
Study on the Regulation Mechanism of 1-MCP Combined with SO2 Treatment on Postharvest Senescence of Bamboo Shoots (Chimonobambusa quadrangularis) in Karst Mountain Area

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Respiratory Rate Measurement
2.3. Weight Loss Rate Measurement
2.4. Color Feature
2.5. Firmness
2.6. Lignin Content
2.7. MDA Content
2.8. Soluble Protein Content and Free Amino Acid Content
2.9. Determination of Ascorbic Acid Content and Total Flavone Content
2.10. Total Phenol Content
2.11. Enzyme Activity Measurement
2.12. RT-PCR
2.13. Statistical Analysis
3. Results
3.1. Changes of Respiration Rate and Weight Loss Rate

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Gene | Primer Sequences | Size (bp) | Annealing (°C) |
|---|---|---|---|---|
| Phyllostachys edulis | Actin | F:TGCCCTTGATTATGAGCAGG R:AACCTTTCTGCTCCGATGGT | 108 | 60 |
| Phyllostachys edulis | SOD | F:CTTTCCACTCGCTCCTCCTC R:TGATACGGGCGTTCACTGTT | 107 | 60 |
| Phyllostachys edulis | PPO | F:GATGATTGCCAGTGCCAAGA R:TCGGTGAAGTCGGTGTTGCT | 270 | 60 |
| Phyllostachys edulis | APX | F:CACCAACCGATGAGAAGAA R:GAGTAATTGGCAGCAACGA | 103 | 60 |
| Phyllostachys edulis | PAL | F:GAACAGCACAACCAAGATG R:TCTTTCTAGCCACCGTCGTC | 190 | 60 |
| Phyllostachys edulis | POD | F:CTTCGTCTTTCTCCTCGCATT R:TCTCAAGGTTTGGGCAGATG | 95 | 60 |
3.2. Changes of Free Amino Acid and Soluble Protein Contents
3.3. Changes of the Total Color Difference (ΔE) and Browning Index (BI)
3.4. Changes of the Total Phenolic Content and Polyphenol Oxidase (PPO) Activity
3.5. Changes of the Ascorbic Acid (ASA) and Total Flavonoids Content
3.6. Changes of the ROS Metabolism
3.7. Changes of Lignification Related Indexes
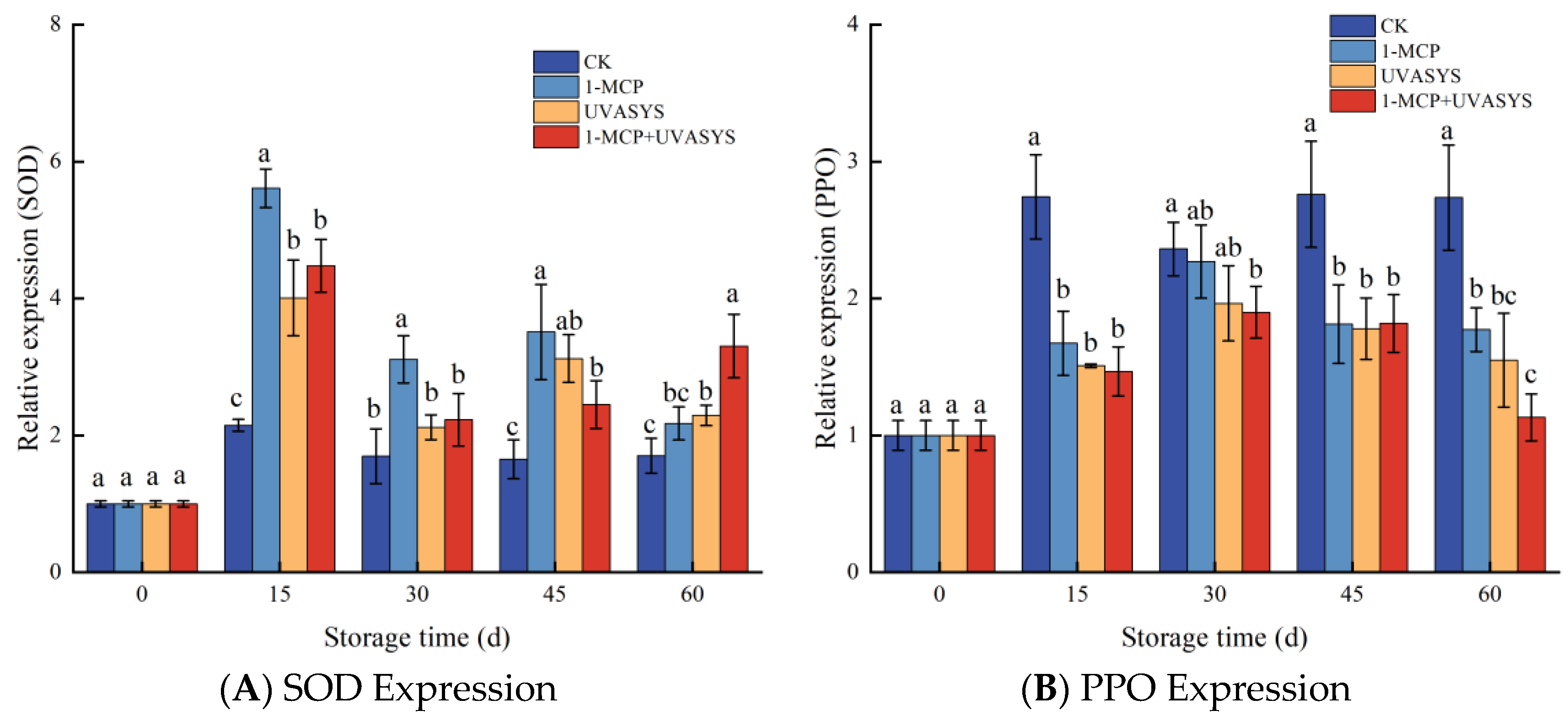
3.8. 1-MCP and UVASYS Affect the Relative Expression of Enzyme Genes
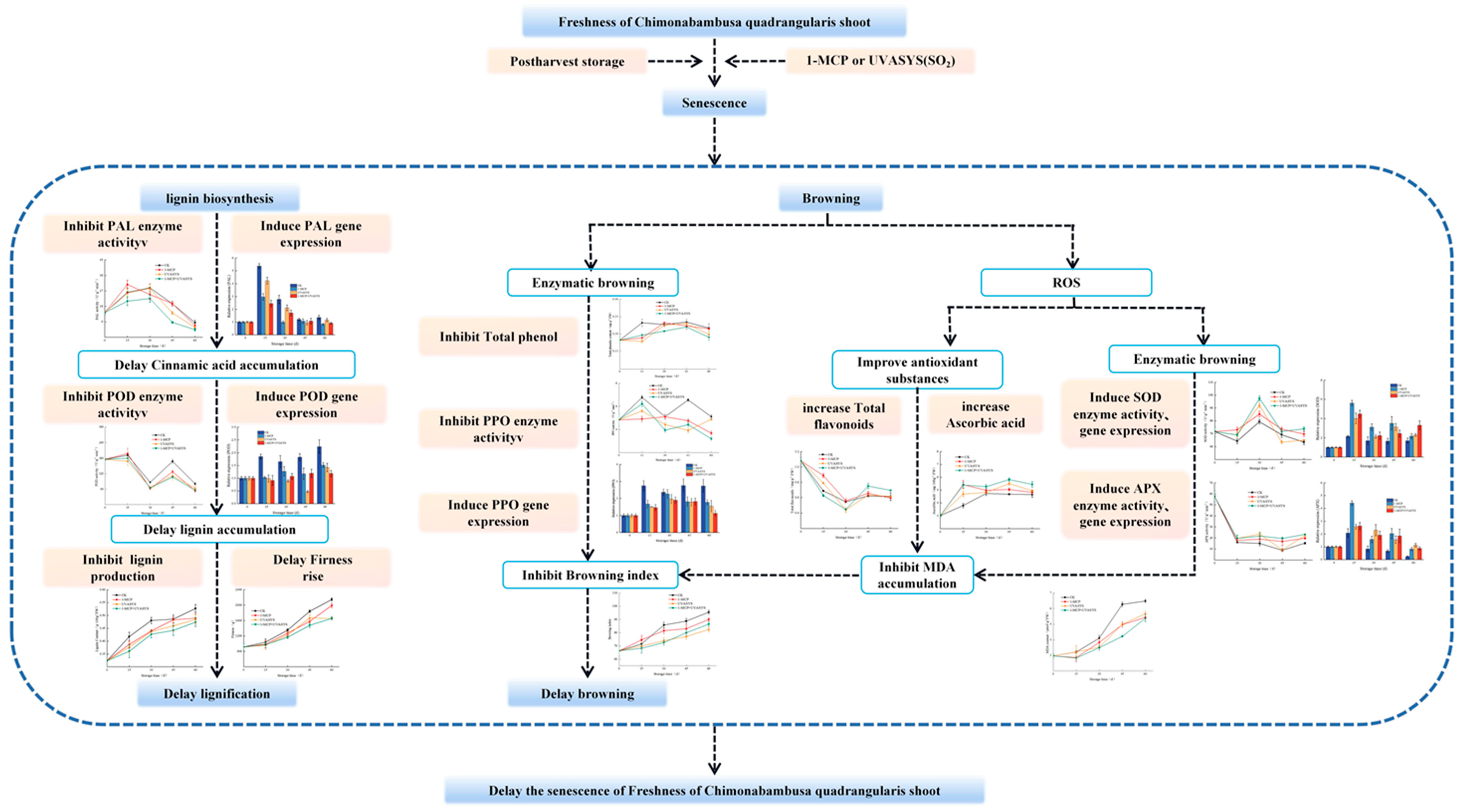
4. Discussion
4.1. Physiological and Nutritional Quality Changes
4.2. Appearance Color and Enzymatic Browning
4.3. Active Oxygen Metabolism
4.4. Lignification
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Christina, L. Climate change. An unsung carbon sink. Science 2011, 334, 886–887. [Google Scholar]
- He, X.Y.; Wang, K.L.; Zhang, W.; Chen, Z.H.; Zhu, Y.G.; Chen, H.S. Positive correlation between soil bacterial metabolic and plant species diversity and bacterial and fungal diversity in a vegetation succession on Karst. Plant Soil 2008, 307, 123–134. [Google Scholar] [CrossRef]
- Wang, S.J.; Liu, Q.M.; Zhang, D.F. Karst rocky desertification in southwestern China: Geomorphology, landuse, impact and rehabilitation. Land Degrad. Dev. 2004, 15, 115–121. [Google Scholar] [CrossRef]
- Bai, X.Y.; Wang, S.J.; Xiong, K.N. Assessing spatial-temporal evolution processes of karst rocky desertification land: Indications for restoration strategies. Land Degrad. Amp. Dev. 2013, 24, 47–56. [Google Scholar] [CrossRef]
- Peng, L.; Xu, X.J.; Liao, X.F. Ampelocalamus luodianensis (Poaceae), a plant endemic to karst, adapts to resource heterogeneity in differing microhabitats by adjusting its biomass allocation. Glob. Ecol. Conserv. 2023, 41, e02374. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, J.; Wang, D. A systematic review on the composition, storage, processing of bamboo shoots, focusing the nutritional and functional benefits. J. Funct. Foods 2020, 71, 104015. [Google Scholar] [CrossRef]
- Persin, Z.K.; Stana-Kleinschek, T.J.; Foster, J. Challenges and opportunities in polysaccharides research and technology: The EPNOEviews for the next decade in the areas of materials, food and health care. Carbohydr. Polym. 2011, 84, 22–32. [Google Scholar] [CrossRef]
- Zhang, J.; Li, S.; Maratab, A.L.I. Effects of UV-B treatment on controlling lignification and quality of bamboo (Phyllostachys prominens) shoots without sheaths during cold storage. J. Integr. Agric. 2020, 19, 1387–1395. [Google Scholar] [CrossRef]
- Li, D.; Limwachiranon, J.; Li, L. Hydrogen peroxide accelerated the lignification process of bamboo shoots by activating the phenylpropanoid pathway and programmed cell death in postharvest storage. Postharvest Biol. Technol. 2019, 153, 79–86. [Google Scholar] [CrossRef]
- Lata, D.; Homa, F.; Nayyer, M.A. Effect of postharvest hydrogen sulphide on lignification and biochemical markers of pointed gourd. Plant Biol. 2022, 24, 704–710. [Google Scholar] [CrossRef]
- Zhang, W.; Jiang, H.; Cao, J. UV-C treatment controls brown rot in postharvest nectarine by regulating ROS metabolism and anthocyanin synthesis. Postharvest Biol. Technol. 2021, 180, 111613. [Google Scholar] [CrossRef]
- Liu, J.; Huang, Q.; Kang, P. Lignin Accumulation in Three Pumelo Cultivars in Association with Sucrose and Energy Depletion. Biomolecules 2019, 9, 701. [Google Scholar] [CrossRef]
- Tobimatsu, Y.; Schuetz, M. Lignin polymerization: How do plants manage the chemistry so well? Curr. Opin. Biotechnol. 2019, 56, 75–81. [Google Scholar] [CrossRef]
- Rabiei, V.; Kakavand, F.; Zaare-Nahandi, F. Nitric oxide and γ-aminobutyric acid treatments delay senescence of cornelian cherry fruits during postharvest cold storage by enhancing antioxidant system activity. Sci. Hortic. 2019, 243, 268–273. [Google Scholar] [CrossRef]
- Cheng, S.; Yu, Y.; Guo, J. Effect of 1-methylcyclopropene and chitosan treatment on the storage quality of jujube fruit and its related enzyme activities. Sci. Hortic. 2020, 265, 109281. [Google Scholar] [CrossRef]
- Dias, C.; Ribeiro, T.; Rodrigues, A.C. Improving the ripening process after 1-MCP application: Implications and strategies. Trends Food Sci. Technol. 2021, 113, 382–396. [Google Scholar] [CrossRef]
- Luo, Z.; Xu, X.; Cai, Z. Effects of ethylene and 1-methylcyclopropene (1-MCP) on lignification of postharvest bamboo shoot. Food Chem. 2007, 105, 521–527. [Google Scholar] [CrossRef]
- Blankenship, S.M.; Dole, J.M. 1-Methylcyclopropene: A review. Postharvest Biol. Technol. 2003, 28, 1–25. [Google Scholar] [CrossRef]
- Wu, X.; An, X.; Yu, M. 1-Methylcyclopropene treatment on phenolics and the antioxidant system in postharvest peach combined with the liquid chromatography/mass spectrometry technique. J. Agric. Food Chem. 2018, 66, 6364–6372. [Google Scholar] [CrossRef]
- Liu, H.; Cao, J.; Jiang, W. Changes in phenolics and antioxidant property of peach fruit during ripening and responses to 1-methylcyclopropene. Postharvest Biol. Technol. 2015, 108, 111–118. [Google Scholar] [CrossRef]
- Langer, S.E.; Marina, M.; Francese, P. New insights into the cell wall preservation by 1-methylcyclopropene treatment in harvest-ripe strawberry fruit. Sci. Hortic. 2022, 299, 111032. [Google Scholar] [CrossRef]
- Dhiman, A.; Suhag, R.; Thakur, D. Current status of Loquat (Eriobotrya japonica Lindl.): Bioactive functions, preservation approaches, and processed products. Food Rev. Int. 2022, 38 (Suppl. 1), 286–316. [Google Scholar] [CrossRef]
- Dukare, A.S.; Paul, S.; Nambi, V.E. Exploitation of microbial antagonists for the control of postharvest diseases of fruits: A review. Crit. Rev. Food Sci. Nutr. 2019, 59, 1498–1513. [Google Scholar] [CrossRef]
- Joradol, A.; Uthaibutra, J.; Lithanatudom, P. Induced expression of NOX and SOD by gaseous sulfur dioxide and chlorine dioxide enhances antioxidant capacity and maintains fruit quality of ‘Daw’ longan fruit during storage through H2O2 signaling. Postharvest Biol. Technol. 2019, 156, 110938. [Google Scholar] [CrossRef]
- Yuan, Y.; Wei, J.; Xing, S. Sulfur dioxide (SO2) accumulation in postharvest grape: The role of pedicels of four different varieties. Postharvest Biol. Technol. 2022, 190, 111953. [Google Scholar] [CrossRef]
- Domingues, A.; Roberto, S.; Ahmed, S. Postharvest Techniques to Prevent the Incidence of Botrytis Mold of ‘BRS Vitoria’ Seedless Grape under Cold Storage. Horticulturae 2018, 4, 17. [Google Scholar] [CrossRef]
- Sabir, F.K.; Unal, S.; Sabir, A. Postharvest Aloe vera Gel Coatings Delay the Physiological Senescence of ‘Alphonse Lavallée’ and ‘Red Globe’ Grapes during Cold Storage as an Alternative to SO2. Erwerbs-Obstbau 2022, 1, 1–10. [Google Scholar]
- Pires, J.; Sousa, S.; Pereira, A. Management of air quality monitoring using principal component and cluster analysis—Part I: SO2 and PM10. Atmos. Environ. 2008, 42, 1249–1260. [Google Scholar] [CrossRef]
- Ahmed, S.; Roberto, S.; Domingues, A. Effects of Different Sulfur Dioxide Pads on Botrytis Mold in ‘Italia’ Table Grapes under Cold Storage. Horticulturae 2018, 4, 29. [Google Scholar] [CrossRef]
- Zhang, D.; Xu, X.; Zhang, Z. 6-Benzylaminopurine improves the quality of harvested litchi fruit. Postharvest Biol. Technol. 2018, 143, 137–142. [Google Scholar] [CrossRef]
- Xu, D.; Zuo, J.; Fang, Y. Effect of folic acid on the postharvest physiology of broccoli during storage. Food Chem. 2021, 339, 127981. [Google Scholar] [CrossRef]
- Gheysarbigi, S.; Mirdehghan, S.H.; Ghasemnezhad, M. The inhibitory effect of nitric oxide on enzymatic browning reactions of in-package fresh pistachios (Pistacia vera L.). Postharvest Biol. Technol. 2020, 159, 110998. [Google Scholar] [CrossRef]
- Li, C.; Suo, J.; Xuan, L. Bamboo shoot-lignification delay by melatonin during low temperature storage. Postharvest Biol. Technol. 2019, 156, 110933. [Google Scholar] [CrossRef]
- Fukushima, R.S.; Kerley, M.S.; Ramos, M.H. The acetyl bromide lignin method accurately quantitates lignin in forage. Anim. Feed Sci. Technol. 2021, 276, 114883. [Google Scholar] [CrossRef]
- Shi, J.; Zuo, J.; Zhou, F.; Gao, L. Low-temperature conditioning enhances chilling tolerance and reduces damage in cold-stored eggplant (Solanum melongena L.) fruit. Postharvest Biol. Technol. 2018, 141, 33–38. [Google Scholar] [CrossRef]
- Karimi, F.; Hamidian, Y.; Behrouzifar, F. An applicable method for extraction of whole seeds protein and its determination through Bradford’s method. Food Chem. Toxicol. 2022, 164, 113053. [Google Scholar] [CrossRef]
- Xu, L.; Fang, X.; Wu, W. Effects of high-temperature pre-drying on the quality of air-dried shiitake mushrooms (Lentinula edodes). Food Chem. 2019, 285, 406–413. [Google Scholar] [CrossRef]
- Shu, P.; Min, D.; Ai, W. L-Arginine treatment attenuates postharvest decay and maintains quality of strawberry fruit by promoting nitric oxide synthase pathway. Postharvest Biol. Technol. 2020, 168, 111253. [Google Scholar] [CrossRef]
- Du, S.; Zhang, J.; Chen, S. The combined effect of 1-methylcyclopropene and citral suppressed postharvest grey mould of tomato fruit by inhibiting the growth of Botrytis cinerea. J. Phytopathol. 2019, 167, 123–134. [Google Scholar]
- Bal, E. Effect of melatonin treatments on biochemical quality and postharvest life of nectarines. J. Food Meas. Charact. 2021, 15, 288–295. [Google Scholar] [CrossRef]
- Yang, X.; Zhao, Y.; Gu, Q. Effects of Naringin on Postharvest Storage Quality of Bean Sprouts. Foods 2022, 11, 2294. [Google Scholar] [CrossRef]
- Petriccione, M.; Pagano, L.; Forniti, R. Postharvest treatment with chitosan affects the antioxidant metabolism and quality of wine grape during partial dehydration. Postharvest Biol. Technol. 2018, 137, 38–45. [Google Scholar] [CrossRef]
- Xia, L.; Lu, Y.; Qian, Y. Cod peptides inhibit browning in fresh-cut potato slices, A potential anti-browning agent of random peptides for regulating food properties. Postharvest Biol. Technol. 2018, 146, 36–42. [Google Scholar]
- Kahramanoğlu, İ.; Chen, C.; Chen, Y. Improving storability of “nanfeng” mandarins by treating with postharvest hot water dipping. J. Food Qual. 2020, 2020, 8524952. [Google Scholar] [CrossRef]
- Qi, X.; Zhao, J.; Jia, Z. Potential metabolic pathways and related processes involved in pericarp browning for postharvest pomegranate fruits. Horticulturae 2022, 8, 924. [Google Scholar] [CrossRef]
- Wu, L.; Ma, N.; Jia, Y.C. An ethylene-induced regulatory module delays flower senescence by regulating cytokinin content. Plant Physiol. 2017, 173, 853–862. [Google Scholar] [CrossRef]
- Li, C.; Yu, W.; Liao, W. Role of Nitric Oxide in Postharvest Senescence of Fruits. Int. J. Mol. Sci. 2022, 23, 10046. [Google Scholar] [CrossRef]
- Wu, Z.; Tu, M.; Yang, X. Effect of cutting on the reactive oxygen species accumulation and energy change in postharvest melon fruit during storage. Sci. Hortic. 2019, 257, 108752. [Google Scholar] [CrossRef]
- Xuan, J.J.; Lin, H.; Shi, J. Effects of a novel chitosan formulation treatment on quality attributes and storage behavior of harvested litchi fruit. Food Chem. 2018, 252, 134–141. [Google Scholar]
- Zhang, W.; Zhao, H.; Zhang, J. Different molecular weights chitosan coatings delay the senescence of postharvest nectarine fruit in relation to changes of redox state and respiratory pathway metabolism. Food Chem. 2019, 289, 160–168. [Google Scholar] [CrossRef]
- Zheng, J.; Li, S.; Xu, Y. Effect of oxalic acid on edible quality of bamboo shoots (Phyllostachys prominens) without sheaths during cold storage. Lebensm. Wiss. Und Technol. 2019, 109, 194–200. [Google Scholar] [CrossRef]
- Ali, M.; Raza, M.A.; Li, S. 1-MCP regulates ethanol fermentation and GABA shunt pathway involved in kiwifruit quality during postharvest storage. Hortic. Plant J. 2021, 7, 23–30. [Google Scholar] [CrossRef]
- Sortino, G.; Farina, V.; Gallotta, A. Effect of low SO2 postharvest treatment on quality parameters of ‘Italia’ table grape during prolonged cold storage. Acta Hortic. 2018, 1194, 695–700. [Google Scholar] [CrossRef]
- Li, Y.; Ishikawa, Y.; Satake, T. Effect of active modified atmosphere packaging with different initial gas compositions on nutritional compounds of shiitake mushrooms (Lentinus edodes). Postharvest Biol. Technol. 2014, 92, 107–113. [Google Scholar] [CrossRef]
- Li, T.; Shi, D.; Wu, Q. Sodium para-aminosalicylate delays pericarp browning of litchi fruit by inhibiting ROS-mediated senescence during postharvest storage. Food Chem. 2019, 278, 552–559. [Google Scholar] [CrossRef]
- Zhang, J.; Murtaza, A.; Zhu, L. High pressure CO2 treatment alleviates lignification and browning of fresh-cut water-bamboo shoots (Zizania latifolia). Postharvest Biol. Technol. 2021, 182, 111690. [Google Scholar] [CrossRef]
- Ali, S.; Khan, A.S.; Malik, A.U. Modified atmosphere packaging delays enzymatic browning and maintains quality of harvested litchi fruit during low temperature storage. Sci. Hortic. 2019, 254, 14–20. [Google Scholar] [CrossRef]
- Luo, S.; Hu, H.; Wang, Y. The role of melatonin in alleviating the postharvest browning of lotus seeds through energy metabolism and membrane lipid metabolism. Postharvest Biol. Technol. 2020, 167, 111243. [Google Scholar] [CrossRef]
- Chen, J.; Xu, Y.; Yi, Y. Regulations and mechanisms of 1-methylcyclopropene treatment on browning and quality of fresh-cut lotus (Nelumbo nucifera Gaertn.) root slices. Postharvest Biol. Technol. 2022, 185, 111782. [Google Scholar] [CrossRef]
- Aguiar, A.C.; Higuchi, M.T.; Ribeiro, L.T.M. Bio-based and SO2-generating plastic liners to extend the shelf life of ‘Benitaka’ table grapes. Postharvest Biol. Technol. 2023, 197, 112217. [Google Scholar] [CrossRef]
- Toivonen, P.M.A.; Brummell, D.A. Biochemical bases of appearance and texture changes in fresh-cut fruit and vegetables. Postharvest Biol. Technol. 2008, 48, 1–14. [Google Scholar] [CrossRef]
- Dong, T.; Cao, Y.; Li, G. A novel aspartic protease inhibitor inhibits the enzymatic browning of potatoes. Postharvest Biol. Technol. 2021, 172, 111353. [Google Scholar] [CrossRef]
- Li, X.; Li, C.; Cheng, Y. Postharvest application of acibenzolar-S-methyl delays the senescence of pear fruit by regulating reactive oxygen species and fatty acid metabolism. J. Agric. Food Chem. 2020, 68, 4991–4999. [Google Scholar] [CrossRef]
- Chen, C.; Chen, J.; Nie, Z. Chitosan coating alleviates postharvest juice sac granulation by mitigating ROS accumulation in harvested pummelo (Citrus grandis L. Osbeck) during room temperature storage. Postharvest Biol. Technol. 2020, 169, 111309. [Google Scholar]
- Dunnill, C.; Patton, T.; Brennan, J. Reactive oxygen species (ROS) and wound healing: The functional role of ROS and emerging ROS-modulating technologies for augmentation of the healing process. Int. Wound J. 2017, 14, 89–96. [Google Scholar] [CrossRef]
- Chen, C.; Nie, Z.; Wan, C. Preservation of Xinyu Tangerines with an Edible Coating Using Ficus hirta Vahl. Fruits Extract-Incorporated Chitosan. Biomolecules 2019, 9, 46. [Google Scholar] [CrossRef]
- Wang, H.; Chen, G.; Shi, L. Influences of 1-methylcyclopropene-containing papers on the metabolisms of membrane lipids in Anxi persimmons during storage. Food Qual. Saf. 2020, 4, 143–150. [Google Scholar] [CrossRef]
- Juan, P.Z. Hairline, a postharvest cracking disorder in table grapes induced by sulfur dioxide. Postharvest Biol. Technol. 2008, 47, 90–97. [Google Scholar]
- Silva, W.B.; Silva, G.M.C.; Santana, D.B. Chitosan delays ripening and ROS production in guava (Psidium guajava L.) fruit. Food Chem. 2018, 242, 232–238. [Google Scholar] [CrossRef]
- Lwin, W.W.; Srilaong, V.; Boonyaritthongchai, P. Electrostatic atomised water particles reduces postharvest lignification and maintain asparagus quality. Sci. Hortic. 2020, 271, 109487. [Google Scholar] [CrossRef]
- Liu, W.; Zhang, J.; Jiao, C. Transcriptome analysis provides insights into the regulation of metabolic processes during postharvest cold storage of loquat (Eriobotrya japonica) fruit. Hortic. Res. 2019, 6, 11. [Google Scholar] [CrossRef]
- García-Rojas, M.; Meneses, M.; Oviedo, K.; Carrasco, C.; Defilippi, B.; González-Agüero, M.; León, G.; Hinrichsen, P. Exogenous gibberellic acid application induces the overexpression of key genes for pedicel lignification and an increase in berry drop in table grape. Plant Physiol. Biochem. 2018, 126, 32–38. [Google Scholar] [CrossRef]
- Boudet, A.M. Lignins and lignification: Selected issues. Plant Physiol. Biochem. 2000, 38, 81–96. [Google Scholar] [CrossRef]
- Yang, B.; Fang, X.; Han, Y. Analysis of lignin metabolism in water bamboo shoots during storage. Postharvest Biol. Technol. 2022, 192, 111989. [Google Scholar] [CrossRef]
- Xie, G.; Feng, Y.; Chen, Y. Effects of 1-Methylcyclopropene (1-MCP) and Ethylene on Postharvest Lignification of Common Beans (Phaseolus vulgaris L.). ACS Omega 2020, 5, 8659–8666. [Google Scholar] [CrossRef]
- Rahimi, J.; Jhalegar, J.; Meti, S. Effect of ozone and sulphur dioxide as postharvest treatment to control Rhizopus rot and quality maintenance of table grape (Vitis vinifera L.). Int. J. Chem. Res. 2020, 8, 1816–1824. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, J.; Ji, N.; Wang, R.; Ma, C.; Lei, J.; Zhang, N.; Liu, R.; Deng, Y. Study on the Regulation Mechanism of 1-MCP Combined with SO2 Treatment on Postharvest Senescence of Bamboo Shoots (Chimonobambusa quadrangularis) in Karst Mountain Area. Agronomy 2023, 13, 1122. https://doi.org/10.3390/agronomy13041122
Xu J, Ji N, Wang R, Ma C, Lei J, Zhang N, Liu R, Deng Y. Study on the Regulation Mechanism of 1-MCP Combined with SO2 Treatment on Postharvest Senescence of Bamboo Shoots (Chimonobambusa quadrangularis) in Karst Mountain Area. Agronomy. 2023; 13(4):1122. https://doi.org/10.3390/agronomy13041122
Chicago/Turabian StyleXu, Jinyang, Ning Ji, Rui Wang, Chao Ma, Jiqing Lei, Ni Zhang, Renchan Liu, and Yunbing Deng. 2023. "Study on the Regulation Mechanism of 1-MCP Combined with SO2 Treatment on Postharvest Senescence of Bamboo Shoots (Chimonobambusa quadrangularis) in Karst Mountain Area" Agronomy 13, no. 4: 1122. https://doi.org/10.3390/agronomy13041122
APA StyleXu, J., Ji, N., Wang, R., Ma, C., Lei, J., Zhang, N., Liu, R., & Deng, Y. (2023). Study on the Regulation Mechanism of 1-MCP Combined with SO2 Treatment on Postharvest Senescence of Bamboo Shoots (Chimonobambusa quadrangularis) in Karst Mountain Area. Agronomy, 13(4), 1122. https://doi.org/10.3390/agronomy13041122