
Introgression of Seedling Plant Resistance to Leaf Rust from Agropyron cristatum into Wheat by Induced Homoeologous Recombination

Abstract
:1. Introduction
2. Materials and Methods
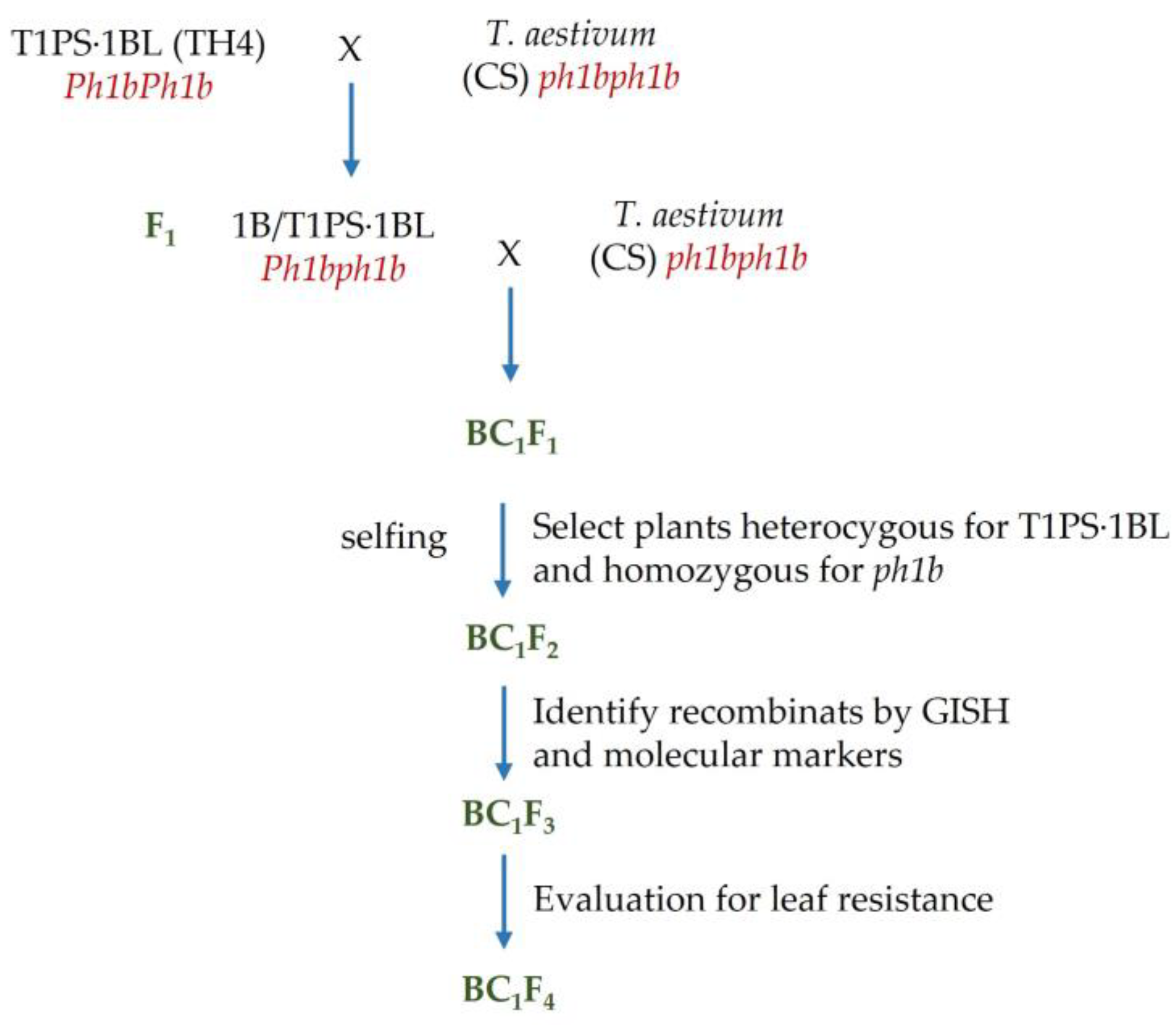
2.1. Plant Material
2.2. DNA Markers Analysis
2.3. Fluorescence In Situ Hybridization
2.4. Evaluation of Leaf Rust Resistance
2.5. Fungal Material
2.6. Inoculation Assays
2.7. Characterization of Macroscopic Components of Resistance
2.8. Characterization of Microscopic Components of Resistance
2.9. Statistical Analysis
3. Results
3.1. Screening for Chromosome Arm 1PS-Specific Molecular Markers
3.2. Identification of Individuals with a Single 1PS·1BL Translocation and Homozygous for the ph1b Mutation
3.3. Wheat-A. cristatum Translocations Involving 1PS
3.4. Macroscopic Components of Resistance to Leaf Rust Infection
3.5. Microscopic Components of Resistance to Leaf Rust Infection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- The Wheat Initiative. Available online: https://www.wheatinitiative.org/ (accessed on 1 June 2022).
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The global burden of pathogens and pests on major food crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Ellis, J.G.; Lagudah, E.S.; Spielmeyer, W.; Dodds, P.N. The past, present and future of breeding rust resistant wheat. Front. Plant Sci. 2014, 5, 641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Méndez, N.; Miguel-Rojas, C.; Jimenez-Berni, J.A.; Gomez-Candon, D.; Pérez-de-Luque, A.; Fereres, E.; Catala-Forner, M.; Villegas, D.; Sillero, J.C. Plant breeding and management strategies to minimize the impact of water scarcity and biotic stress in cereal crops under Mediterranean conditions. Agronomy 2022, 12, 75. [Google Scholar] [CrossRef]
- Huerta-Espino, J.; Singh, R.P.; Germán, S.; McCallum, B.D.; Park, R.F.; Chen, W.Q.; Bhardwaj, S.C.; Goyeau, H. Global status of wheat leaf rust caused by Puccinia triticina. Euphytica 2011, 179, 143–160. [Google Scholar] [CrossRef]
- McIntosh, R.; Dubcovsky, J.; Rogers, W.; Morris, C.; Appels, R.; Xia, X. Catalogue of gene symbols for wheat 2020 Supplement. 2017. Available online: https://wheat.pw.usda.gov/GG3/Wheat_Gene_Catalog_Documents (accessed on 11 June 2022).
- Omara, R.I.; Nehela, Y.; Mabrouk, O.I.; Elsharkawy, M.M. The emergence of new aggressive leaf rust races with the potential to supplant the resistance of wheat cultivars. Biology 2021, 10, 925. [Google Scholar] [CrossRef] [PubMed]
- Dewey, D.R. The genomic system of classification as a guide to intergeneric hybridization with the perennial Triticeae. In Gene Manipulation in Plant Improvement; Gustafson, J.P., Ed.; Springer: Boston, MA, USA, 1984; pp. 209–279. [Google Scholar]
- Asay, K.H.; Dewey, D.R.; Gomm, F.B.; Horton, W.H.; Jensen, K.B. Genetic progress through hybridization of induced and natural tetraploids in crested wheatgrass. J. Range Manag. 1986, 39, 261–263. [Google Scholar] [CrossRef]
- Ochoa, V.; Madrid, E.; Said, M.; Rubiales, D.; Cabrera, A. Molecular and cytogenetic characterization of a common wheat-Agropyron cristatum chromosome translocation conferring resistance to leaf rust. Euphytica 2015, 201, 89–95. [Google Scholar] [CrossRef]
- Jiang, B.; Liu, T.; Li, H.; Han, H.; Li, L.; Zhang, J.; Yang, X.; Zhou, S.; Li, X.; Liu, W. Physical mapping of a novel locus conferring leaf rust resistance on the long arm of Agropyron cristatum chromosome 2P. Front Plant Sci. 2018, 9, 817. [Google Scholar] [CrossRef] [Green Version]
- Ji, X.; Liu, T.; Xu, S.; Wang, Z.; Han, H.; Zhou, S.; Guo, B.; Zhang, J.; Yang, X.; Li, X.; et al. Comparative transcriptome analysis reveals the gene expression and regulatory characteristics of broad-spectrum immunity to leaf rust in a wheat-Agropyron cristatum 2P addition line. Int. J. Mol. Sci. 2022, 23, 7330. [Google Scholar] [CrossRef]
- Zhang, Z.; Song, L.; Han, H.; Zhou, S.; Zhang, J.; Yang, X.; Li, X.; Liu, W.; Li, L. Physical localization of a locus from Agropyron cristatum conferring resistance to stripe rust in common wheat. Int. J. Mol. Sci. 2017, 18, 2403. [Google Scholar] [CrossRef]
- Copete, A.; Cabrera, A. Chromosomal location of genes for resistance to powdery mildew in Agropyron cristatum and mapping of conserved orthologous set molecular markers. Euphytica 2017, 213, 189. [Google Scholar] [CrossRef]
- Li, H.; Jiang, B.; Wang, J.; Lu, Y.; Zhang, J.; Pan, C.; Yang, X.; Li, X.; Liu, W.; Li, L. Mapping of novel powdery mildew resistance gene(s) from Agropyron cristatum chromosome 2P. Theor. Appl. Genet. 2017, 130, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Bayat, H.; Nemati, H.; Tehranifar, A.; Gazanchian, A. Screening different crested wheatgrass (Agropyron cristatum (L.) Gaertner.) accessions for drought stress tolerance. Arch. Agron. Soil Sci. 2016, 62, 769–780. [Google Scholar] [CrossRef]
- Dewey, D.R. Breeding crested wheatgrass for salt tolerance. Crop Sci. 1962, 2, 403–407. [Google Scholar] [CrossRef]
- Limin, A.E.; Fowler, D.B. Cold hardiness of forage grasses grown on the Canadian prairies. Can, J. Plant Sci. 1987, 67, 1111–1115. [Google Scholar] [CrossRef]
- Cabrera, A.; Castellano, L.; Recio, R.; Alvarez, J.B. Chromosomal location and molecular characterization of three grain hardness genes in Agropyron cristatum. Euphytica 2019, 215, 165. [Google Scholar] [CrossRef]
- Cabrera, A.; Copete-Parada, A.; Madrid, E. Cloning and characterization of a putative orthologue of the wheat vernalization (VRN1) gene in perennial wheatgrass (Agropyron cristatum). Plant Breed. 2020, 139, 1290–1298. [Google Scholar] [CrossRef]
- Riley, R.; Chapman, V. Genetic control of the cytologically diploid behaviour of hexaploid wheat. Nature 1958, 182, 713–715. [Google Scholar] [CrossRef]
- Sears, E.R. An induced mutant with homoeologous pairing in common wheat. Can. J. Genet. Cytol. 1977, 19, 585–593. [Google Scholar] [CrossRef]
- Sears, E.R. A wheat mutation conditioning an intermediate level of homoeologous chromosome pairing. Can. J. Genet. Cytol. 1982, 24, 715–719. [Google Scholar] [CrossRef]
- Qi, L.; Friebe, B.; Zhang, P.; Gill, B.S. Homoeologous recombination, chromosome engineering and crop improvement. Chromosome Res. 2007, 15, 3–19. [Google Scholar] [CrossRef] [PubMed]
- Hao, M.; Zhang, L.; Ning, S.; Huang, L.; Yuan, Z.; Wu, B.; Yan, Z.; Dai, S.; Jiang, B.; Zheng, Y.; et al. The resurgence of introgression breeding, as exemplified in wheat improvement. Front Plant Sci. 2020, 6, 252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Payne, P.I. Genetics of wheat storage proteins and the effects of allelic variation on bread-making quality. Annu. Rev. Plant Physiol. 1987, 38, 141–153. [Google Scholar] [CrossRef]
- Graybosch, R.A.; Peterson, C.J.; Hansen, L.E.; Worral, D.; Shelton, D.R. and Lukaszewski, A.J. Comparative flour quality and protein characteristics of 1BL/1RS and 1AL/1RSwheat-rye translocations. J. Cereal Sci. 1993, 17, 95–106. [Google Scholar] [CrossRef]
- Jubault, M.; Tanguy, A.M.; Abélard, P.; Coriton, O.; Dusautoir, J.C.; Jahier, J. Attempts to induce homoeologous pairing between wheat and Agropyron cristatum genomes. Genome 2006, 49, 190–193. [Google Scholar] [CrossRef]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acid Res. 1980, 8, 4321–4325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Liu, W.; Lu, Y.; Liu, Q.; Yang, X.; Li, X. A resource of large-scale molecular markers for monitoring Agropyron cristatum chromatin introgression in wheat background based on transcriptome sequences. Sci. Rep. 2017, 7, 11942. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Sun, H.; Guan, C.; Cui, J.; Zhang, Q.; Liu, M.; Zhang, M.; Guo, Q.; Hou, Y.; Xiang, M.; et al. Physical information of 2705 PCR-based molecular markers and the evaluation of their potential use in wheat. J. Genet. 2019, 98, 69. [Google Scholar] [CrossRef]
- Raats, D.; Frenkel, Z.; Krugman, T.; Dodek, I.; Sela, H.; Šimková, H.; Mage, F.; Cattonaro, F.; Vautrin, S.; Fahima, T.; et al. The physical map of wheat chromosome 1BS provides insights into its gene space organization and evolution. Genome Biol. 2013, 14, R138. [Google Scholar] [CrossRef]
- Wang, L.H.; Zhao, X.L.; He, Z.H.; Ma, W.; Appels, R.; Peña, R.J.; Xia, X.C. Characterization of low-molecular-weight glutenin subunit Glu-B3 genes and development of STS markers in common wheat (Triticum aestivum L.). Theor. Appl. Genet. 2009, 118, 525–539. [Google Scholar] [CrossRef]
- Devos, K.M.; Bryan, G.J.; Collins, A.J.; Stephenson, P.; Gale, M.D. Application of two microsatellite sequences in wheat storage proteins as molecular markers. Theor. Appl. Genet. 1995, 90, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Howell, T.; Hale, I.; Jankuloski, L.; Bonafede, M.; Gilbert, M.; Dubcovsky, J. Mapping a region within the 1RS.1BL translocation in common wheat affecting grain yield and canopy water status. Theor. Appl. Genet. 2014, 127, 2695–2709. [Google Scholar] [CrossRef] [Green Version]
- Sourdille, P.; Singh, P.; Cadalen, T.; Brown-Guedira, G.L.; Gay, G.; Qi, L.; Gill, B.S.; Dufour, P.; Murigneux, A.; Bernard, M. Microsatellite-based deletion bin system for the establishment of genetic-physical map relationships in wheat (Triticum aestivum L.). Func. Integr. Genomics 2004, 4, 12–25. [Google Scholar] [CrossRef] [PubMed]
- Somers, D.J.; Isaac, P.; Edwards, K. A high-density microsatellite consensus map for bread wheat (Triticum aestivum L.). Theor. Appl. Genet. 2004, 109, 1105–1114. [Google Scholar] [CrossRef]
- Röder, M.S.; Korzun, V.; Wendehake, K.; Plaschle, J.; Tixier, M.H.; Leroy, P.; Ganal, M.W. A microsatellite map of wheat. Genetics 1998, 149, 2007–2023. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lai, J.; Liu, G.; Chen, F. Development of a Scar marker for the Ph1 locus in common wheat and its application. Crop Sci. 2002, 42, 1365–1368. [Google Scholar] [CrossRef]
- Cabrera, A.; Martin, A.; Barro, F. In-situ comparative mapping (ISCM) of Glu-1 loci in Triticum and Hordeum. Chromosome Res. 2002, 10, 49–54. [Google Scholar] [CrossRef]
- Gerlach, W.L.; Bedbrook, J.R. Cloning and characterization of ribosomal RNA genes from wheat and barley. Nucleic Acids Res. 1979, 7, 1869–1885. [Google Scholar] [CrossRef]
- Porras, R.; Miguel-Rojas, C.; Pérez-de-Luque, A.; Sillero, J.C. Macro- and microscopic characterization of components of resistance against Puccinia striiformis f. sp. tritici in a collection of Spanish bread wheat cultivars. Agronomy 2022, 12, 1239. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schimid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- McNeal, F.H.; Konzak, C.F.; Smith, E.P.; Tate, W.S.; Russell, T.S. A uniform system for recording and processing cereal research data. US Agric. Res. Serv. 1971, 42, 34–121. [Google Scholar]
- Soleiman, N.H.; Solis, I.; Sillero, J.C.; Herrera-Foesel, S.A.; Ammar, K.; Martínez, F. Evaluation of macroscopic and microscopic components of partial resistance to leaf rust in durum wheat. J. Phytopathol. 2014, 162, 359–366. [Google Scholar] [CrossRef]
- Rubiales, D.; Niks, R.E. Characterization of Lr34, a major gene conferring non hypersensitive resistance to wheat leaf rust. Plant Dis. 1995, 79, 1208–1212. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Found. Stat. Comput.: Vienna, Austria, 2022; Available online: https://www.r-project.org/ (accessed on 1 April 2022).
- Chen, Q.; Jahier, J.; Cauderon, Y. Production and cytogenetic analysis of BC1, BC2 and BC3 progenies of an intergeneric hybrid between Triticum aestivum (L.) Thell. and tetraploid Agropyron cristatum (L.) Gaerth. Theor. Appl. Genet. 1992, 94, 698–703. [Google Scholar] [CrossRef]
- Martín, A.; Cabrera, A.; Esteban, E.; Hernández, P.; Ramírez, M.C.; Rubiales, D. A fertile amphiploid between diploid wheat (Triticum tauschii) and crested wheatgrass (Agropyron cristatum). Genome 1999, 42, 519–524. [Google Scholar] [CrossRef]
- Soliman, M.H.; Rubiales, D.; Cabrera, A. A fertile amphiploid between durum wheat (Triticum turgidum) and the Agroticum amphiploid (Agropyron cristatum x T. tauschii). Hereditas 2001, 135, 183–186. [Google Scholar] [CrossRef] [Green Version]
- Soliman, M.H.; Cabrera, A.; Sillero, J.C.; Rubiales, D. Genomic constitution and expression of disease resistance in Agropyron cristatum x durum wheat derivatives. Breed. Sci. 2007, 57, 17–21. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Ma, H.; Zhang, J.; Zhou, S. Molecular cytogenetic characterization of an Agropyron cristatum 6PL chromosome segment conferring superior kernel traits in wheat. Euphytica 2018, 214, 198. [Google Scholar] [CrossRef]
- Copete-Parada, A.; Palomino, C.; Cabrera, A. Development and characterization of wheat-Agropyron cristatum introgression lines induced by gametocidal genes and Ph1 mutant. Agronomy 2021, 11, 277. [Google Scholar] [CrossRef]
- Lukaszewski, A.J. Physical distribution of translocation breakpoints in homoeologous recombinants induced by the absence of the Ph1 gene in wheat and triticale. Theor. Appl. Genet. 1995, 90, 714–719. [Google Scholar] [CrossRef]
- Said, M.; Copete Parada, A.; Gaál, E.; Molnár, I.; Cabrera, A.; Doležel, J.; Vrána, J. Uncovering homeologous relationships between tetraploid Agropyron cristatum and bread wheat genomes using COS markers. Theor. Appl. Genet. 2019, 132, 2881–2898. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Yan, B.; Li, F.; Zhang, J.; Zhang, J.; Ma, H.; Liu, W.; Lu, Y.; Yang, X.; Liu, X.; et al. RNA-Seq analysis provides first insights into the phylogenetic relationships and interespecific variation between Agropyron cristatum and wheat. Front. Plant Sci. 2017, 8, 1644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Said, M.; Hřibová, E.; Danilova, T.V.; Karafiátová, M.; Čížková, J.; Friebe, B.; Doležel, J.; Gill, B.S.; Vrána, J. The Agropyron cristatum karyotype, chromosome structure and cross-genome homeology as revealed by fluorescence in situ hybridization with tandem repeats and wheat single-gene probes. Theor. Appl. Genet. 2018, 131, 2213–2227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mago, R.; Miah, H.; Lawrence, G.J.; Wellings, C.R.; Spielmeyer, W.; Bariana, H.S.; McIntosh, R.A.; Pryor, A.J.; Ellis, J.G. High-resolution mapping and mutation analysis separate the rust resistance genes Sr31, Lr26 and Yr9 on the short arm of rye chromosome 1. Theor. Appl. Genet. 2005, 112, 41–50. [Google Scholar] [CrossRef]
- Lukaszewski, A.J. Manipulation of the 1RS.1BL translocation in wheat by induced homologous recombination. Crop Sci. 2000, 40, 216–225. [Google Scholar] [CrossRef]
- Liu, W.X.; Rouse, M.; Friebe, B.; Jin, Y.; Gill, B.; Pumphrey, M.O. Discovery and molecular mapping of a new gene conferring resistance to stem rust, Sr53, derived from Aegilops geniculata and characterization of spontaneous translocation stocks with reduced alien chromatin. Chromosome Res. 2011, 19, 669–682. [Google Scholar] [CrossRef]
- Niu, Z.; Klindworth, D.L.; Yu, G.; Friesen, T.L.; Chao, S.; Jin, Y.; Cai, X.; Xu, S.S. Development and characterization of wheat lines carrying stem rust resistance gene Sr43 derived from Thinopyrum ponticum. Theor. Appl. Genet. 2014, 127, 969–980. [Google Scholar] [CrossRef]
- Li, H.; Dong, Z.; Ma, C.; Tian, X.; Qi, Z.; Wu, N.; Friebe, B.; Xiang, Z.; Xia, Q.; Liu, W.; et al. Physical mapping of stem rust resistance gene Sr52 from Dasypyrum villosum based on ph1b-induced homoeologous recombination. Int. J. Mol. Sci. 2019, 20, 4887. [Google Scholar] [CrossRef] [Green Version]
- Lukaszewski, A.J.; Cowger, C. Re-engineering of the Pm21 transfer from Haynaldia villosa to bread wheat by induced homoeologous recombination. Crop Sci. 2017, 57, 2590–2594. [Google Scholar] [CrossRef]
- Wan, W.; Xiao, J.; Li, M.; Tang, X.; Wen, M.; Cheruiyot, A.K.; Li, Y.; Wang, H.; Wang, X. Fine mapping of wheat powdery mildew resistance gene Pm6 using 2B/2G homoeologous recombinants induced by the ph1b mutant. Theor. Appl. Genet. 2020, 133, 1265–1275. [Google Scholar] [CrossRef]
- Zhao, R.; Wang, H.; Xiao, J.; Bie, T.; Cheng, S.; Jia, Q.; Yuan, C.; Zhang, R.; Cao, A.; Chen, P.; et al. Induction of 4VS chromosome recombination using the CS ph1b mutant and mapping of the yellow mosaic virus resistance gene from Haynaldia villosa. Theor. Appl. Genet. 2013, 126, 2921–2930. [Google Scholar] [CrossRef] [PubMed]
- Danilova, T.V.; Zhang, G.; Liu, W.; Friebe, B.; Gill, B.S. Homoeologous recombination-based transfer and molecular cytogenetic mapping of a wheat streak mosaic virus and Triticum mosaic virus resistance gene Wsm3 from Thinopyrum intermedium to wheat. Theor. Appl. Genet. 2017, 130, 549–556. [Google Scholar] [CrossRef] [PubMed]
- Fedak, G.; Chi, D.; Wolfe, D.; Oullet, T.; Cao, W.; Han, F.; Xue, A. Transfer of fusarium head blight resistance from Thinopyrum elongatum to bread wheat cultivar Chinese Spring. Genome 2021, 64, 959–968. [Google Scholar] [CrossRef] [PubMed]
- Kolmer, J.A. Genetics of resistance to wheat leaf rust. Annu. Rev. Phytopathol. 1996, 34, 435–455. [Google Scholar] [CrossRef] [PubMed]
- Martínez, F.; Niks, R.E.; Singh, R.P.; Rubiales, D. Characterization of Lr46, a gene conferring partial resistance to wheat leaf rust. Hereditas 2001, 135, 111–114. [Google Scholar] [CrossRef] [Green Version]
- Kloppers, F.J.; Pretorius, Z.A. Effects of combinations amongst genes Lr13, Lr34 and Lr37 on components of resistance in wheat to leaf rust. Plant Pathol. 1997, 46, 737–750. [Google Scholar] [CrossRef]
- Martínez, F.; Sillero, J.C.; Rubiales, D. Effect of host plant resistance on haustorium formation in cereal rust fungi. J. Phytopathol. 2004, 152, 381–382. [Google Scholar] [CrossRef]
- Wang, X.; McCallum, B.D.; Fetch, T.; Bakkeren, G.; Marais, G.F.; Saville, B.J. Comparative microscopic and molecular analysis of Thatcher near-isogenic lines with wheat leaf rust resistance genes Lr2a, Lr3, LrB or Lr9 upon challenge with different Puccinia triticina races. Plant Pathol. 2013, 62, 698–707. [Google Scholar] [CrossRef]
- Saleem, K.; Sørensen, C.K.; Labouriau, R.; Hovmøller, M.S. Spatiotemporal changes in fungal growth and host responses of six yellow rust resistant near-isogenic lines of wheat. Plant Pathol. 2019, 68, 1320–1330. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | RLP (Hours) | IT 3 | RIF (Pustules/cm2) | RPS (mm2) |
|---|---|---|---|---|
| Califa Sur | 100.00 ± 0.00 [163 1] c 2 | 9 | 100.00 (10.00 ± 0.00) [68] a | 100.00 ± 0.00 [0.132] a |
| CS | 104.42 ± 2.55 b | 9 | 97.70 (9.87 ± 0.67) a | 88.33 ± 5.17 b |
| TH4 | 118.41 ± 0.65 a | 6 (9) | 18.58 (4.28 ± 0.58) c | 33.48 ± 4.81 e |
| BC1F4-15-10-2 4 | 106.43 ± 1.45 b | 2 (6) | 10.47 (3.23 ± 0.23) d | 35.72 ± 5.70 e |
| BC1F4-37-2-21 4 | 104.52 ± 0.81 b | 2 (6) | 11.79 (3.43 ± 0.23) d | 38.47 ± 7.04 de |
| BC1F4-43-2-4 5 | 105.27 ± 2.26 b | 5 (6) | 19.39 (4.37 ± 0.63) c | 47.34 ± 9.32 d |
| BC1F4-45-2-7 6 | 94.92 ± 0.13 d | 9 | 73.87 (8.59 ± 0.24) b | 76.17 ± 2.83 c |
| Genotype | RCS (mm2) | EA | EST |
|---|---|---|---|
| Califa Sur | 100.00 ± 0.00 [0.455 1] a 2 | 9.47 (3.06 ± 0.42) c | 90.53 (3.06 ± 0.42) a |
| CS | 100.97 ± 5.95 a | 11.75 (3.42 ± 0.25) c | 88.25 (3.42 ± 0.25) a |
| TH4 | 29.81 ± 3.95 e | 52.53 (7.24 ± 0.29) a | 47.47 (7.24 ± 0.29) c |
| BC1F4-15-10-2 3 | 56.27 ± 1.91 d | 29.20 (5.32 ± 1.13) b | 70.80 (5.32 ± 1.13) b |
| BC1F4-37-2-21 3 | 57.32 ± 3.22 d | 31.31 (5.54 ± 0.99) b | 68.69 (5.54 ± 0.99) b |
| BC1F4-43-2-4 4 | 67.59 ± 1.22 c | 16.82 (4.09 ± 0.43) c | 83.18 (4.09 ± 0.43) a |
| BC1F4-45-2-7 5 | 92.61 ± 5.62 b | 14.10 (3.75 ± 0.20) c | 85.90 (3.75 ± 0.20) a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cabrera, A.; Porras, R.; Palomino, C.; Sillero, J.C. Introgression of Seedling Plant Resistance to Leaf Rust from Agropyron cristatum into Wheat by Induced Homoeologous Recombination. Agronomy 2023, 13, 334. https://doi.org/10.3390/agronomy13020334
Cabrera A, Porras R, Palomino C, Sillero JC. Introgression of Seedling Plant Resistance to Leaf Rust from Agropyron cristatum into Wheat by Induced Homoeologous Recombination. Agronomy. 2023; 13(2):334. https://doi.org/10.3390/agronomy13020334
Chicago/Turabian StyleCabrera, Adoración, Rafael Porras, Carmen Palomino, and Josefina Carmen Sillero. 2023. "Introgression of Seedling Plant Resistance to Leaf Rust from Agropyron cristatum into Wheat by Induced Homoeologous Recombination" Agronomy 13, no. 2: 334. https://doi.org/10.3390/agronomy13020334
APA StyleCabrera, A., Porras, R., Palomino, C., & Sillero, J. C. (2023). Introgression of Seedling Plant Resistance to Leaf Rust from Agropyron cristatum into Wheat by Induced Homoeologous Recombination. Agronomy, 13(2), 334. https://doi.org/10.3390/agronomy13020334