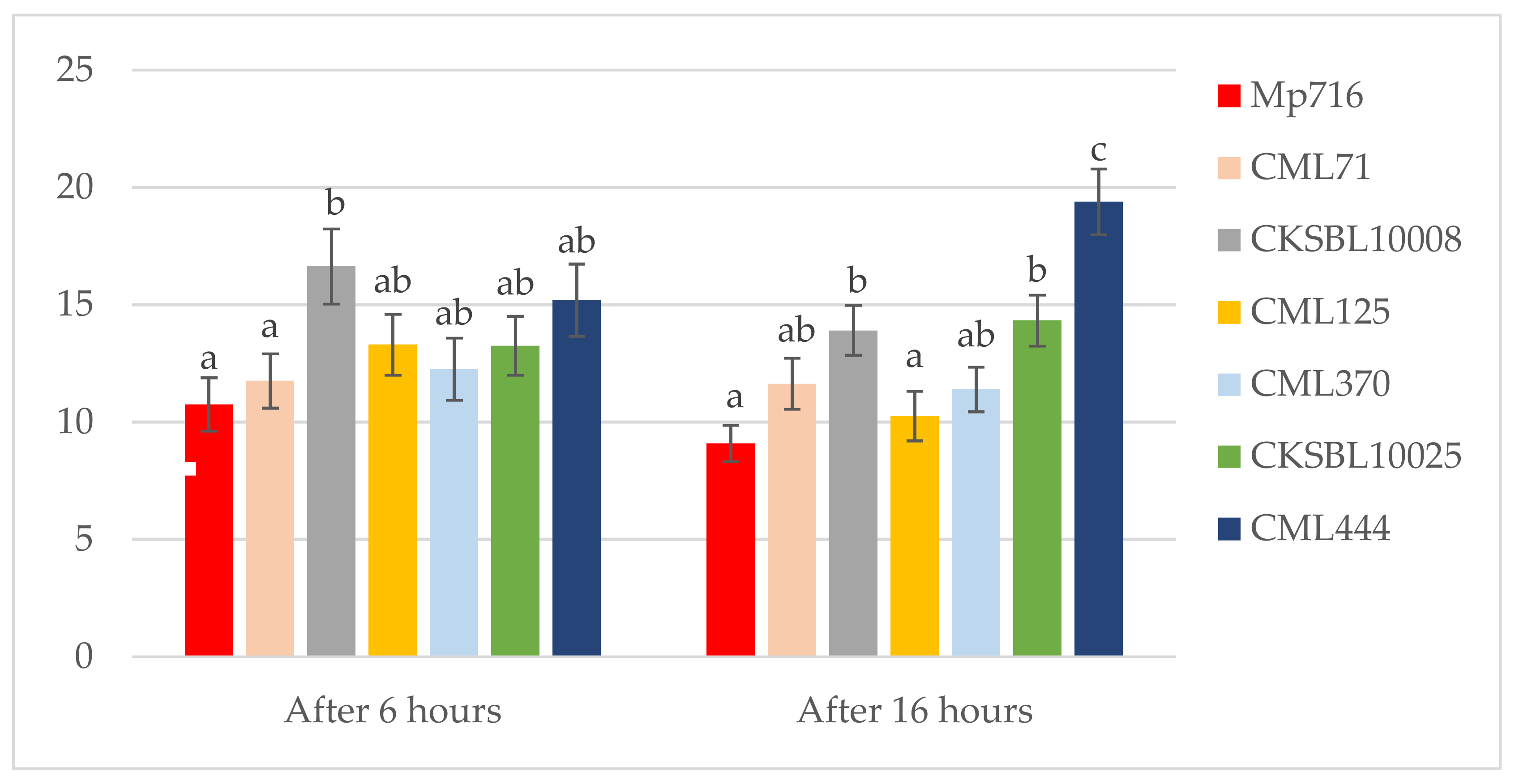
3.1. Antibiosis Assessments
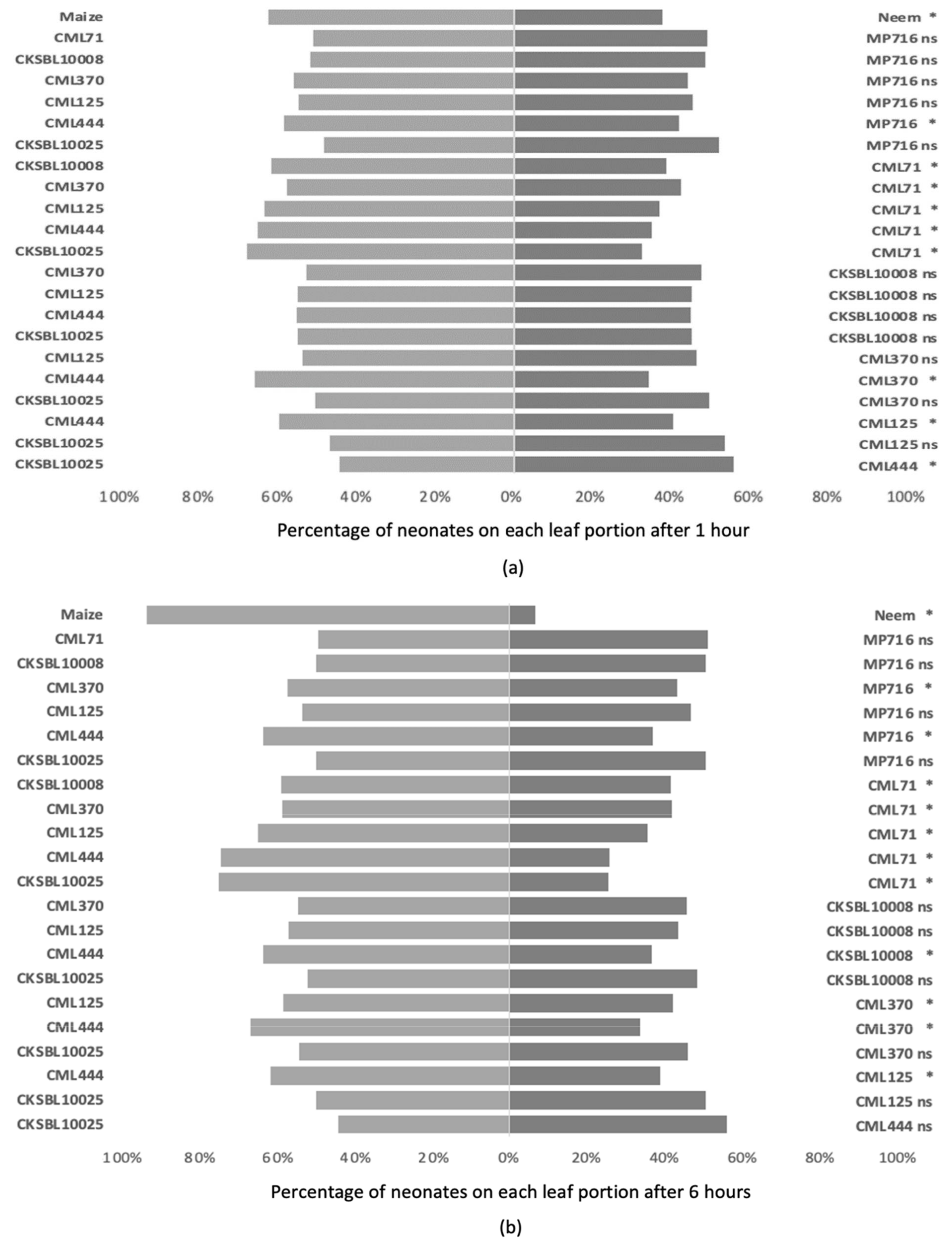
3.2. Antixenosis Assessments on Leaves
3.2.1. Binary Choice Tests
3.2.2. Multiple Choice Tests
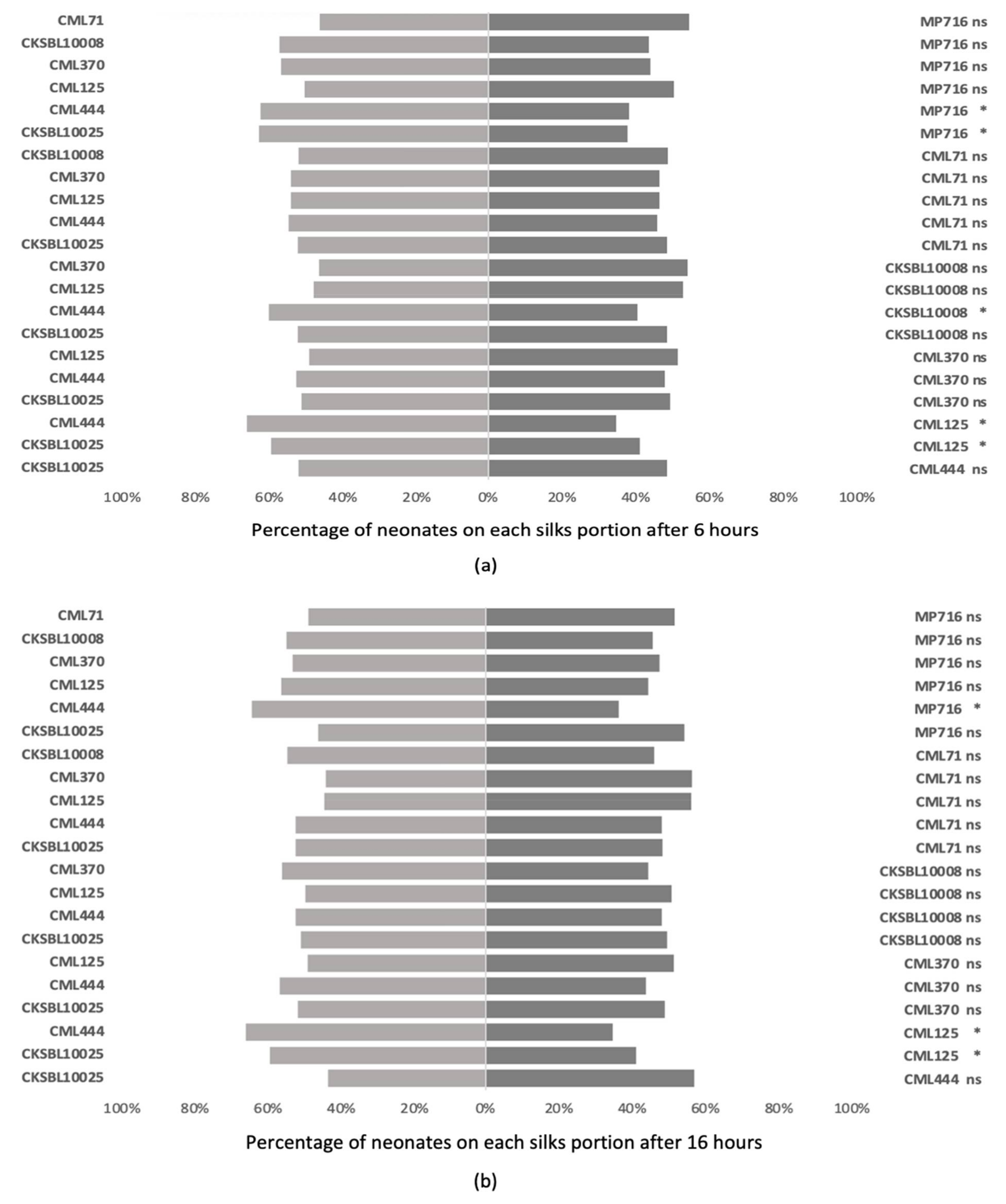
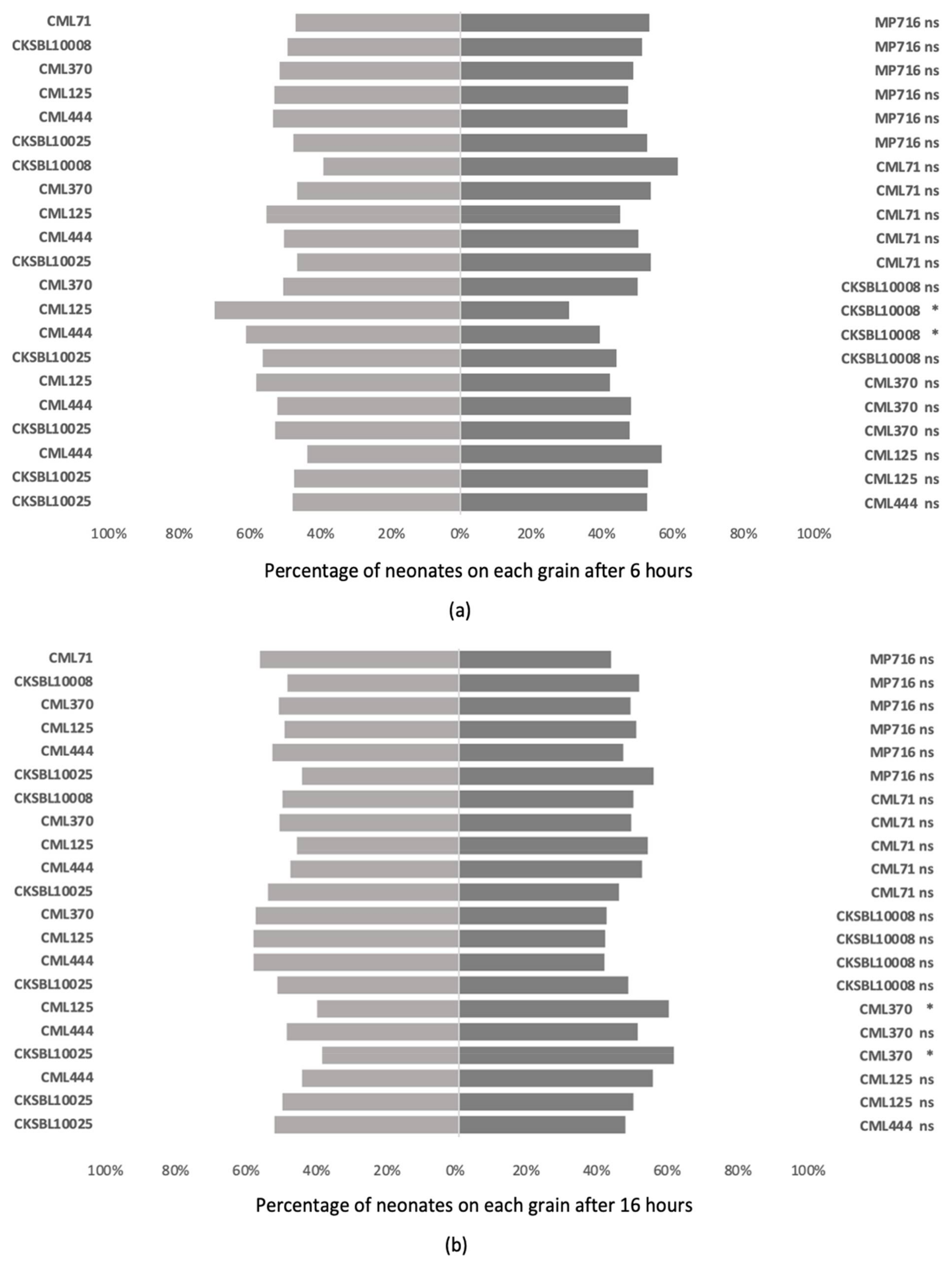
3.3. Antixenosis Assessments on Maize Silks and Grains
3.3.1. Binary Choice Tests
3.3.2. Multiple Choice Tests
4. Discussion
5. Conclusions and Perspectives
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Erenstein, O.; Jaleta, M.; Sonder, K.; Mottaleb, K.; Prasanna, B.M. Global maize production, consumption, and trade: Trends and R&D implications. Food Secur. 2022, 14, 1295–1319. [Google Scholar] [CrossRef]
- Goergen, G.; Kumar, P.L.; Sankung, S.B.; Togola, A.; Tamò, M. First report of outbreaks of the fall armyworm Spodoptera frugiperda (J E Smith) (Lepidoptera, Noctuidae), a new alien invasive pest in West and Central Africa. PLoS ONE 2016, 11, e0165632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Day, R.; Abrahams, P.; Bateman, M.; Beale, T.; Clottey, V.; Cock, M.; Colmenarez, Y.; Corniani, N.; Early, R.; Godwin, J.; et al. Fall armyworm: Impacts and implications for Africa. Outlooks on pest management. Outlooks Pest Manag. 2017, 28, 196–201. [Google Scholar] [CrossRef] [Green Version]
- Food and Agriculture Organization (FAO). Fall Armyworm Threatens Food Security and Livelihoods Across Africa 2018, Rome. Available online: http://www.fao.org/3/i8503en/I8503EN.pdf (accessed on 13 December 2022).
- Mihm, J.A. Insect Resistant Maize-Recent Advances and Utilization; Mihm, J.A., Ed.; CIMMYT: El Batan, Mexico, 1997; p. 304. [Google Scholar]
- Prasanna, B.M.; Bruce, A.; Beyene, Y.; Makumbi, D.; Gowda, M.; Asim, M.; Martinelli, S.; Graham, P.H.; Srinivas, P. Host Plant Resistance for Fall Armyworm Management in Maize: Relevance, Status and Prospects in Africa and Asia. Theor. Appl. Genet. 2022, 135, 3897–3916. [Google Scholar] [CrossRef] [PubMed]
- Davis, F.M.; Ng, S.S.; Williams, W.P. Mechanisms of resistance in corn to leaf feeding by southwestern corn borer and European corn borer (Lepidoptera: Pyralidae). J. Econ. Entomol. 1989, 82, 919–922. [Google Scholar] [CrossRef]
- Wiseman, B.R.; Williams, W.P.; Davis, F.M. Fall armyworm: Resistance mechanisms in selected corn genotypes. J. Econ. Entomol. 1981, 74, 622–624. [Google Scholar] [CrossRef]
- Williams, W.P.; Davis, F.M. Registration of maize germplasm line Mp716. Crop Sci. 2002, 42, 671–672. [Google Scholar] [CrossRef]
- Otim, M.H.; Fiaboe, K.; Akello, J.; Mudde, B.; Obonyom, A.T.; Bruce, A.Y.; Opio, W.A.; Chinwada, P.; Hailu, G.; Paparu, P. Managing a transboundary pest: The fall armyworm on maize in Africa. In Moths and Caterpillars; IntechOpen: London, UK, 2021; pp. 1–16. [Google Scholar] [CrossRef]
- Prasanna, B.M.; Huesing, J.E.; Eddy, R.; Peschke, V.M. (Eds.) Fall Armyworm in Africa: A Guide for Integrated Pest Management, 1st ed.; CIMMYT: Mexico City, Mexico, 2018; p. 109. [Google Scholar]
- Painter, R.H. Insect Resistance in Crop Plants; The University Press of Kansas: Lawrence, KS, USA, 1951; p. 520. [Google Scholar]
- Pioneer Agronomy Sciences. Staging Corn Growth. 2022. Available online: https://www.pioneer.com/us/agronomy/staging_corn_growth.html (accessed on 13 December 2022).
- Santos, L.M.; Redaelli, L.R.; Diefenbach, L.M.G.; Efrom, C.F.S. Larval and pupal stage of Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) in sweet and field corn genotypes. Braz. J. Biol. 2003, 63, 627–633. [Google Scholar] [CrossRef] [Green Version]
- Chapman, J.W.; Williams, T.; Escribano, A.; Caballero, P.; Cave, R.D.; Goulson, D. Age-related cannibalism and horizontal transmission of a nuclear polyhedrosis virus in larval Spodoptera frugiperda. Ecol. Entomol. 1999, 24, 268–275. [Google Scholar] [CrossRef]
- Ojeda-Avila, T.; Woods, H.A.; Raguso, R.A. Effects of dietary variation on growth, composition and maturation of Manduca sexta (Sphingidae: Lepidoptera). J. Insect Physiol. 2003, 49, 293–306. [Google Scholar] [CrossRef]
- Rharrabe, K.; Jacquin-Joly, E.; Marion-Poll, F. Electrophysiological and behavioral responses of Spodoptera littoralis caterpillars to attractive and repellent plant volatiles. Front. Ecol. Evol. 2014, 2, 5. [Google Scholar] [CrossRef] [Green Version]
- de Fouchier, A.; Sun, X.; Caballero-Vidal, G.; Travaillard, S.; Jacquin-Joly, E.; Montagné, N. Behavioral Effect of Plant Volatiles Binding to Spodoptera littoralis Larval Odorant Receptors. Front. Behav. Neurosci. 2018, 12, 264. [Google Scholar] [CrossRef] [Green Version]
- Morales, X.C.; Tamiru, A.; Sobhy, I.S.; Bruce, T.J.A.; Midega, C.A.O.; Khan, Z. Evaluation of African maize cultivars for resistance to fall armyworm Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) larvae. Plants 2021, 10, 392. [Google Scholar]
- Hothorn, T.; Bretz, F.; Westfall, P.; Heiberger, R.M.; Schuetzenmeister, A.; Scheibe, S. Package ‘Multcomp’. 2021. Available online: https://cran.r-project.org/web/packages/multcomp/multcomp.pdf (accessed on 13 December 2022).
- Dinno, A. Dunn’s Test of Multiple Comparisons Using Rank Sums. Package Dunn.test. 2017. Available online: https://cran.r-project.org/web/packages/dunn.test/dunn.test.pdf (accessed on 4 January 2023).
- R Core Team. R: A Language and Environment for Statistical Computing. 2018. Available online: https://www.R-project.org/ (accessed on 13 December 2022).
- Chang, Y.M.; Luthe, D.S.; Davis, F.M.; Williams, W.P. Influence of whorl region from resistant and susceptible corn genotypes on fall armyworms (Lepidopera: Noctuidae) growth and development. J. Econ. Entomol. 2000, 93, 477–483. [Google Scholar] [CrossRef]
- Chen, Y.; Ni, X.; Buntin, G.D. Physiological, nutritional, and biochemical bases of corn resistance to foliage-feeding fall armyworm. J. Chem. Ecol. 2009, 35, 297–306. [Google Scholar] [CrossRef]
- Scott, G.E.; Davis, F.M. Registration of MpSWCB-4 population of maize (Reg. No. GP 87). Crop Sci. 1981, 21, 148. [Google Scholar] [CrossRef]
- Maize and Wheat Improvement Center (CIMMYT). A Complete Listing of Improved Maize Germplasm from CIMMYT; Maize Program Special Report; Maize and Wheat Improvement Center (CIMMYT): Mexico City, Mexico, 1998; Available online: http://hdl.handle.net/10883/757 (accessed on 13 December 2022).
- Sauvion, N.; Calatayud, P.-A.; Thiéry, D.; Marion-Poll, F. Interactions Insectes-Plantes; Editions Quae: Versaille, France, 2013; ISBN 978-2-7592-2018-2. [Google Scholar]
- Chuang, W.P.; Herde, M.; Ray, S.; Castano-Duque, L.; Howe, G.A.; Luthe, D.S. Caterpillar attack triggers accumulation of the toxic maize protein RIP2. New Phytol. 2014, 201, 928–939. [Google Scholar] [CrossRef]
- Kim, E.Y.; Jung, J.K.; Kim, I.H.; Kim, Y. Chymotrypsin is a molecular target of insect resistance of three corn varieties against the Asian corn borer, Ostrinia furnacalis. PLoS ONE 2022, 17, e0266751. [Google Scholar] [CrossRef]
- Shivaji, R.; Camas, A.; Ankala, A.; Engelberth, J.; Tumlinson, J.H.; Williams, W.P.; Wilkinson, J.R.; Luthe, D.S. Plants on constant alert: Elevated levels of jasmonic acid and jasmonate-induced transcripts in caterpillar-resistant maize. J. Chem. Ecol. 2010, 36, 179–191. [Google Scholar] [CrossRef]
- Viana, P.A.; Potenza, M.R. Avaliação de antibiose e não-preferência em cultivares de milho selecionados com resistência à lagarta-do-cartucho. Bragantia 2000, 59, 27–33. [Google Scholar] [CrossRef]
- Waiss, A.C., Jr.; Chan, B.G.; Elliger, C.A.; Wiseman, B.R.; McMillian, W.W.; Widstrom, N.W.; Zuber, M.S.; Keaster, A.J. Maysin, a flavone glycoside from corn silks with antibiotic activity toward corn earworm. J. Econ. Entomol. 1979, 72, 256–258. [Google Scholar] [CrossRef]
- Wiseman, B.R.; Widstrom, N.W. Mechanisms of resistance in ‘Zapalote Chico’ corn silks to fall armyworm (Lepidoptera: Noctuidae) larvae. J. Econ. Entomol. 1986, 79, 1390–1393. [Google Scholar] [CrossRef]
- Wiseman, B.R.; McMillian, W.W.; Widstrom, N.W. Feeding of corn earworm in the laboratory on excised silks of selected corn entries with notes on Orius insidiosus. Fla. Entomol. 1976, 59, 305–308. [Google Scholar] [CrossRef]
- Wiseman, B.R.; Widstrom, N.W.; McMillian, W.W. Influence of corn silks on corn earworm feeding response. Fla. Entomol. 1981, 64, 395–399. [Google Scholar] [CrossRef]
- Wiseman, B.R.; Widstrom, N.W.; McMillian, W.W. Ear characteristics and mechanisms of resistance among selected corns to corn earworm. Fla. Entomol. 1977, 60, 97–103. [Google Scholar] [CrossRef]
- Williams, W.P.; Buckley, P.M.; Davis, F.M. Larval growth and behavior of the fall armyworm (Lepidoptera: Noctuidae) on callus initiated from susceptible and resistant corn hybrids. J. Econ. Entomol. 1985, 78, 951–954. [Google Scholar] [CrossRef]
- Yang, G.; Espelie, K.E.; Wiseman, B.R.; Isenhour, D.J. Effect of corn foliar cuticular lipids on the movement of fall armyworm neonate larvae. Fla. Entomol. 1993, 76, 302–316. [Google Scholar] [CrossRef]
- Smith, W.E.C.; Shivaji, R.; Williams, W.P.; Luthe, D.S.; Sandoya, G.V.; Smith, C.L.; Sparks, D.L.; Brown, A.E. A maize line resistant to herbivory constitutively releases (E)-β-Caryophyllene. J. Econ. Entomol. 2012, 105, 120–128. [Google Scholar] [CrossRef]
- Wiseman, B.R.; Widstrom, N.W.; McMillian, W.W. Influence of resistant and susceptible corn silks on selected developmental parameters of corn earworm (Lepidoptera: Noctuidae) larvae. J. Econ. Entomol. 1983, 76, 1288–1290. [Google Scholar] [CrossRef]
- Singh, G.M.; Xu, J.; Schaefer, D.; Day, R.; Wang, Z.; Zhang, F. Maize diversity for fall armyworm resistance in a warming world. Crop Sci. 2022, 62, 1–19. [Google Scholar] [CrossRef]
- Rocandio-Rodríguez, M.; Torres-Castillo, J.A.; Juárez-Aragón, M.C.; Chacón-Hernández, J.C.; Moreno-Ramírez, Y.d.R.; Mora-Ravelo, S.G.; Delgado-Martínez, R.; Hernández-Juárez, A.; Heinz-Castro, R.T.Q.; Reyes-Zepeda, F. Evaluation of Resistance of Eleven Maize Races (Zea mays L.) to the Red Spider Mite (Tetranychus merganser, Boudreaux). Plants 2022, 11, 1414. [Google Scholar] [CrossRef]
- Yang, G.; Wiseman, B.R.; Isenhour, D.J.; Espelie, K.E. Chemical and ultrastructural analysis of corn cuticular lipids and their effect on feeding by fall armyworm larvae. J. Chem. Ecol. 1993, 19, 2055–2074. [Google Scholar] [CrossRef]
- Yang, G.; Isenhour, D.J.; Espelie, K.E. Activity of maize leaf cuticular lipids in resistance to leaf-feeding by the fall armyworm. Fla. Entomol. 1991, 74, 229–236. [Google Scholar] [CrossRef]
- Haq, I.U.; Khurshid, A.; Inayat, R.; Kexin, Z.; Changzhong, L.; Ali, S.; Zuan, A.T.K.; Al-Hashimi, A.; Abbasi, A.M. Silicon-based induced resistance in maize against fall armyworm [Spodoptera frugiperda (Lepidoptera: Noctuidae)]. PLoS ONE 2021, 16, e0259749. [Google Scholar] [CrossRef]
- Nuambote-Yobila, O.; Musyoka, B.; Njuguna, E.; Bruce, A.; Khamis, F.; Subramanian, S.; Marion-Poll, F.; Calatayud, P.-A. Influence of Si in maize plants on Kenyan populations of Spodoptera frugiperda (Lepidoptera: Noctuidae). Phytoparasitica 2022, 50, 1025–1032. [Google Scholar] [CrossRef]
- Malook, S.U.; Liu, X.F.; Liu, W.; Qi, J.; Zhou, S. The race goes on: A fall armyworm resistant maize inbred line influences insect oral secretion elicitation activity and nullifies herbivore suppression of plant defense. BioRxiv 2021. [Google Scholar] [CrossRef]
- Ortega, A.; Vasal, S.K.; Mihm, J.A.; Hershey, C. Breeding for insect resistance in maize. In Breeding Plants Resistant to Insects; Maxwell, F.G., Jennings, P.R., Eds.; John Wiley and Sons: New York, NY, USA, 1980; pp. 371–419. [Google Scholar]
- Matova, P.M.; Kamutando, C.N.; Kutywayo, D.; Magorokosho, C.; Labuschagne, M. Fall armyworm tolerance of maize parental lines, experimental hybrids, and commercial cultivars in Southern Africa. Agronomy 2022, 12, 1463. [Google Scholar] [CrossRef]
- Goggin, F.L.; Lorence, A.; Topp, C.N. Applying high-throughput phenotyping to plant-insect interactions: Picturing more resistant crops. Curr. Opin. Insect Sci. 2015, 9, 69–76. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inbred Line | Larval Development Time n = 62–82 | Larval Survival n = 103–116 | Sex Ratio n = 11–15 | Weight of the Male Pupae n = 44–61 | Weight of the Female Pupae n = 38–61 |
|---|---|---|---|---|---|
| Mp716 | 17.7 ± 0.1 a | 82.7 ± 0.03 | 0.6 ± 0.08 | 197.2 ± 3.2 a | 188.3 ± 3.1 |
| CML71 | 17.7 ± 0.1 a | 73.3 ± 0.04 | 0.5 ± 0.08 | 191.6 ± 4.3 a | 186.4 ± 5.0 |
| CKSBL10008 | 17.9 ± 0.1 a | 80.3 ± 0.03 | 0.5 ± 0.04 | 197.1 ± 4.2 a | 181.4 ± 2.0 |
| CML125 | 17.0 ± 0.00 b | 73.4 ± 0.04 | 0.6 ± 0.09 | 191.8 ± 3.8 a | 187.0 ± 3.2 |
| CML370 | 17.1 ± 0.1 b | 82.7 ± 0.03 | 0.5 ± 0.01 | 201.9 ± 3.5 ab | 178.5 ± 2.9 |
| CKSBL10025 | 17.2 ± 0.1 b | 77.9 ± 0.04 | 0.6 ± 0.07 | 205.0 ± 2.8 ab | 188.8 ± 4.5 |
| CML444 | 17.0 ± 0.1 b | 81.3 ± 0.03 | 0.5 ± 0.06 | 214.9 ± 3.4 b | 193.3 ± 5.4 |
| p-Value of the ANOVA | ≤0.0001 | 0.2327 | 0.76 | ≤0.0001 | 0.1254 |
| Inbred Line | Pre-Oviposition Period n = 13–23 | Oviposition Period n = 15–23 | Number of Eggs Laid per Female n = 14–21 | % Eggs Hatched n = 15–22 |
|---|---|---|---|---|
| Mp716 | 2.4 ± 0.1 a | 5.7 ± 0.6 | 589.6 ± 41.2 | 95.8 ± 2.2 |
| CML71 | 2.5 ± 0.2 a | 5.2 ± 0.9 | 505.1 ± 68.4 | 98.3 ± 0.9 |
| CKSBL10008 | 2.3 ± 0.1 ab | 4.3 ± 0.6 | 484.9 ± 67.4 | 95.3 ± 2.3 |
| CML125 | 2.3 ± 0.1 ab | 5.0 ± 0.7 | 420.9 ± 84.7 | 98.9 ± 0.6 |
| CML370 | 2.0 ± 0.00 b | 3.1 ± 0.5 | 411.1 ± 62.9 | 92.1 ± 4.4 |
| CKSBL10025 | 2.0 ± 0.00 b | 5.0 ± 0.6 | 545.0 ± 47.4 | 95.7 ± 2.1 |
| CML444 | 2.0 ± 0.00 b | 4.9 ± 0.7 | 528.8 ± 59.5 | 92.6 ± 2.3 |
| p-Value of ANOVA | 0.0145 | 0.1539 | 0.3386 | 0.32 |
| Inbred Line | RGR under Lab Conditions | RGR under Net House Conditions | |||||
|---|---|---|---|---|---|---|---|
| On Leaves (10 Days) n = 116–127 | On Silks (10 Days) n = 40–74 | On Shelled Ear (7 Days) n = 10–17 | On Leaves (7 Days) n = 24–32 | On Leaves (14 Days) n = 23–31 | On Silks (14 Days) n = 10–31 | On Ears (14 Days) n = 46–136 | |
| Mp716 | 10.6 ± 0.3 bc | 2.8 ± 0.1 a | 3.4 ± 0.4 a | 1.2 ± 0.1 ab | 16.4 ± 1.0 abc | 1.5 ± 0.3 | 11.3 ± 0.8 ab |
| CML71 | 8.1 ± 0.4 a | 2.7 ± 0.1 a | 3.2 ± 0.4 a | 1.2 ± 0.1 ab | 15.0 ± 1.4 ab | 2.3 ± 0.5 | 10.3 ± 0.5 a |
| CKSBL10008 | 9.2 ± 0.4 ab | 4.6 ± 0.2 b | 3.8 ± 0.4 a | 1.0 ± 0.1 a | 13.8 ± 0.6 a | 3.5 ± 0.8 | 17.7 ± 0.8 c |
| CML125 | 12.0 ± 0.5 cd | 2.3 ± 0.1 a | 4.5 ± 0.5 a | 1.7 ± 0.1 b | 18.1 ± 1.2 bc | 3.2 ± 0.7 | 10.3 ± 0.6 a |
| CML370 | 11.3 ± 0.4 cd | 2.7 ± 0.1 a | 3.2 ± 0.6 a | 1.4 ± 0.1 ab | 19.1 ± 1.2 c | 2.5 ± 0.5 | 13.5 ± 0.9 b |
| CKSBL10025 | 11.2 ± 0.4 cd | 2.7 ± 0.1 a | 9.0 ± 1.2 b | 1.7 ± 0.1 b | 19.2 ± 1.2 c | 2.5 ± 0.5 | 20.1 ± 1.7 c |
| CML444 | 13.0 ± 0.5 d | 2.4 ± 0.1 a | 4.9 ± 0.8 a | 3.6 ± 0.3 c | 28.1 ± 1.0 d | 2.7 ± 0.44 | 11.4± 0.7 ab |
| p-Value of ANOVA | ≤0.0001 | ≤0.0001 | ≤0.0001 | ≤0.0001 | ≤0.0001 | 0.1188 | ≤0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nuambote-Yobila, O.; Bruce, A.Y.; Okuku, G.O.; Marangu, C.; Makumbi, D.; Beyene, Y.; Mahungu, N.-M.; Maruthi Prasanna, B.; Marion-Poll, F.; Calatayud, P.-A. Assessment of Resistance Mechanisms to Fall Armyworm, Spodoptera frugiperda in Tropical Maize Inbred Lines. Agronomy 2023, 13, 203. https://doi.org/10.3390/agronomy13010203
Nuambote-Yobila O, Bruce AY, Okuku GO, Marangu C, Makumbi D, Beyene Y, Mahungu N-M, Maruthi Prasanna B, Marion-Poll F, Calatayud P-A. Assessment of Resistance Mechanisms to Fall Armyworm, Spodoptera frugiperda in Tropical Maize Inbred Lines. Agronomy. 2023; 13(1):203. https://doi.org/10.3390/agronomy13010203
Chicago/Turabian StyleNuambote-Yobila, Ovide, Anani Y. Bruce, Gerphas Ogola Okuku, Charles Marangu, Dan Makumbi, Yoseph Beyene, Nzola-Meso Mahungu, Boddupalli Maruthi Prasanna, Frédéric Marion-Poll, and Paul-André Calatayud. 2023. "Assessment of Resistance Mechanisms to Fall Armyworm, Spodoptera frugiperda in Tropical Maize Inbred Lines" Agronomy 13, no. 1: 203. https://doi.org/10.3390/agronomy13010203
APA StyleNuambote-Yobila, O., Bruce, A. Y., Okuku, G. O., Marangu, C., Makumbi, D., Beyene, Y., Mahungu, N. -M., Maruthi Prasanna, B., Marion-Poll, F., & Calatayud, P. -A. (2023). Assessment of Resistance Mechanisms to Fall Armyworm, Spodoptera frugiperda in Tropical Maize Inbred Lines. Agronomy, 13(1), 203. https://doi.org/10.3390/agronomy13010203