
High-Voltage Electrostatic Fields Adversely Affect the Performance of Diamondback Moths over Five Consecutive Generations



Abstract
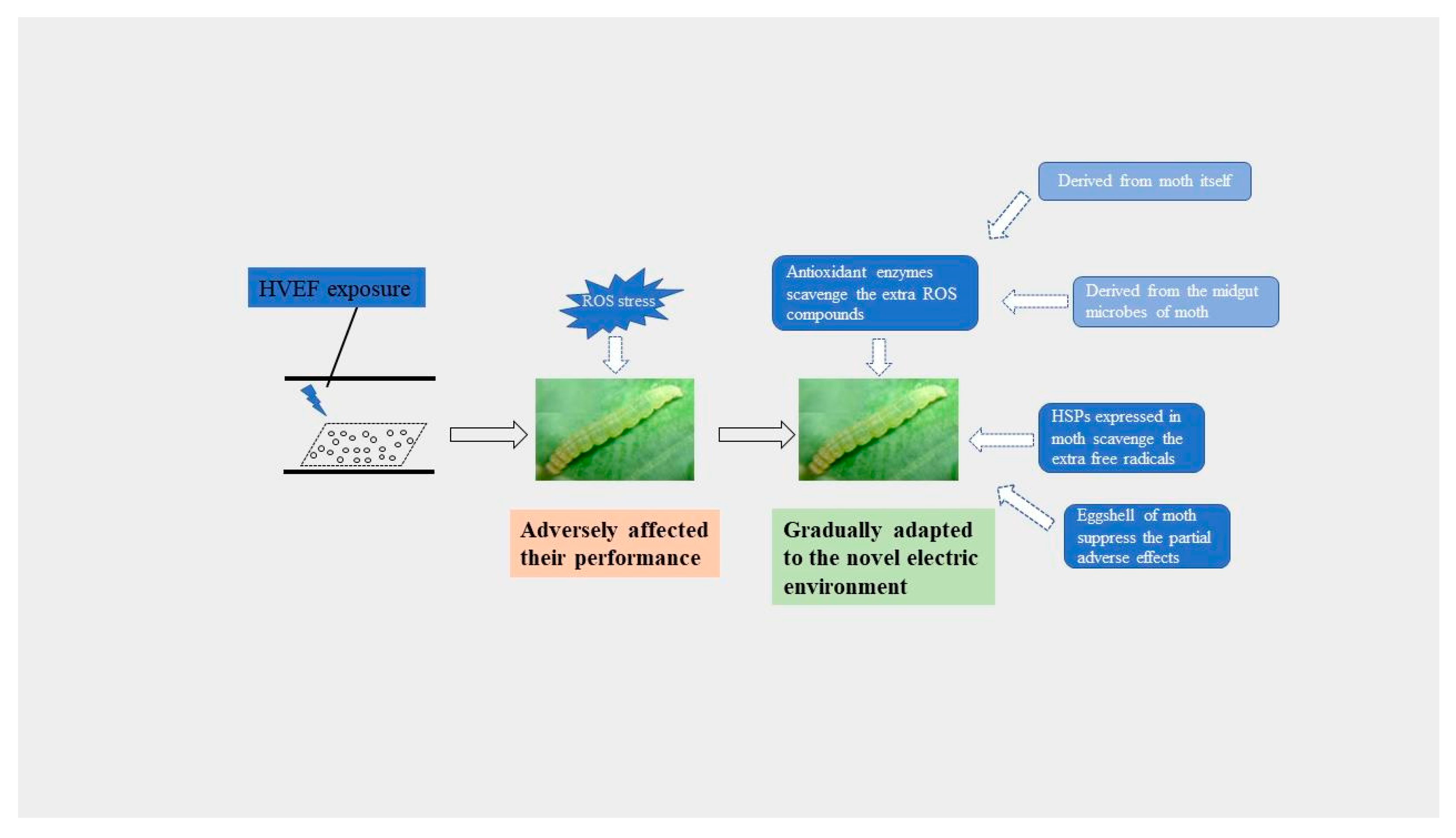
:1. Introduction
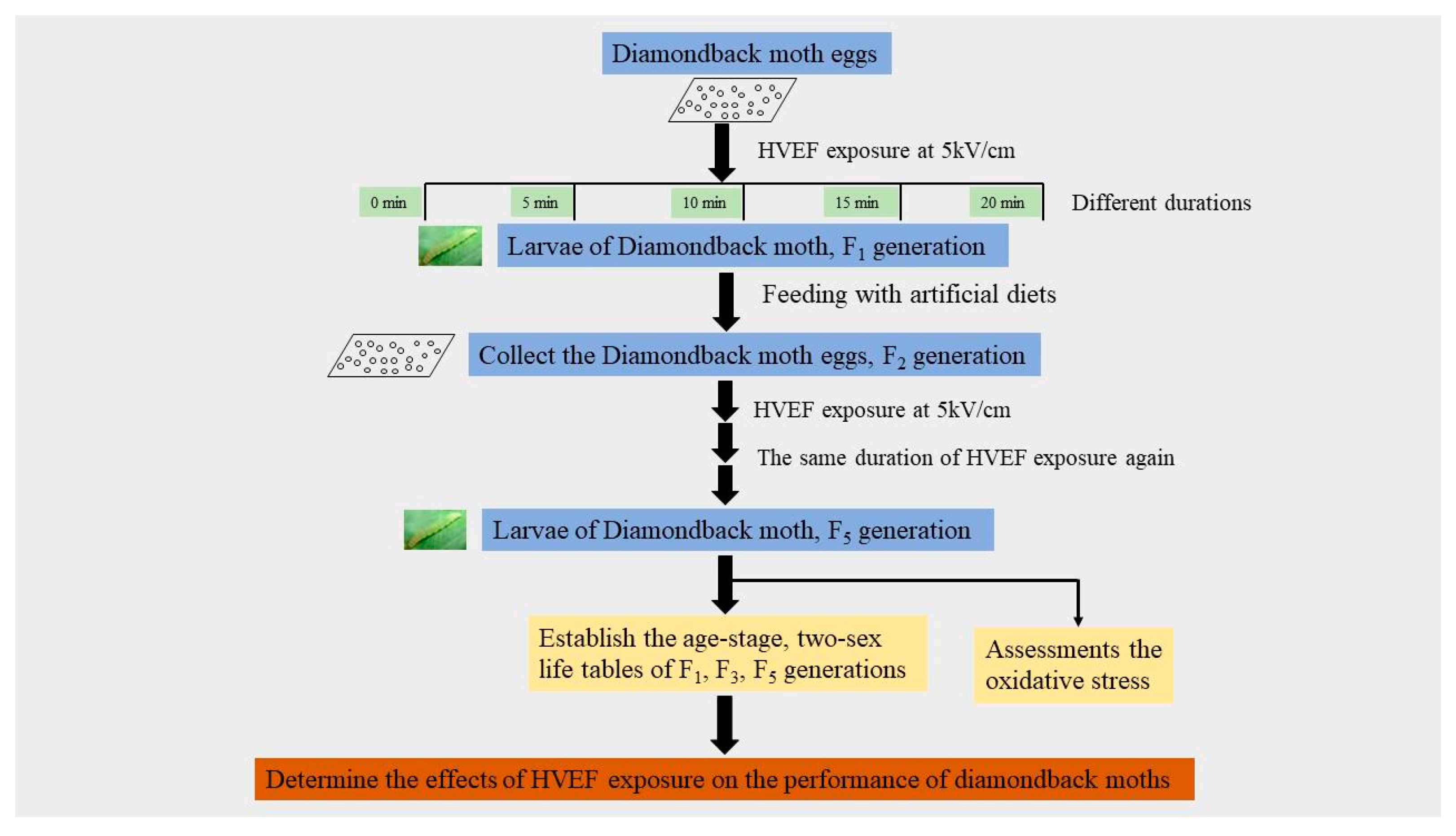
2. Materials and Methods
2.1. Insect Specimens and Rearing Conditions
2.2. HVEF Treatment
2.3. Life Table Analysis
2.4. Oxidative Stress Assessment
2.5. Statistical Analysis
3. Results
3.1. HVEF Treatment Adversely Affected Net Reproduction Rate (R0) of P. xylostella
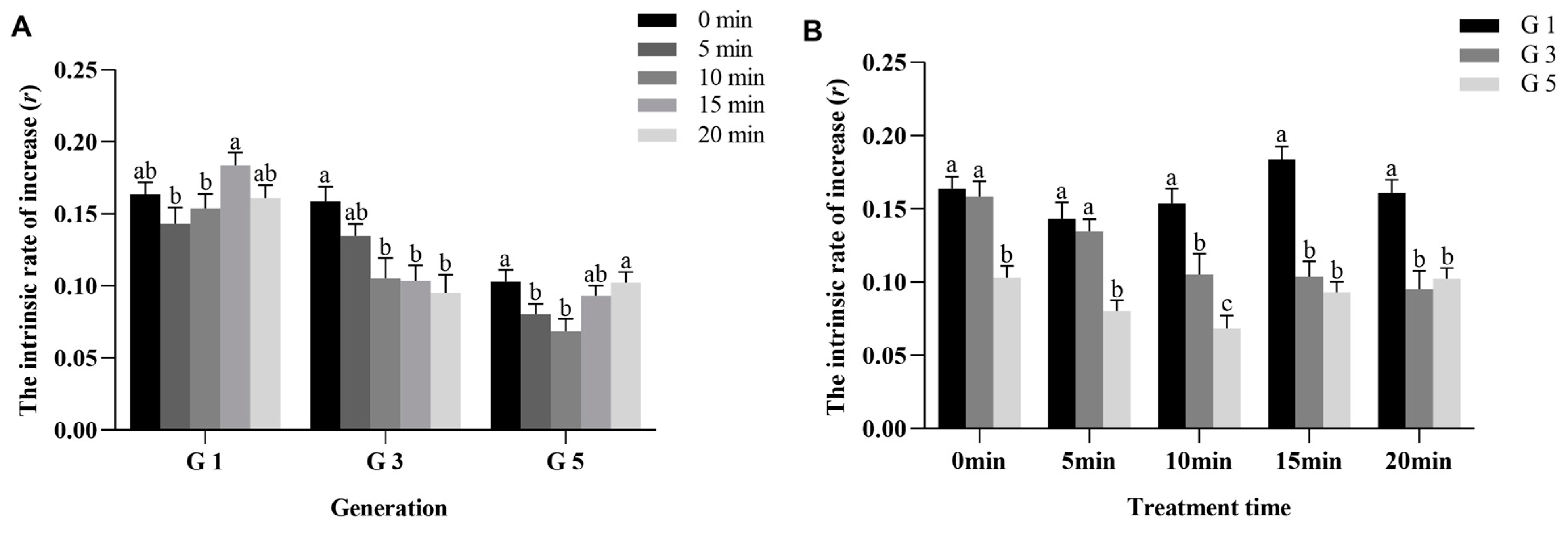
3.2. HVEF Stress Significantly Affected the r of P. xylostella in All Generations
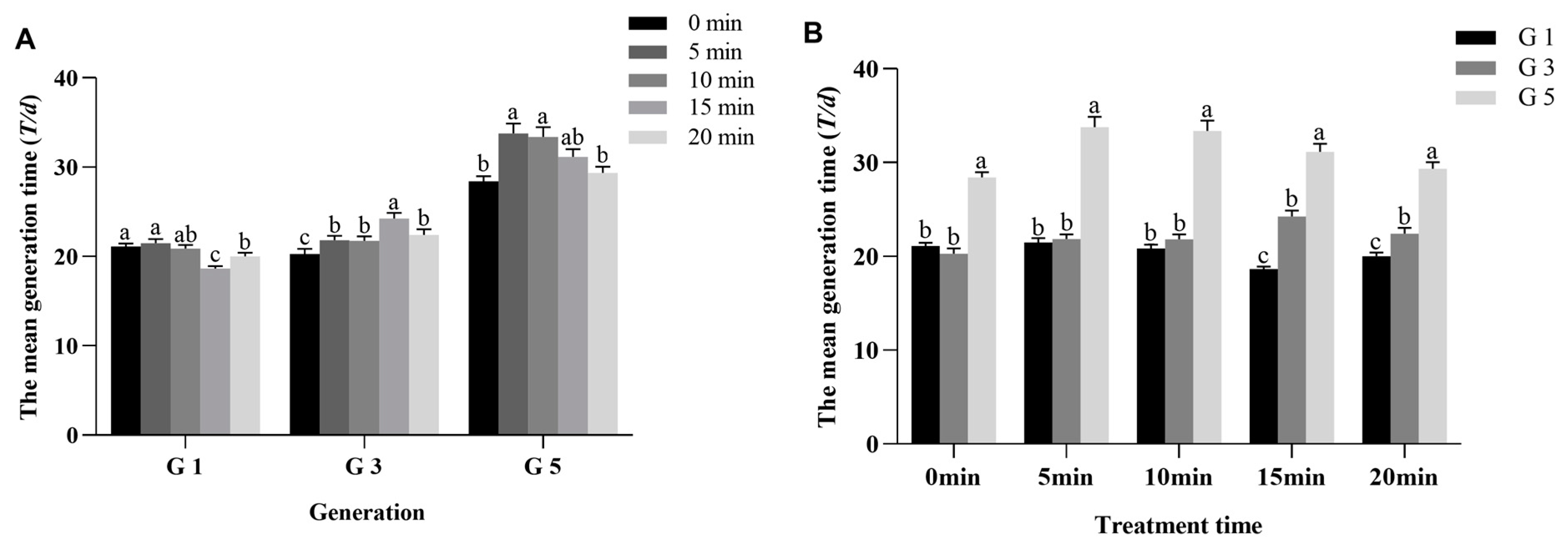
3.3. HVEF Treatment Gradually Prolonged the T of P. xylostella
3.4. HVEF Stress Exerted a Suppressive Effect on Population Growth of P. xylostella
3.5. Effect of HVEF Exposure on Survival and Oviposition Parameters of P. xylostella
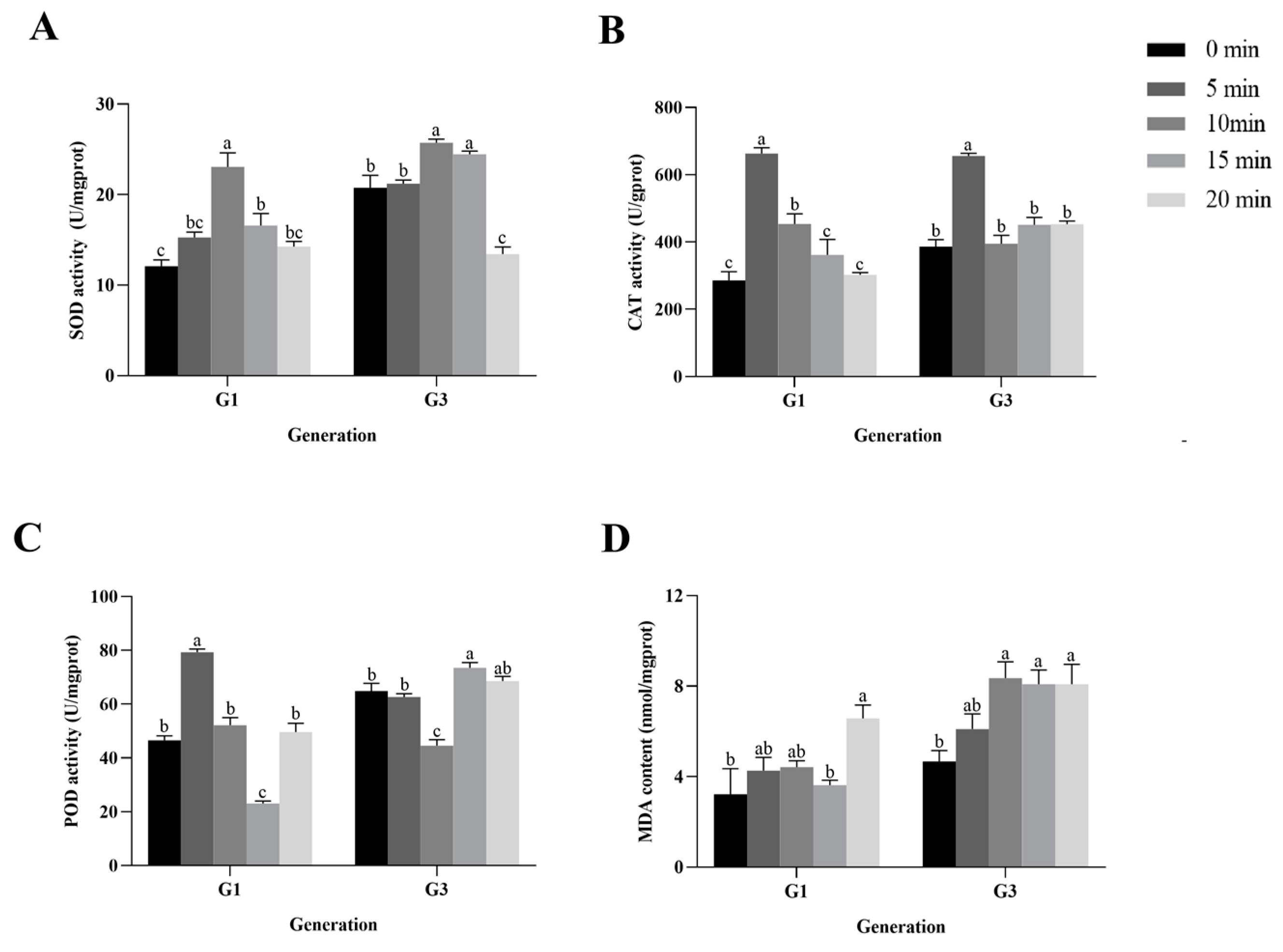
3.6. HVEF Exposure Led to Significant Oxidative Damage to Fourth-Instar P. xylostella
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Furlong, M.J.; Wright, D.J.; Dosdall, L.M. Diamondback moth ecology and management: Problems, progress, and prospects. Annu. Rev. Entomol. 2013, 58, 517–541. [Google Scholar] [CrossRef]
- Gautam, M.P.; Singh, H.; Kumar, S.; Kumar, V.; Singh, S.N. Diamondback moth, Plutella xylostella (Linnaeus) (Insecta: Lepidoptera: Plutellidae) a major insect of cabbage in India: A review. J. Entomol. Zool. Stud. 2018, 6, 1394–1399. [Google Scholar]
- Jaleel, W.; Saeed, S.; Naqqash, M.N.; Sial, M.U.; Ali, M.; Zaka, S.M.; Sarwar, Z.M.; Ishtiaq, M.; Qayyum, M.A.; Aine, Q.U.; et al. Effects of temperature on baseline susceptibility and stability of insecticide resistance against Plutella xylostella (Lepidoptera: Plutellidae) in the absence of selection pressure. Saudi J. Biol. Sci. 2020, 27, 1–5. [Google Scholar] [CrossRef]
- Zalucki, M.P.; Shabbir, A.; Silva, R.; Adamson, D.; Shu-Sheng, L.; Furlong, M.J. Estimating the economic cost of one of the world’s major insect pests, Plutella xylostella (Lepidoptera: Plutellidae): Just how long is a piece of string? J. Econ. Entomol. 2012, 105, 1115–1129. [Google Scholar] [CrossRef]
- Wang, J.; Zheng, X.B.; Yuan, J.J.; Wang, S.Y.; Xu, B.Y.; Wang, S.L.; Zhang, Y.J.; Wu, Q.J. Insecticide resistance monitoring of the diamondback moth (Lepidoptera: Plutellidae) populations in China. J. Econ. Entomol. 2021, 114, 1282–1290. [Google Scholar] [CrossRef] [PubMed]
- You, S.; Liu, Z.; Xiong, L.; Liao, J.; He, W.Y.; You, M.S. Advances in research on the resistance of Plutella xylostella to Bacillus thuringiensis (Bt). Chin. J. Appl. Entomoloy 2018, 55, 951–962. [Google Scholar]
- Li, Z.Y.; Xiao, Y.; Wu, Q.J.; Shen, A.D.; Wang, X.L.; Zhang, J.M.; Feng, X. Progress in research on diamondback moth (Plutella xylostella) outbreaks and the management of resistance in this pest. Chin. J. Appl. Entomol. 2020, 57, 549–567. [Google Scholar]
- Talekar, N.S.; Shelton, A.M. Biology, ecology, and management of the diamondback moth. Annu. Rev. Entomol. 2003, 38, 275–301. [Google Scholar] [CrossRef]
- Luo, Y.J.; Wu, W.W.; Yang, Z.B.; Pu, E.T.; Guo, Z.X.; Yin, K.S.; He, C.X. Advances in insecticide resistance of diamondback moth (Plutella xylostella L.) in Yunnan. J. Yunnan Univ. 2008, 30, 178–182. [Google Scholar]
- Santos, V.C.; Haa, D.S.; Da, S.J.; Mjdc, D.F. Insecticide resistance in populations of the diamondback moth, Plutella xylostella (L.) (Lepidoptera: Plutellidae), from the state of Pernambuco, Brazil. Neotrop. Entomol. 2011, 40, 264–270. [Google Scholar] [CrossRef] [Green Version]
- Gao, D.L.; Liu, Y.; Wang, L.L.; Xu, N.N.; Zhuang, Z.G.; Xu, Y.; Hu, Z.J.; Zhuang, Z.X. Insecticides against resistance population of Plutella xylostella: Toxicity analysis and field control efficacy. Chin. Agric. Sci. Bull. 2018, 34, 145–149. [Google Scholar]
- Bai, Y.; Hu, Y. Original mechanism of biological effects of electrostatic field on crop seeds. Trans. Chin. Soc. Agric. Eng. 2003, 19, 49–51. [Google Scholar] [CrossRef]
- Wu, C.Y.; Zhang, L.; Zhen, S. Biological effect mechanism and application of high voltage electrostatic field on animal body. Prog. Vet. Med. 2004, 25, 7–9. [Google Scholar] [CrossRef]
- Lee, J.Y.; Kang, S.W.; Yoon, C.S.; Kim, J.J.; Choi, D.R.; Kim, S.W. Verticillium lecanii spore formulation using UV protectant and wetting agent and the biocontrol of cotton aphids. Biotechnol. Lett. 2006, 28, 1041–1045. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.S.; Sun, H.; Liang, C.Y.; Yi, T.G.; Sun, F. Effect of high voltage electrostatic field on physiological and biochemistry indexes in developmental mice. J. Jilin Univ. 2005, 31, 839–842. [Google Scholar] [CrossRef]
- Wang, G.; Huang, J.; Gao, W.; Lu, J.; Li, J.; Liao, R.; Jaleel, C.A. The effect of high-voltage electrostatic field (HVEF) on aged rice (Oryza sativa L.) seeds vigor and lipid peroxidation of seedlings. J. Electrost. 2009, 67, 759–764. [Google Scholar] [CrossRef]
- Lu, K.; Song, Y.; Zeng, R. The role of cytochrome P450-mediated detoxification in insect adaptation to xenobiotics. Curr. Opin. Insect Sci. 2021, 43, 103–107. [Google Scholar] [CrossRef]
- Cakmak, T.; Cakmak, Z.E.; Dumlupinar, R.; Tekinay, T. Analysis of apoplastic and symplastic antioxidant system in shallot leaves: Impacts of weak static electric and magnetic field. J. Plant Physiol. 2012, 169, 1066–1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, J.; Cao, Z.; Yang, J.; Zhao, H.; Pan, W. Effects of static electric fields on growth and development of wheat aphid Sitobion aveanae (Hemiptera: Aphididae) through multiple generations. Electromagn. Biol. Med. 2016, 35, 1–7. [Google Scholar] [CrossRef]
- Cao, Z.; Li, G.Y.; He, J.; Zhao, H.Y.; Piyaratne, M.; Hu, Z.Q.; Hu, X.S. Effects of high voltage electrostatic fields on protective enzyme activity in wheat plants and on the population dynamics of Sitobion avenae Fabricius (Hemiptera: Aphididae). Acta Ecol. Sin. 2016, 36, 1001–1009. [Google Scholar]
- Luo, K.; Luo, C.; Li, G.Y.; Xin, J.Y.; Gao, R.; Hu, Z.Q.; Zhang, G.S.; Zhao, H.Y. High-voltage electrostatic field-induced oxidative stress: Characterization of the physiological effects in Sitobion avenae (Hemiptera: Aphididae) across multiple generations. Bioelectromagnetics 2019, 40, 52–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.L.; Zhao, B.G.; Xia, L.Q.; Xu, J.T. Effects of Bombyx mori eggs under the treatment of high voltage electrostatic field on dietary efficiency and larva growth and development. J. Shandong Agric. Univ. 2006, 37, 505–508. [Google Scholar]
- Diebolt, J.R. The influence of electrostatic and magnetic fields on mutation in Drosophila melanogaster spermatozoa. Mutat. Res. 1978, 57, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Harutyunyan, H.; Artsruni, G. Biological effects of static electric field: Plasma/serum proteome analysis of rats. J. Bioelectr. 2013, 32, 79–94. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.Q.; Miao, S.; Luo, K.J. Mass rearing techniques of diamondback moth. Entomol. Knowl. 2001, 38, 68–70. [Google Scholar] [CrossRef]
- Chi, H.; Liu, H. Two new methods for the study of insect population ecology. Bull. Inst. Zool. Acad. Sin. 1985, 24, 225–240. [Google Scholar]
- Chi, H.; Fu, J.W.; You, M.S. Age-stage, two-sex life table and its application in population ecology and integrated pest management. Acta Entomol. Sin. 2019, 62, 255–262. [Google Scholar] [CrossRef]
- Li, G.Y.; Gao, R.; He, J.; Cao, Z.; Hu, Z.Q.; Hu, X.S.; Zhao, H.Y. Effect of high-voltage electrostatic field (HVEF) on the growth and fecundity of Sitobion avenae Fabricius (Hemiptera: Aphididae). Acta Ecol. Sin. 2016, 36, 3987–3994. [Google Scholar]
- Luo, K.; Cao, Z.; Gao, R.; He, J.; Li, G.Y.; Gao, H.H.; Zhang, G.S.; Zhao, H.Y. Direct exposure of wheat seeds to high-voltage electrostatic fields adversely affects the performance of Sitobion avenae (Hemiptera: Aphididae). J. Econ. Entomol. 2016, 109, 2418–2423. [Google Scholar] [CrossRef]
- Marchioro, C.A.; Foerster, L.A. Development and survival of the diamondback moth, Plutella xylostella (L.) (Lepidoptera: Yponomeutidae) as a function of temperature: Effect on the number of generations in tropical and subtropical regions. Neotrop. Entomol. 2011, 40, 533–541. [Google Scholar]
- Zheng, Z.S.; Xing, K.; Zhao, F.; Wang, J.M.; Wen, X.D.; Yang, F. Effects of short hot stress and its duration on longevity and fecundity of diamondback moth adults. China Plant Prot. 2015, 35, 15–20. [Google Scholar]
- Luo, L.Y.; Zhang, X.F.; Wei, X.Q.; Zhang, K.; Weng, Q.F. Effect of 60Co-γ radiation on inherited sterility of Plutella xylostella (Lepidoptera: Plutellidae). J. Northwest AF Univ. 2018, 46, 149–154. [Google Scholar] [CrossRef]
- Li, X.; Zhang, K.; Deng, Y.K.; He, R.K.; Zhang, X.F.; Zhong, G.H.; Hu, Q.B.; Weng, Q.F. Effects of 60Co-γ radiation on testis physiological aspects of Plutella xylostella (Linnaeus). Ecotoxicol. Environ. Saf. 2019, 169, 937–943. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Seunghwan, L.; Yikweon, J. Macroevolutionary patterns in the Aphidini Aphids (Hemiptera: Aphididae): Diversification, host association, and biogeographic origins. PLoS ONE 2011, 6, e24749. [Google Scholar] [CrossRef]
- Hori, T.; Inoue, N.; Suzuki, H.; Harakawa, S. Exposure to 50 Hz electric fields reduces stress-induced glucocorticoid levels in BALB/c mice in a kV/m- and duration-dependent manner. Bioelectromagnetics 2015, 36, 302–308. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Park, J.; Noh, G.; Park, S.; Noh, K.; Kwon, I.; Ahn, S. Effects of moderate intensity static magnetic fields on osteoclastic differentiation in mouse bone marrow cells. Bioelectromagnetics 2018, 39, 394–404. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Wang, Z.; Zhang, L.; Li, Z.; Tian, X.; Fang, J.; Lu, Q.; Zhang, X. Cellular ATP levels are affected by moderate and strong static magnetic fields. Bioelectromagnetics 2018, 39, 352–360. [Google Scholar] [CrossRef]
- Hu, L.F.; Qian, A.R.; Yang, P.F.; Zhang, W.; Xie, L.; Shang, P. Effects of moderate static magnetic field on cell proliferation and cell cycle of leukem ia cells. J. Fourth Mil. Med. Univ. 2009, 30, 397–400. [Google Scholar] [CrossRef]
- Cunha, C.; Panseri, S.; Marcacci, M.; Tampieri, A. Evaluation of the effects of a moderate intensity static magnetic field application on human Osteoblast-Like cells. Am. J. Biomed. Eng. 2012, 2, 263–268. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.F.; Chen, J.; Wang, Y.G.; Du, H.L. Optimization of condition for improving sorghum seed vigor by high voltage electric field. Trans. CSAE 2015, 31, 253–259. [Google Scholar] [CrossRef]
- Gruijl, F.; Longstreth, A.J.; Norval, B.M.; Cullen, C.; Slaper, D.H. Health effects from stratospheric ozone depletion and interactions with climate change. Photochem. Photobiol. Sci. 2003, 2, 16–28. [Google Scholar] [CrossRef]
- Zhaorigetu, S.; Yanaka, N.; Sasaki, M.; Watanabe, H.; Kato, N. Inhibitory effects of silk protein, sericin on UVB-induced acute damage and tumor promotion by reducing oxidative stress in the skin of hairless mouse. J. Photochem. Photobiol. B Biol. 2003, 71, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.Y.; Zhang, C.Y.; Lei, C.L. A proteomic analysis of Helicoverpa armigera adults after exposure to UV light irradiation. J. Insect Physiol. 2010, 56, 405–411. [Google Scholar] [CrossRef] [PubMed]
- Ute, W.; Esser, P.R.; Birgit, S.H.; Martin, S.F.; Jürgen, L.; Schempp, C.M. UVB-induced DNA damage, generation of reactive oxygen species, and inflammation are effectively attenuated by the flavonoid luteolin in vitro and in vivo. Free. Radic. Biol. Med. 2015, 50, 1081–1093. [Google Scholar] [CrossRef]
- Packer, L.; Wirtz, K.W.A. Signalling Mechanisms—from Transcription Factors to Oxidative Stress; Springer: Berlin/Heidelberg, Germany, 1995. [Google Scholar]
- Wang, M.L.; Zhu, W.; Zhang, J.H.; Xu, A.; Wang, H.T.; Li, X.; Zhou, G.L.; Zhang, H.; Qin, Q.L. Oxidative damage is one of the most important causes of death of Thitarodes xiaojinensis (Lepidoptera: Hepialidae) larvae under heat stress. Acta Entomol. Sin. 2014, 57, 769–776. [Google Scholar] [CrossRef]
- Nguyen, T.; Michaud, D.; Cloutier, C. A proteomic analysis of the aphid Macrosiphum euphorbiae under heat and radiation stress. Insect Biochem. Mol. Biol. 2009, 39, 20–30. [Google Scholar] [CrossRef]
- Dharshini, L.C.P.; Vishnupriya, S.; Sakthivel, K.M.; Rasmi, R.R. Oxidative stress responsive transcription factors in cellular signalling transduction mechanisms. Cell. Signal. 2020, 72, 109670. [Google Scholar] [CrossRef]
- Blagojevi, D.P.; Grubor-Laji, G. Multifunctionality of antioxidant system in insects. Arch. Biol. Sci. 2000, 52, 185–194. [Google Scholar]
- Ray, G.; Batra, S.; Shukla, N.K.; Suryanarayan, D.; Raina, V.; Ashok, S.; Husain, S.A. Lipid peroxidation, free radical production and antioxidant status in breast cancer. Breast Cancer Res. Treat. 2000, 59, 163–170. [Google Scholar] [CrossRef]
- Li, X.; Luo, L.; Karthi, S.; Zhang, K.; Luo, J.J.; Hu, Q.B.; Weng, Q.F. Effects of 200 Gy 60Co-γ radiation on the regulation of antioxidant enzymes, Hsp70 genes, and serum molecules of Plutella xylostella (Linnaeus). Molecules 2018, 23, 1011. [Google Scholar] [CrossRef] [Green Version]
- Xia, X.F.; Lin, H.L.; Zheng, D.D.; Guang, Y.; Sheng, Y.M. Identification and expression patterns of heat shock protein genes in the diamondback moth, Plutella xylostella (Lepidoptera: Yponomeutidae). Acta Entomol. Sin. 2013, 56, 457–464. [Google Scholar] [CrossRef]
- Alkhedir, H.; Karlovsky, P.; Mashaly, M.A.A.; Vidal, S. Phylogenetic relationships of the symbiotic bacteria in the aphid Sitobion avenae (Hemiptera: Aphididae). Environ. Entomol. 2015, 44, 1358–1366. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Luo, K.; Meng, L.Q.; Wan, B.; Zhao, H.Y.; Hu, Z.Q. Ecological impact of a secondary bacterial symbiont on the clones of Sitobion avenae (Fabricius) (Hemiptera: Aphididae). Sci. Rep. 2017, 7, 40754. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Meng, J.Y.; Su, L.; Zhang, C.Y. Cloning of heat shock protein gene MpHsp70 and its expression analysis under UV-B stress in Myzus persicae (Hemiptera: Aphididae). Acta Entomol. Sin. 2019, 62, 133–140. [Google Scholar] [CrossRef]
- Ding, C.Y.; Zhao, L.; Liu, S.; Li, M.Y. Molecular characterization of heat shock protein 70 (HSP70) genes from Pieris rapae (Lepidoptera: Pieridae) and their responses to insecticide stress. Acta Entomol. Sin. 2021, 64, 1407–1416. [Google Scholar] [CrossRef]
- Liu, F.; Meng, J.Y.; Su, L.; Zhang, C.Y. Cloning of the vitellogenin receptor gene and its expression under UV-A stress in Ostrinia furnacalis. Chin. J. Appl. Entomol. 2021, 58, 96–107. [Google Scholar]
- Seal, J.R.; Havrilla, C.M.; Porter, N.A.; Hachey, D.L. Analysis of unsaturated compounds by Ag+ coordination ionspray mass spectrometry: Studies of the formation of the Ag+/lipid complex. J. Am. Soc. Mass Spectrom. 2003, 14, 872–880. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.J.; Niu, C.Y. Influence of ultraviolet light irradiation on HMG-CoA reductase, vitellogenin mRNA expression and oviposition of Helicoverpa armigera. J. Huazhong Agric. Univ. 2014, 33, 46–50. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Statistic | Generation | 0 min | 5 min | 10 min | 15 min | 20 min |
|---|---|---|---|---|---|---|
| λ | G1 | 1.180 ± 0.006 bXY | 1.164 ± 0.009 bX | 1.192 ± 0.011 aX | 1.223 ± 0.004 abX | 1.190 ± 0.013 abX |
| G3 | 1.208 ± 0.011 aX | 1.178 ± 0.001 bX | 1.160 ± 0.009 bY | 1.155 ± 0.007 bY | 1.121 ± 0.009 cY | |
| G5 | 1.156 ± 0.010 aY | 1.104 ± 0.003 bY | 1.092 ± 0.005 bZ | 1.137 ± 0.008 aY | 1.139 ± 0.008 aY | |
| dt | G1 | 4.191 ± 0.116 abXY | 4.600 ± 0.237 aY | 3.974 ± 0.212 abY | 3.450 ± 0.053 bY | 4.025 ± 0.280 abY |
| G3 | 3.686 ± 0.167 cY | 4.230 ± 0.024 bcY | 4.706 ± 0.236 bY | 4.832 ± 0.201 bX | 6.101 ± 0.388 aX | |
| G5 | 4.819 ± 0.284 bX | 7.014 ± 0.196 aX | 7.944 ± 0.417 aX | 5.415 ± 0.286 bX | 5.341 ± 0.277 bX |
| Generation | Treatment Time (min) | Adult Preoviposition Period (APOP) (d) | Oviposition Period (OP) (d) | Oviposition Days (OD) (d) | Eggs per Day during Oviposition Period | Fecundity per Female |
|---|---|---|---|---|---|---|
| G1 | 0 | 0.77 ± 0.184 aX | 5.66 ± 0.502 aY | 5.23 ± 0.440 aX | 18.866 ± 1.680 abX | 91.714 ± 7.650 aX |
| 5 | 1.19 ± 0.184 aX | 4.83 ± 0.557 aZ | 4.25 ± 0.501 aY | 16.550 ± 2.272 abX | 74.125 ± 10.856 abX | |
| 10 | 1.13 ± 0.229 aY | 4.30 ± 0.379 aY | 3.83 ± 0.335 aY | 13.339 ± 1.715 bX | 57.957 ± 8.162 bX | |
| 15 | 1.24 ± 0.297 aY | 4.51 ± 0.330 aZ | 4.09 ± 0.291 aY | 22.432 ± 2.099 aX | 95.378 ± 7.606 aX | |
| 20 | 1.04 ± 0.217 aX | 5.12 ± 0.435 aY | 5.23 ± 0.440 aX | 18.145 ± 1.613 abX | 84.049 ± 7.580 abX | |
| G3 | 0 | 1.42 ± 0.433 aX | 6.36 ± 0.526 aY | 5.28 ± 0.457 aX | 13.504 ± 1.281 aY | 84.167 ± 8.912 aX |
| 5 | 2.09 ± 0.401 aX | 7.84 ± 0.709 aY | 5.98 ± 0.504 aXY | 10.156 ± 1.141 abY | 68.911 ± 6.734 abX | |
| 10 | 2.22 ± 0.462 aXY | 5.70 ± 0.658 aY | 4.91 ± 0.579 aXY | 9.140 ± 1.329 abX | 56.348 ± 10.946 abX | |
| 15 | 2.52 ± 0.862 aX | 7.52 ± 0.996 aY | 5.87 ± 0.695 aX | 11.087 ± 2.087 abY | 66.565 ± 9.159 abX | |
| 20 | 1.82 ± 0.440 aX | 5.73 ± 0.715 aXY | 4.64 ± 0.605 aX | 7.608 ± 0.885 bZ | 45.591 ± 8.294 bY | |
| G5 | 0 | 0.25 ± 0.083 bX | 8.36 ± 0.853 aX | 6.57 ± 0.791 aX | 13.253 ± 1.609 aY | 105.143 ± 13.774 aX |
| 5 | 1.33 ± 0.211 bX | 10.52 ± 1.133 aX | 7.24 ± 0.828 aX | 8.446 ± 0.857 aY | 79.476 ± 9.600 aX | |
| 10 | 2.95 ± 0.671 aX | 7.50 ± 0.947 aX | 5.70 ± 0.758 aX | 8.688 ± 1.256 aX | 64.500 ± 9.876 aX | |
| 15 | 0.60 ± 0.212 bY | 9.77 ± 0.956 aX | 7.27 ± 0.733 aX | 9.467 ± 1.369 aY | 89.000 ± 12.100 aX | |
| 20 | 1.55 ± 0.652 bX | 7.4 ± 0.868 aX | 6.21 ± 0.609 aX | 12.240 ± 1.280 aY | 87.000 ± 10.787 aX |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, L.; Xu, S.; Shang, H.; Guo, J.; Yan, X.; Liu, C.; Li, G.; Luo, K. High-Voltage Electrostatic Fields Adversely Affect the Performance of Diamondback Moths over Five Consecutive Generations. Agronomy 2023, 13, 1008. https://doi.org/10.3390/agronomy13041008
Jia L, Xu S, Shang H, Guo J, Yan X, Liu C, Li G, Luo K. High-Voltage Electrostatic Fields Adversely Affect the Performance of Diamondback Moths over Five Consecutive Generations. Agronomy. 2023; 13(4):1008. https://doi.org/10.3390/agronomy13041008
Chicago/Turabian StyleJia, Li, Shicai Xu, Huanzhang Shang, Jiao Guo, Xia Yan, Changhai Liu, Guangwei Li, and Kun Luo. 2023. "High-Voltage Electrostatic Fields Adversely Affect the Performance of Diamondback Moths over Five Consecutive Generations" Agronomy 13, no. 4: 1008. https://doi.org/10.3390/agronomy13041008
APA StyleJia, L., Xu, S., Shang, H., Guo, J., Yan, X., Liu, C., Li, G., & Luo, K. (2023). High-Voltage Electrostatic Fields Adversely Affect the Performance of Diamondback Moths over Five Consecutive Generations. Agronomy, 13(4), 1008. https://doi.org/10.3390/agronomy13041008