Calcium- and Magnesium-Enriched Organic Fertilizer and Plant Growth-Promoting Rhizobacteria Affect Soil Nutrient Availability, Plant Nutrient Uptake, and Secondary Metabolite Production in Aloe vera (Aloe barbadensis Miller) Grown under Field Conditions

 , ,
, , 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Applications
2.2. Soil Properties Analysis
2.3. Morpho-Anatomical Measurments
2.4. Leaf and Gel Analysis for Minerals
2.5. Acemannan Quantification
2.6. Quantification of Total Phenolic Content
2.7. Data Analysis
3. Results and Discussion
3.1. Soil Properties
3.2. Plant Growth
3.3. Leaf Rind Mineral Concentrations
3.4. Gel Macronutrient Concentration
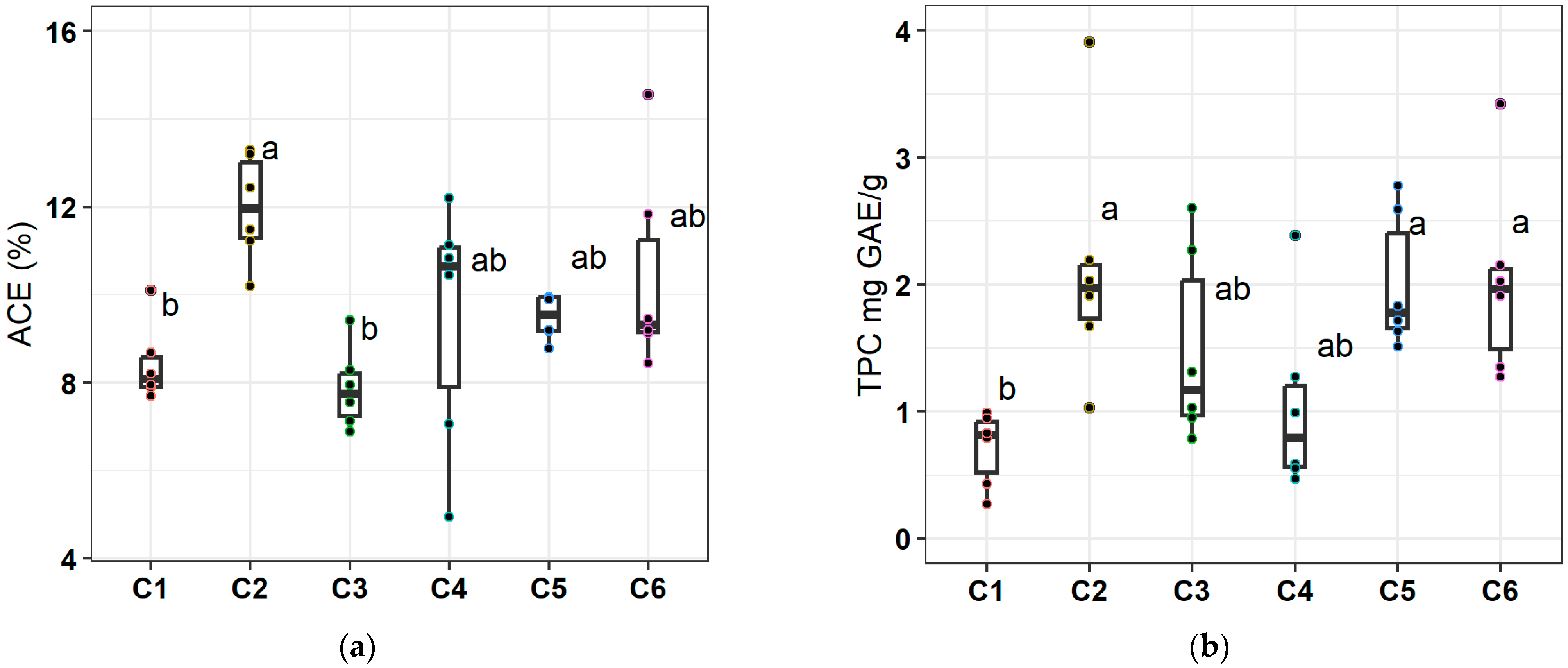
3.5. Acemannan and TPC Concentrations
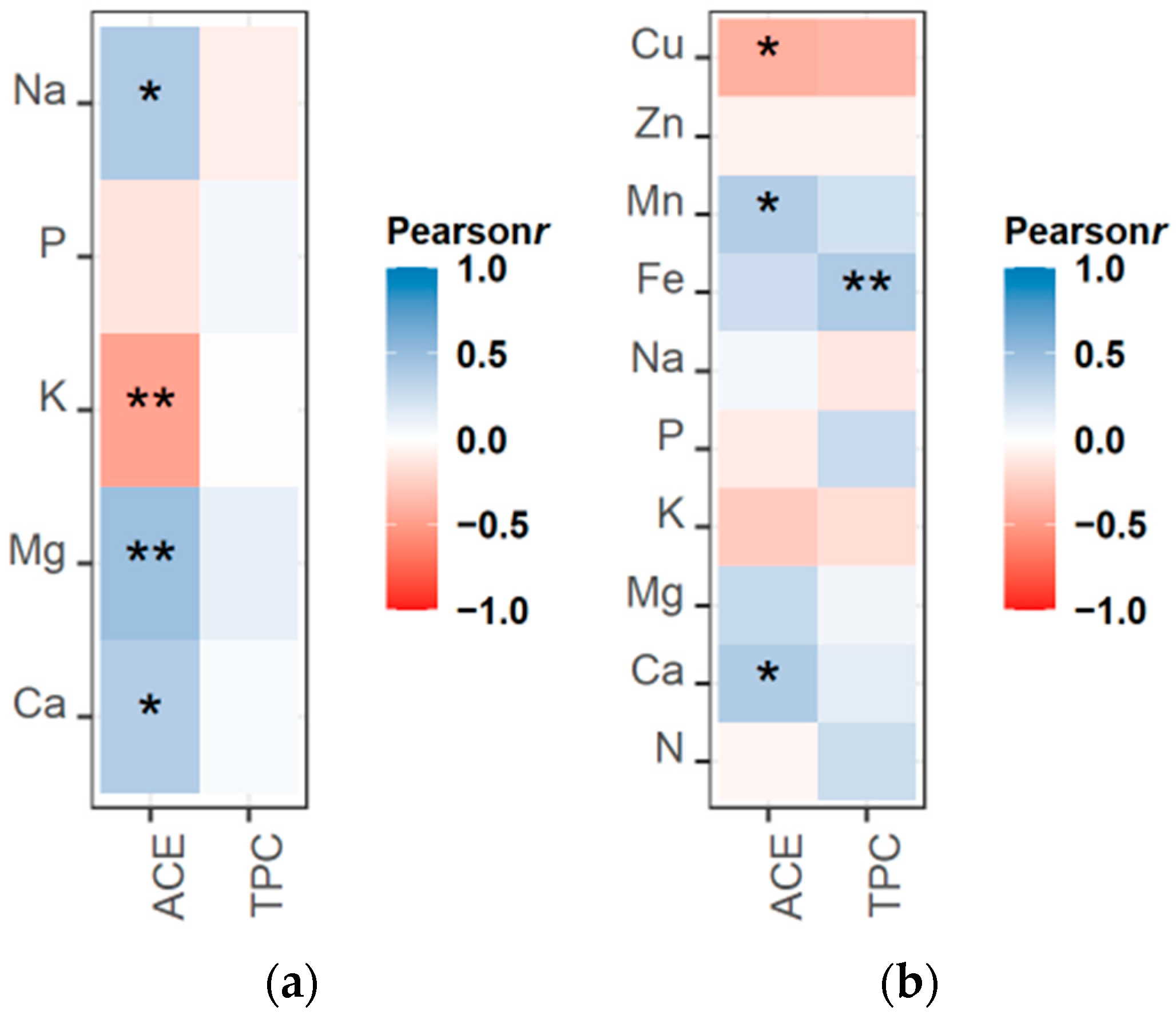
3.6. Correlation Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Añibarro-Ortega, M.; Pinela, J.; Barros, L.; Ćirić, A.; Silva, S.P.; Coelho, E.; Mocan, A.; Calhelha, R.C.; Soković, M.; Coimbra, M.A.; et al. Compositional Features and Bioactive Properties of Aloe vera Leaf (Fillet, Mucilage, and Rind) and Flower. Antioxidants 2019, 8, 444. [Google Scholar] [CrossRef]
- Kahramanoğlu, İ.; Chen, C.; Chen, J.; Wan, C. Chemical Constituents, Antimicrobial Activity, and Food Preservative Characteristics of Aloe vera Gel. Agronomy 2019, 9, 831. [Google Scholar] [CrossRef]
- Sánchez-Machado, D.I.; López-Cervantes, J.; Sendón, R.; Sanches-Silva, A. Aloe vera: Ancient Knowledge with New Frontiers. Trends Food Sci. Technol. 2017, 61, 94–102. [Google Scholar] [CrossRef]
- Eshun, K.; He, Q. Aloe vera: A Valuable Ingredient for the Food, Pharmaceutical and Cosmetic Industries—A Review. Crit. Rev. Food Sci. Nutr. 2004, 44, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Salinas, P.; Salinas, C.; Contreras, R.A.; Zuñiga, G.E.; Dupree, P.; Cardemil, L. Water Deficit and Abscisic Acid Treatments Increase the Expression of a Glucomannan Mannosyltransferase Gene (GMMT) in Aloe vera Burm. F. Phytochemistry 2019, 159, 90–101. [Google Scholar] [CrossRef] [PubMed]
- Kaparakou, E.H.; Kanakis, C.D.; Gerogianni, M.; Maniati, M.; Vekrellis, K.; Skotti, E.; Tarantilis, P.A. Quantitative Determination of Aloin, Antioxidant Activity, and Toxicity of Aloe vera Leaf Gel Products from Greece. J. Sci. Food Agric. 2021, 101, 414–423. [Google Scholar] [CrossRef] [PubMed]
- Cristiano, G.; Murillo-Amador, B.; De Lucia, B. Propagation Techniques and Agronomic Requirements for the Cultivation of Barbados Aloe (Aloe vera (L.) Burm. F.)—A Review. Front. Plant Sci. 2016, 7, 1410. [Google Scholar] [CrossRef]
- Ning, C.; Gao, P.; Wang, B.; Lin, W.; Jiang, N.; Cai, K. Impacts of Chemical Fertilizer Reduction and Organic Amendments Supplementation on Soil Nutrient, Enzyme Activity and Heavy Metal Content. J. Integr. Agric. 2017, 16, 1819–1831. [Google Scholar] [CrossRef]
- Vega-Gálvez, A.; Miranda, M.; Aranda, M.; Henriquez, K.; Vergara, J.; Tabilo-Munizaga, G.; Pérez-Won, M. Effect of High Hydrostatic Pressure on Functional Properties and Quality Characteristics of Aloe vera Gel (Aloe barbadensis Miller). Food Chem. 2011, 129, 1060–1065. [Google Scholar] [CrossRef]
- Kumar, S.; Yadav, A.; Yadav, M.; Yadav, J.P. Effect of Climate Change on Phytochemical Diversity, Total Phenolic Content and in Vitro Antioxidant Activity of Aloe vera (L.) Burm.f. BMC Res. Notes 2017, 10, 60. [Google Scholar] [CrossRef] [Green Version]
- Canche-Escamilla, G.; Colli-Acevedo, P.; Borges-Argaez, R.; Quintana-Owen, P.; May-Crespo, J.F.; Cáceres-Farfan, M.; Yam Puc, J.A.; Sansores-Peraza, P.; Vera-Ku, B.M. Extraction of Phenolic Components from an Aloe vera (Aloe barbadensis Miller) Crop and Their Potential as Antimicrobials and Textile Dyes. Sustain. Chem. Pharm. 2019, 14, 100168. [Google Scholar] [CrossRef]
- Quezada, M.P.; Salinas, C.; Gotteland, M.; Cardemil, L. Acemannan and Fructans from Aloe vera (Aloe barbadensis Miller) Plants as Novel Prebiotics. J. Agric. Food Chem. 2017, 65, 10029–10039. [Google Scholar] [CrossRef]
- Minjares-Fuentes, J.R.; Femenia, A. Effect of Processing on the Bioactive Polysaccharides and Phenolic Compounds from Aloe vera (Aloe barbadensis Miller). In Dietary Fiber Functionality in Food and Nutraceuticals; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2017; pp. 263–287. ISBN 978-1-119-13810-5. [Google Scholar]
- Hamman, J.H. Composition and Applications of Aloe vera Leaf Gel. Molecules 2008, 13, 1599–1616. [Google Scholar] [CrossRef] [PubMed]
- Sadgrove, N.J.; Simmonds, M.S.J. Pharmacodynamics of Aloe vera and Acemannan in Therapeutic Applications for Skin, Digestion, and Immunomodulation. Phytother. Res. 2021, 35, 6572–6584. [Google Scholar] [CrossRef] [PubMed]
- Voiniciuc, C. Modern Mannan: A Hemicellulose’s Journey. New Phytol. 2022, 234, 1175–1184. [Google Scholar] [CrossRef]
- He, C.; Wu, K.; Zhang, J.; Liu, X.; Zeng, S.; Yu, Z.; Zhang, X.; Teixeira da Silva, J.A.; Deng, R.; Tan, J.; et al. Cytochemical Localization of Polysaccharides in Dendrobium Officinale and the Involvement of DoCSLA6 in the Synthesis of Mannan Polysaccharides. Front. Plant Sci. 2017, 8, 173. [Google Scholar] [CrossRef]
- Femenia, A.; Sánchez, E.S.; Simal, S.; Rosselló, C. Compositional Features of Polysaccharides from Aloe vera (Aloe barbadensis Miller) Plant Tissues. Carbohydr. Polym. 1999, 39, 109–117. [Google Scholar] [CrossRef]
- Yang, L.; Wen, K.-S.; Ruan, X.; Zhao, Y.-X.; Wei, F.; Wang, Q. Response of Plant Secondary Metabolites to Environmental Factors. Molecules 2018, 23, 762. [Google Scholar] [CrossRef]
- Douglas, A.E. Strategies for Enhanced Crop Resistance to Insect Pests. Annu. Rev. Plant Biol. 2018, 69, 637–660. [Google Scholar] [CrossRef]
- Murthy, H.N.; Lee, E.-J.; Paek, K.-Y. Production of Secondary Metabolites from Cell and Organ Cultures: Strategies and Approaches for Biomass Improvement and Metabolite Accumulation. Plant Cell Tiss. Organ Cult. 2014, 118, 1–16. [Google Scholar] [CrossRef]
- Arsenault, P.R.; Wobbe, K.K.; Weathers, P.J. Recent Advances in Artemisinin Production through Heterologous Expression. Curr. Med. Chem. 2008, 15, 2886–2896. [Google Scholar] [CrossRef]
- Sampaio, B.L.; Edrada-Ebel, R.; Da Costa, F.B. Effect of the Environment on the Secondary Metabolic Profile of Tithonia Diversifolia: A Model for Environmental Metabolomics of Plants. Sci. Rep. 2016, 6, 29265. [Google Scholar] [CrossRef]
- Pant, P.; Pandey, S.; Dall’Acqua, S. The Influence of Environmental Conditions on Secondary Metabolites in Medicinal Plants: A Literature Review. Chem. Biodivers. 2021, 18, e2100345. [Google Scholar] [CrossRef]
- Qiu, C.; Sun, J.; Shen, J.; Zhang, S.; Ding, Y.; Gai, Z.; Fan, K.; Song, L.; Chen, B.; Ding, Z.; et al. Fulvic Acid Enhances Drought Resistance in Tea Plants by Regulating the Starch and Sucrose Metabolism and Certain Secondary Metabolism. J. Proteom. 2021, 247, 104337. [Google Scholar] [CrossRef]
- Pereira, M.M.A.; Morais, L.C.; Marques, E.A.; Martins, A.D.; Cavalcanti, V.P.; Rodrigues, F.A.; Gonçalves, W.M.; Blank, A.F.; Pasqual, M.; Dória, J. Humic Substances and Efficient Microorganisms: Elicitation of Medicinal Plants—A Review. JAS 2019, 11, 268. [Google Scholar] [CrossRef]
- Wei, X.; Chen, J.; Gao, B.; Wang, Z. Chapter 39—Role of Controlled and Slow Release Fertilizers in Fruit Crop Nutrition. In Fruit Crops; Srivastava, A.K., Hu, C., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 555–566. ISBN 978-0-12-818732-6. [Google Scholar]
- Soil Atlas 2015|Global Soil Week. Available online: https://gsf.globalsoilweek.org/soilatlas-2015 (accessed on 14 December 2022).
- Riahi, L.; Cherif, H.; Miladi, S.; Neifar, M.; Bejaoui, B.; Chouchane, H.; Masmoudi, A.S.; Cherif, A. Use of Plant Growth Promoting Bacteria as an Efficient Biotechnological Tool to Enhance the Biomass and Secondary Metabolites Production of the Industrial Crop Pelargonium Graveolens L’Hér. under Semi-Controlled Conditions. Ind. Crops Prod. 2020, 154, 112721. [Google Scholar] [CrossRef]
- Tennakoon, P.L.K.; Rajapaksha, R.M.C.P.; Hettiarachchi, L.S.K. Tea Yield Maintained in PGPR Inoculated Field Plants despite Significant Reduction in Fertilizer Application. Rhizosphere 2019, 10, 100146. [Google Scholar] [CrossRef]
- Building a Sustainable and Food-Secure World Starts with the Right to Food. Available online: http://www.fao.org/publications/highlights-detail/en/c/1458917/ (accessed on 6 February 2023).
- Das, P.P.; Singh, K.R.; Nagpure, G.; Mansoori, A.; Singh, R.P.; Ghazi, I.A.; Kumar, A.; Singh, J. Plant-Soil-Microbes: A Tripartite Interaction for Nutrient Acquisition and Better Plant Growth for Sustainable Agricultural Practices. Environ. Res. 2022, 214, 113821. [Google Scholar] [CrossRef] [PubMed]
- Egamberdieva, D.; Shrivastava, S.; Varma, A. (Eds.) Plant-Growth-Promoting Rhizobacteria (PGPR) and Medicinal Plants; Soil Biology; Springer International Publishing: Cham, Switzerland, 2015; Volume 42, ISBN 978-3-319-13400-0. [Google Scholar]
- Ibort, P.; Imai, H.; Uemura, M.; Aroca, R. Proteomic Analysis Reveals That Tomato Interaction with Plant Growth Promoting Bacteria Is Highly Determined by Ethylene Perception. J. Plant Physiol. 2018, 220, 43–59. [Google Scholar] [CrossRef]
- Lugtenberg, B.; Kamilova, F. Plant-Growth-Promoting Rhizobacteria. Annu. Rev. Microbiol. 2009, 63, 541–556. [Google Scholar] [CrossRef] [Green Version]
- Cai, Z.; Kastell, A.; Speiser, C.; Smetanska, I. Enhanced Resveratrol Production in Vitis Vinifera Cell Suspension Cultures by Heavy Metals without Loss of Cell Viability. Appl. Biochem. Biotechnol. 2013, 171, 330–340. [Google Scholar] [CrossRef] [PubMed]
- Cappellari, L.D.R.; Santoro, M.V.; Schmidt, A.; Gershenzon, J.; Banchio, E. Induction of Essential Oil Production in Mentha x Piperita by Plant Growth Promoting Bacteria Was Correlated with an Increase in Jasmonate and Salicylate Levels and a Higher Density of Glandular Trichomes. Plant Physiol. Biochem. 2019, 141, 142–153. [Google Scholar] [CrossRef] [PubMed]
- Walker, V.; Bertrand, C.; Bellvert, F.; Moënne-Loccoz, Y.; Bally, R.; Comte, G. Host Plant Secondary Metabolite Profiling Shows a Complex, Strain-Dependent Response of Maize to Plant Growth-Promoting Rhizobacteria of the Genus Azospirillum. New Phytol. 2011, 189, 494–506. [Google Scholar] [CrossRef] [PubMed]
- Miliordos, D.-E.; Tsiknia, M.; Kontoudakis, N.; Dimopoulou, M.; Bouyioukos, C.; Kotseridis, Y. Impact of Application of Abscisic Acid, Benzothiadiazole and Chitosan on Berry Quality Characteristics and Plant Associated Microbial Communities of Vitis Vinifera L Var. Mouhtaro Plants. Sustainability 2021, 13, 5802. [Google Scholar] [CrossRef]
- Desrut, A.; Moumen, B.; Thibault, F.; Le Hir, R.; Coutos-Thévenot, P.; Vriet, C. Beneficial Rhizobacteria Pseudomonas simiae WCS417 Induce Major Transcriptional Changes in Plant Sugar Transport. J. Exp. Bot. 2020, 71, 7301–7315. [Google Scholar] [CrossRef]
- Bouyoucos, G.J. A Recalibration of the Hydrometer Method for Making Mechanical Analysis of Soils1. Agron. J. 1951, 43, 434–438. [Google Scholar] [CrossRef]
- Carter, M.R. Soil Sampling and Methods of Analysis; CRC Press: Boca Raton, FL, USA, 1993; ISBN 978-0-87371-861-5. [Google Scholar]
- Nelson, D.W.; Sommers, L.E. Total Carbon, Organic Carbon, and Organic Matter. In Methods of Soil Analysis; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 1983; pp. 539–579. ISBN 978-0-89118-977-0. [Google Scholar]
- Bremner, J.M.; Mulvaney, C.S. Nitrogen—Total. In Methods of Soil Analysis; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 1983; pp. 595–624. ISBN 978-0-89118-977-0. [Google Scholar]
- Lindsay, W.L.; Norvell, W.A. Development of a DTPA Soil Test for Zinc, Iron, Manganese, and Copper. Soil Sci. Soc. Am. J. 1978, 42, 421–428. [Google Scholar] [CrossRef]
- Massas, I.; Kalivas, D.; Ehaliotis, C.; Gasparatos, D. Total and Available Heavy Metal Concentrations in Soils of the Thriassio Plain (Greece) and Assessment of Soil Pollution Indexes. Environ. Monit. Assess. 2013, 185, 6751–6766. [Google Scholar] [CrossRef]
- Olsen, S.R. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate; U.S. Department of Agriculture: Washington, DC, USA, 1954. [Google Scholar]
- Dick, W.A.; Tabatabai, M.A. Determination of Orthophosphate in Aqueous Solutions Containing Labile Organic and Inorganic Phosphorus Compounds. J. Environ. Qual. 1977, 6, 82–85. [Google Scholar] [CrossRef]
- Bremner, J.M. Total Nitrogen. In Methods of Soil Analysis; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 1965; pp. 1149–1178. ISBN 978-0-89118-204-7. [Google Scholar]
- Eberendu, A.R.; Luta, G.; Edwards, J.A.; McAnalley, B.H.; Davis, B.; Rodriguez, S.; Henry, C.R. Quantitative Colorimetric Analysis of Aloe Polysaccharides as a Measure of Aloe vera Quality in Commercial Products. J. AOAC Int. 2005, 88, 684–691. [Google Scholar] [CrossRef] [Green Version]
- Cardarelli, M.; Rouphael, Y.; Rea, E.; Lucini, L.; Pellizzoni, M.; Colla, G. Effects of Fertilization, Arbuscular Mycorrhiza, and Salinity on Growth, Yield, and Bioactive Compounds of Two Aloe Species. Horts 2013, 48, 568–575. [Google Scholar] [CrossRef]
- Folin, O.; Ciocalteu, V. On Tyrosine and Tryptophane Determinations in Proteins. J. Biol. Chem. 1927, 73, 627–650. [Google Scholar] [CrossRef]
- Mendiburu, F.D.; Simon, R. Agricolae—Ten Years of an Open Source Statistical Tool for Experiments in Breeding, Agriculture and Biology; PeerJ Inc.: California, CA, USA, 2015. [Google Scholar]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Olk, D.C.; Dinnes, D.L.; Rene Scoresby, J.; Callaway, C.R.; Darlington, J.W. Humic Products in Agriculture: Potential Benefits and Research Challenges—A Review. J. Soils Sediments 2018, 18, 2881–2891. [Google Scholar] [CrossRef]
- Saxena, A.K.; Kumar, M.; Chakdar, H.; Anuroopa, N.; Bagyaraj, D.J. Bacillus Species in Soil as a Natural Resource for Plant Health and Nutrition. J. Appl. Microbiol. 2020, 128, 1583–1594. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.; Nadeem, S.M.; Naveed, M.; Zahir, Z.A. Potassium-Solubilizing Bacteria and Their Application in Agriculture. In Potassium Solubilizing Microorganisms for Sustainable Agriculture; Meena, V.S., Maurya, B.R., Verma, J.P., Meena, R.S., Eds.; Springer India: New Delhi, India, 2016; pp. 293–313. ISBN 978-81-322-2776-2. [Google Scholar]
- Meena: Agriculturally Important Microbes for Sustainable. Available online: https://scholar.google.com/scholar_lookup?hl=en&publication_year=2017&pages=25-61&author=N.+Kumawat&author=R.+Kumar&author=S.+Kumar&author=V.S.+Meena&isbn=%00null%00&title=Applications+in+Crop+Production+and+Protection (accessed on 29 December 2022).
- Bhaskar, R.; Xavier, L.S.E.; Udayakumaran, G.; Kumar, D.S.; Venkatesh, R.; Nagella, P. Biotic Elicitors: A Boon for the in-Vitro Production of Plant Secondary Metabolites. Plant Cell Tiss. Organ Cult. 2022, 149, 7–24. [Google Scholar] [CrossRef]
- Mahapatra, S.; Yadav, R.; Ramakrishna, W. Bacillus subtilis Impact on Plant Growth, Soil Health and Environment: Dr. Jekyll and Mr. Hyde. J. Appl. Microbiol. 2022, 132, 3543–3562. [Google Scholar] [CrossRef] [PubMed]
- Khajeeyan, R.; Salehi, A.; Movahhedi Dehnavi, M.; Farajee, H.; Kohanmoo, M.A. Growth Parameters, Water Productivity and Aloin Content of Aloe vera Affected by Mycorrhiza and PGPR Application under Different Irrigation Regimes. South Afr. J. Bot. 2022, 147, 1188–1198. [Google Scholar] [CrossRef]
- Lopez, B.R.; Tinoco-Ojanguren, C.; Bacilio, M.; Mendoza, A.; Bashan, Y. Endophytic Bacteria of the Rock-Dwelling Cactus Mammillaria Fraileana Affect Plant Growth and Mobilization of Elements from Rocks. Environ. Exp. Bot. 2012, 81, 26–36. [Google Scholar] [CrossRef]
- Di, Y.; Kui, L.; Singh, P.; Liu, L.; Xie, L.; He, L.; Li, F. Identification and Characterization of Bacillus subtilis B9: A Diazotrophic Plant Growth-Promoting Endophytic Bacterium Isolated from Sugarcane Root. J Plant Growth Regul. 2022, 41, 1–18. [Google Scholar] [CrossRef]
- Kalinowski, B.E.; Liermann, L.J.; Givens, S.; Brantley, S.L. Rates of Bacteria-Promoted Solubilization of Fe from Minerals: A Review of Problems and Approaches. Chem. Geol. 2000, 169, 357–370. [Google Scholar] [CrossRef]
- Sayyed, R.Z.; Chincholkar, S.B.; Reddy, M.S.; Gangurde, N.S.; Patel, P.R. Siderophore Producing PGPR for Crop Nutrition and Phytopathogen Suppression. In Bacteria in Agrobiology: Disease Management; Maheshwari, D.K., Ed.; Springer: Berlin, Heidelberg, 2013; pp. 449–471. ISBN 978-3-642-33639-3. [Google Scholar]
- Lv, L.; Luo, J.; Ahmed, T.; Zaki, H.E.M.; Tian, Y.; Shahid, M.S.; Chen, J.; Li, B. Beneficial Effect and Potential Risk of Pantoea on Rice Production. Plants 2022, 11, 2608. [Google Scholar] [CrossRef]
- Gilliham, M.; Dayod, M.; Hocking, B.J.; Xu, B.; Conn, S.J.; Kaiser, B.N.; Leigh, R.A.; Tyerman, S.D. Calcium Delivery and Storage in Plant Leaves: Exploring the Link with Water Flow. J. Exp. Bot. 2011, 62, 2233–2250. [Google Scholar] [CrossRef]
- Kim, Y.-C.; Glick, B.R.; Bashan, Y.; Ryu, C.-M. Enhancement of Plant Drought Tolerance by Microbes. In Plant Responses to Drought Stress: From Morphological to Molecular Features; Aroca, R., Ed.; Springer: Berlin, Heidelberg, 2012; pp. 383–413. ISBN 978-3-642-32653-0. [Google Scholar]
- Chowdhury, T.; Chowdhury, M.A.H.; Qingyue, W.; Enyoh, C.E.; Wang, W.; Khan, M.S.I. Nutrient Uptake and Pharmaceutical Compounds of Aloe vera as Influenced by Integration of Inorganic Fertilizer and Poultry Manure in Soil. Heliyon 2021, 7, e07464. [Google Scholar] [CrossRef]
- Ragel, P.; Raddatz, N.; Leidi, E.O.; Quintero, F.J.; Pardo, J.M. Regulation of K+ Nutrition in Plants. Front. Plant Sci. 2019, 10, 281. [Google Scholar] [CrossRef]
- Zhang, C.; Zhou, T.; Zhu, L.; Du, Z.; Li, B.; Wang, J.; Wang, J.; Sun, Y. Using Enzyme Activities and Soil Microbial Diversity to Understand the Effects of Fluoxastrobin on Microorganisms in Fluvo-Aquic Soil. Sci. Total Environ. 2019, 666, 89–93. [Google Scholar] [CrossRef]
- Sandhya, V.; Ali, S.Z.; Grover, M.; Reddy, G.; Venkateswarlu, B. Effect of Plant Growth Promoting Pseudomonas spp. on Compatible Solutes, Antioxidant Status and Plant Growth of Maize under Drought Stress. Plant Growth Regul. 2010, 62, 21–30. [Google Scholar] [CrossRef]
- Singh, R.P.; Jha, P.; Jha, P.N. Bio-Inoculation of Plant Growth-Promoting Rhizobacterium Enterobacter cloacae ZNP-3 Increased Resistance Against Salt and Temperature Stresses in Wheat Plant (Triticum aestivum L.). J. Plant Growth Regul. 2017, 36, 783–798. [Google Scholar] [CrossRef]
- Jiménez-Gómez, A.; García-Estévez, I.; García-Fraile, P.; Escribano-Bailón, M.T.; Rivas, R. Increase in Phenolic Compounds of Coriandrum sativum L. after the Application of a Bacillus halotolerans Biofertilizer. J. Sci. Food Agric. 2020, 100, 2742–2749. [Google Scholar] [CrossRef]
- Jiménez-Gómez, A.; García-Estévez, I.; Escribano-Bailón, M.T.; García-Fraile, P.; Rivas, R. Bacterial Fertilizers Based on Rhizobium laguerreae and Bacillus halotolerans Enhance Cichorium endivia L. Phenolic Compound and Mineral Contents and Plant Development. Foods 2021, 10, 424. [Google Scholar] [CrossRef]
- Chandrasekaran, M.; Chun, S.C.; Oh, J.W.; Paramasivan, M.; Saini, R.K.; Sahayarayan, J.J. Bacillus subtilis CBR05 for Tomato (Solanum lycopersicum) Fruits in South Korea as a Novel Plant Probiotic Bacterium (PPB): Implications from Total Phenolics, Flavonoids, and Carotenoids Content for Fruit Quality. Agronomy 2019, 9, 838. [Google Scholar] [CrossRef] [Green Version]
- Li, H.Q.; Jiang, X.W. Inoculation with Plant Growth-Promoting Bacteria (PGPB) Improves Salt Tolerance of Maize Seedling. Russ. J. Plant Physiol. 2017, 64, 235–241. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Soil Properties 1 | Sampling 2 | Treatments 3 | |||||
|---|---|---|---|---|---|---|---|
| C1 | C2 | C3 | C4 | C5 | C6 | ||
| pH | S1 S2 | 6.15 ± 0.04 cd 6.50 ± 0.10 bc | 6.45 ± 0.06 b 6.72 ± 0.20 ab | 6.79 ± 0.03 a 7.01 ± 0.04 a | 6.25 ± 0.05 c 6.16 ± 0.04 c | 6.71 ± 0.03 a 6.51 ± 0.09 bc | 6.03 ± 0.05 d 6.4 ± 0.01 bc |
| EC | S1 S2 | 84.65 ± 1.35 a 75.97 ± 12.54 ab | 103.05 ± 10.82 a 113.95 ± 19.57 a | 80.55 ± 6.61 a 60.15 ± 3.66 b | 76.10 ± 4.67 a 51.30 ± 2.02 b | 104.30 ± 20.72 a 57.95 ± 13.36 b | 127.60 ± 21.01 a 78.95 ± 6.61 ab |
| TOC | S1 S2 | 0.41 ± 0.01 b 0.51 ± 0.00 ab | 0.53 ± 0.01 a 0.55 ± 0.02 a | 0.47 ± 0.02 ab 0.48 ± 0.016 ab | 0.51 ± 0.10 ab 0.50 ± 0.03 ab | 0.52 ± 0.03 a 0.52 ± 0.01 ab | 0.53 ± 0.03 a 0.44 ± 0.02 b |
| ΤN | S1 S2 | 0.068 ± 0.001 a 0.057 ± 0.001 b | 0.058 ± 0.000 a 0.067 ± 0.000 a | 0.068 ± 0.002 a 0.062 ± 0.001 ab | 0.067 ± 0.001 a 0.061 ± 0.001 b | 0.068 ± 0.000 a 0.06 ± 0.001 b | 0.066 ± 0.00 a 0.03 ± 0.001 b |
| C/N ratio | S1 S2 | 6.14 ± 0.71 b 8.99 ± 0.43 a | 9.07 ± 0.19 a 8.14 ± 0.34 a | 6.90 ± 0.45 ab 7.70 ± 0.41 a | 7.68 ± 0.24 ab 8.17 ± 0.61 a | 7.70 ± 0.59 ab 8.77 ± 0.06 a | 8.04 ± 0.62 ab 8.03 ± 0.10 a |
| Ca | S1 S2 | 630.5 ± 10.7 b 971.5 ± 42.4 abc | 888.7 ± 149.7 a 832.5 ± 53.4 bc | 830.0 ± 15.0 b 1022.0 ± 61.1 ab | 622.5 ± 31.4 b 857.5 ± 60.9 bc | 835.00 ± 106.2 b 1073.0 ± 36.3 a | 633.5 ± 9.5 b 770.0 ± 12.7 c |
| Mg | S1 S2 | 40.00 ± 0.23 c 36.06 ± 1.08 ab | 47.9 ± 1.73 abc 47.43 ± 4.81 a | 59.65 ± 2.04 a 34.06 ± 1.31 b | 39.10 ± 1.44 c 33.02 ± 0.47 b | 53.40 ± 5.71 ab 39.93 ± 3.80 ab | 42.15 ± 2.28 bc 38.24 ± 0.76 ab |
| K | S1 S2 | 74.00 ± 0.60 d 53.0 ± 0.28 a | 109.00 ± 4.60 bc 106.20 ± 16.89 a | 142.50 ± 1.40 ab 64.00 ± 9.23 a | 87.00 ± 4.00 cd 47.00 ± 0.57 a | 167.50 ± 5.50 a 92.00 ± 27.71 a | 133.00 ± 16.70 b 84.00 ± 8.08 a |
| P | S1 S2 | 28.27 ± 5.08 cd 16.97 ± 0.96 a | 59.01 ± 7.30 ab 30.81 ± 7.56 a | 69.27 ± 1.09 a 21.63 ± 0.86 a | 20.25 ± 3.36 d 19.00 ± 3.08 a | 46.77 ± 2.19 bc 19.22 ± 0.87 a | 28.11 ± 3.82 cd 18.92 ± 4.87 a |
| Na | S1 S2 | 13.55 ± 0.09 a 23.65 ± 2.28 a | 14.80 ± 0.57 a 22.70 ± 1.32 a | 16.50 ± 0.10 a 20.00 ± 1.38 a | 16.50 ± 1.44 a 23.70 ± 1.79 a | 16.50 ± 1.79 a 16.00 ± 3.69 a | 13.36 ± 0.14 a 22.00 ± 0.69 a |
| Soil Micronutrients 1 | Sampling 2 | Treatments 3 | |||||
|---|---|---|---|---|---|---|---|
| C1 | C2 | C3 | C4 | C5 | C6 | ||
| Fe | S1 S2 | 26.75 ± 0.49 b 47.82 ± 13.70 a | 55.46 ± 10.73 a 30.99 ± 7.09 a | 34.88 ± 0.75 ab 20.69 ± 2.69 a | 33.35 ± 0.54 ab 32.03 ± 5.46 a | 30.40 ± 1.67 b 24.04 ± 1.67 a | 35.58 ± 2.62 ab 48.71 ± 5.96 a |
| Mn | S1 S2 | 9.78 ± 0.14 a 17.28 ± 3.93 a | 11.43 ± 0.57 a 30.99 ± 7.09 a | 9.58 ± 0.17 a 20.69 ± 2.69 a | 10.14 ± 0.83 a 32.03 ± 5.46 a | 9.49 ± 0.22 a 24.04 ± 1.67 a | 10.28 ± 0.62 a 25.09 ± 7.67 a |
| Zn | S1 S2 | 3.45 ± 0.05 b 4.18 ± 0.08 a | 4.29 ± 0.19 a 4.44 ± 0.13 a | 3.76 ± 0.07 ab 4.46 ± 0.19 a | 3.44 ± 0.12 b 4.20 ± 0.11 a | 3.83 ± 0.04 ab 4.11 ± 0.03 a | 3.65 ± 0.30 ab 4.48 ± 0.04 a |
| Cu | S1 S2 | 1.20 ± 0.14 b 1.90 ± 0.11 a | 1.86 ± 0.09 a 1.90 ± 0.01 a | 1.76 ± 0.03 a 1.63 ± 0.08 a | 1.32 ± 0.06 b 1.86 ± 0.11 a | 1.68 ± 0.06 a 1.80 ± 0.01 a | 1.16 ± 0.02 b 1.90 ± 0.02 a |
| Plant Growth 1 | Treatments 2 | |||||
|---|---|---|---|---|---|---|
| C1 | C2 | C3 | C4 | C5 | C6 | |
| Height | 52.50 ± 1.90 a | 52.12 ± 2.53 a | 54.00 ± 1.05 a | 51.79 ± 1.85 a | 52.00 ± 1.48 a | 52.40 ± 1.59 a |
| Width | 57.83 ± 2.87 a | 58.41 ± 3.44 a | 55.94 ± 1.67 a | 55.75 ± 3.38 a | 56.28 ± 1.49 a | 52.23 ± 3.91 a |
| NOL | 14.70 ± 0.78 a | 16.17 ± 0.83 a | 16.89 ± 0.69 a | 16.50 ± 0.66 a | 16.44 ± 0.50 a | 15.65 ± 0.65 a |
| NOO | 6.83 ± 1.52 a | 6.66 ± 1.79 a | 7.44 ± 1.82 a | 6.83 ± 1.58 a | 7.05 ± 1.33 a | 6.70 ± 1.65 a |
| Rind Minerals 1 | Treatments 2 | |||||
|---|---|---|---|---|---|---|
| C1 | C2 | C3 | C4 | C5 | C6 | |
| N | 7.52 ± 0.65 b | 8.69 ± 0.48 ab | 10.3 ± 0.35 a | 8.63 ± 0.38 ab | 9.61 ± 0.27 ab | 10.03 ± 0.73 a |
| Ca | 18.36 ± 0.82 bc | 23.75 ± 0.40 a | 18.68 ± 1.53 bc | 21.6 ± 0.63 ab | 16.57 ± 0.65 c | 19.20 ± 1.67 bc |
| Mg | 5.71 ± 0.19 b | 7.61 ± 0.21 a | 6.50 ± 0.29 ab | 6.56 ± 0.37 ab | 5.20 ± 0.53 b | 6.39 ± 0.50 ab |
| K | 14.68 ± 0.92 a | 13.43 ± 1.48 a | 19.45 ± 2.56 a | 12.42 ± 1.04 a | 15.77 ± 1.43 a | 19.36 ± 2.04 a |
| P | 1.07 ± 0.10 b | 1.06 ± 0.13 b | 1.84 ± 0.18 ab | 1.42 ± 0.19 ab | 2.44 ± 0.48 a | 2.01 ± 0.39 ab |
| Na | 7.04 ± 0.60 a | 5.52 ± 0.40 ab | 3.63 ± 0.74 b | 6.07 ± 0.60 ab | 3.86 ± 0.77 b | 6.73 ± 0.17 a |
| Fe | 20.50 ± 0.24 b | 36.41 ± 3.17 a | 31.56 ± 0.87 a | 34.44 ± 3.73 a | 32.03 ± 1.88 a | 35.18 ± 1.78 a |
| Mn | 46.34 ± 6.59 c | 94.49 ± 10.55 ab | 38.72 ± 3.89 c | 107.67 ± 22.8 a | 51.27 ± 4.02 bc | 102.82 ± 9.82 a |
| Zn | 17.46 ± 0.77 ab | 17.08 ± 2.08 ab | 15.41 ± 2.62 ab | 22.69 ± 2.76 a | 13.06 ± 2.25 b | 23.34 ± 1.88 a |
| Cu | 2.66 ± 0.59 a | 1.70 ± 0.2 a | 2.84 ± 0.34 a | 2.21 ± 0.17 a | 1.94 ± 0.18 a | 2.10 ± 0.2 a |
| Gel Minerals 1 | Treatments 2 | |||||
|---|---|---|---|---|---|---|
| C1 | C2 | C3 | C4 | C5 | C6 | |
| Ca | 40.57 ± 1.76 b | 48.36 ± 2.16 a | 27.62 ± 1.14 c | 43.13 ± 0.24 ab | 38.21 ± 0.47 b | 31.58 ± 2.26 c |
| Mg | 5.21 ± 0.19 b | 6.65 ± 0.25 a | 5.28 ± 0.20 b | 6.02 ± 0.11 a | 5.00 ± 0.18 b | 5.20 ± 0.16 b |
| K | 14.37 ± 1.23 b | 8.72 ± 1.15 c | 35.76 ± 2.24 a | 6.91 ± 0.70 c | 9.97 ± 0.53 bc | 12.11 ± 0.92 bc |
| P | 0.68 ± 0.07 c | 0.55 ± 0.08 c | 1.11 ± 0.08 b | 0.77 ± 0.02 bc | 1.79 ± 0.18 a | 0.78 ± 0.08 bc |
| Na | 10.17 ± 0.36 ab | 11.10 ± 0.33 a | 8.97 ± 0.14 b | 10.74 ± 0.52 a | 6.72 ± 0.17 c | 10.45 ± 0.38 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nikolaou, C.N.; Chatziartemiou, A.; Tsiknia, M.; Karyda, A.G.; Ehaliotis, C.; Gasparatos, D. Calcium- and Magnesium-Enriched Organic Fertilizer and Plant Growth-Promoting Rhizobacteria Affect Soil Nutrient Availability, Plant Nutrient Uptake, and Secondary Metabolite Production in Aloe vera (Aloe barbadensis Miller) Grown under Field Conditions. Agronomy 2023, 13, 482. https://doi.org/10.3390/agronomy13020482
Nikolaou CN, Chatziartemiou A, Tsiknia M, Karyda AG, Ehaliotis C, Gasparatos D. Calcium- and Magnesium-Enriched Organic Fertilizer and Plant Growth-Promoting Rhizobacteria Affect Soil Nutrient Availability, Plant Nutrient Uptake, and Secondary Metabolite Production in Aloe vera (Aloe barbadensis Miller) Grown under Field Conditions. Agronomy. 2023; 13(2):482. https://doi.org/10.3390/agronomy13020482
Chicago/Turabian StyleNikolaou, Christina N., Artemios Chatziartemiou, Myrto Tsiknia, Asimina Georgia Karyda, Constantinos Ehaliotis, and Dionisios Gasparatos. 2023. "Calcium- and Magnesium-Enriched Organic Fertilizer and Plant Growth-Promoting Rhizobacteria Affect Soil Nutrient Availability, Plant Nutrient Uptake, and Secondary Metabolite Production in Aloe vera (Aloe barbadensis Miller) Grown under Field Conditions" Agronomy 13, no. 2: 482. https://doi.org/10.3390/agronomy13020482
APA StyleNikolaou, C. N., Chatziartemiou, A., Tsiknia, M., Karyda, A. G., Ehaliotis, C., & Gasparatos, D. (2023). Calcium- and Magnesium-Enriched Organic Fertilizer and Plant Growth-Promoting Rhizobacteria Affect Soil Nutrient Availability, Plant Nutrient Uptake, and Secondary Metabolite Production in Aloe vera (Aloe barbadensis Miller) Grown under Field Conditions. Agronomy, 13(2), 482. https://doi.org/10.3390/agronomy13020482