
Competitiveness of Bensulfuron-Methyl-Susceptible and -Resistant Biotypes of Ammannia auriculata Willd. in Direct-Seeded Rice
 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Greenhouse Study
2.2. Field Study
2.3. Statistical Analysis
3. Results
3.1. Greenhouse Experiment
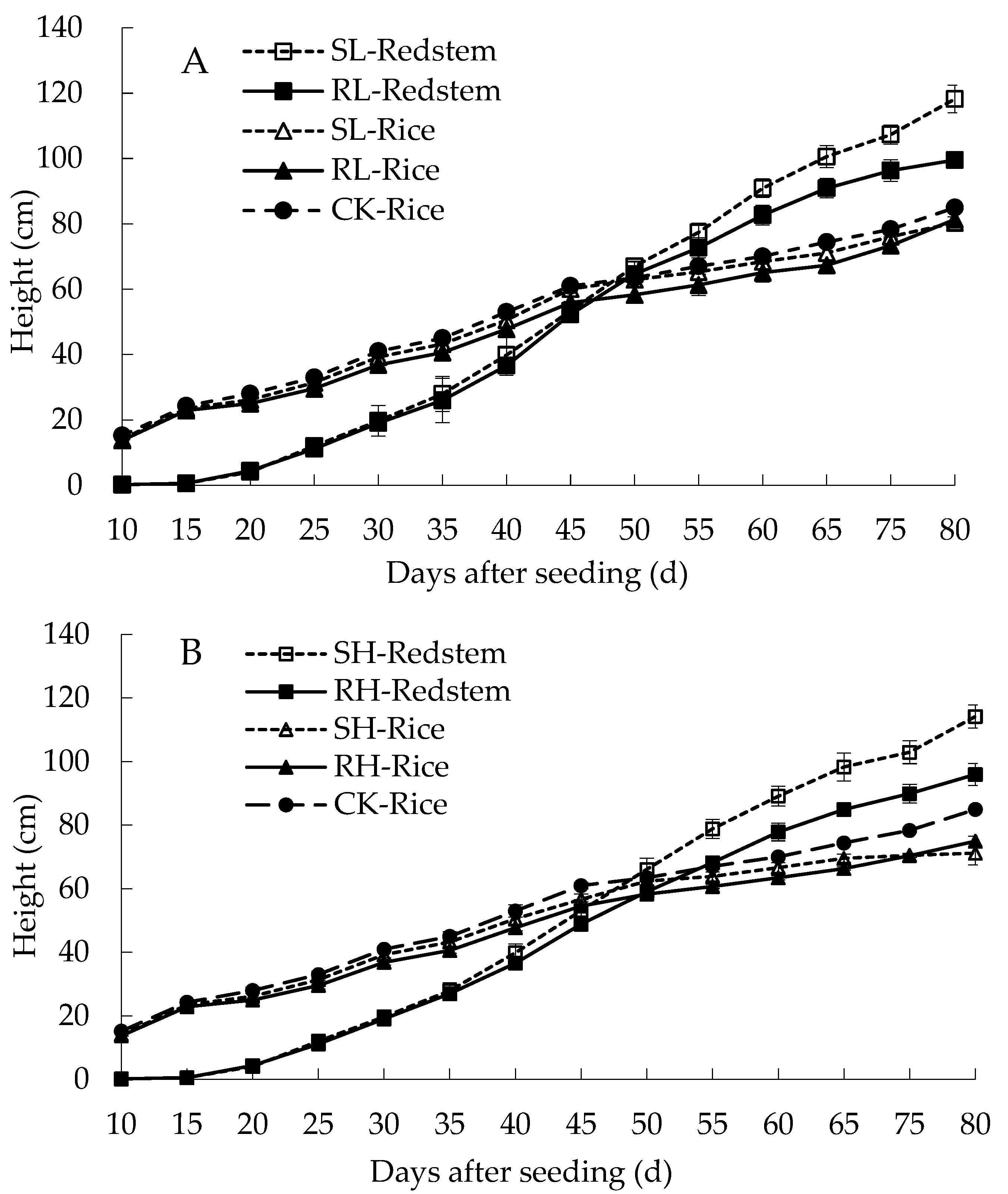
3.1.1. Plant Height of Eared Redstem and Rice
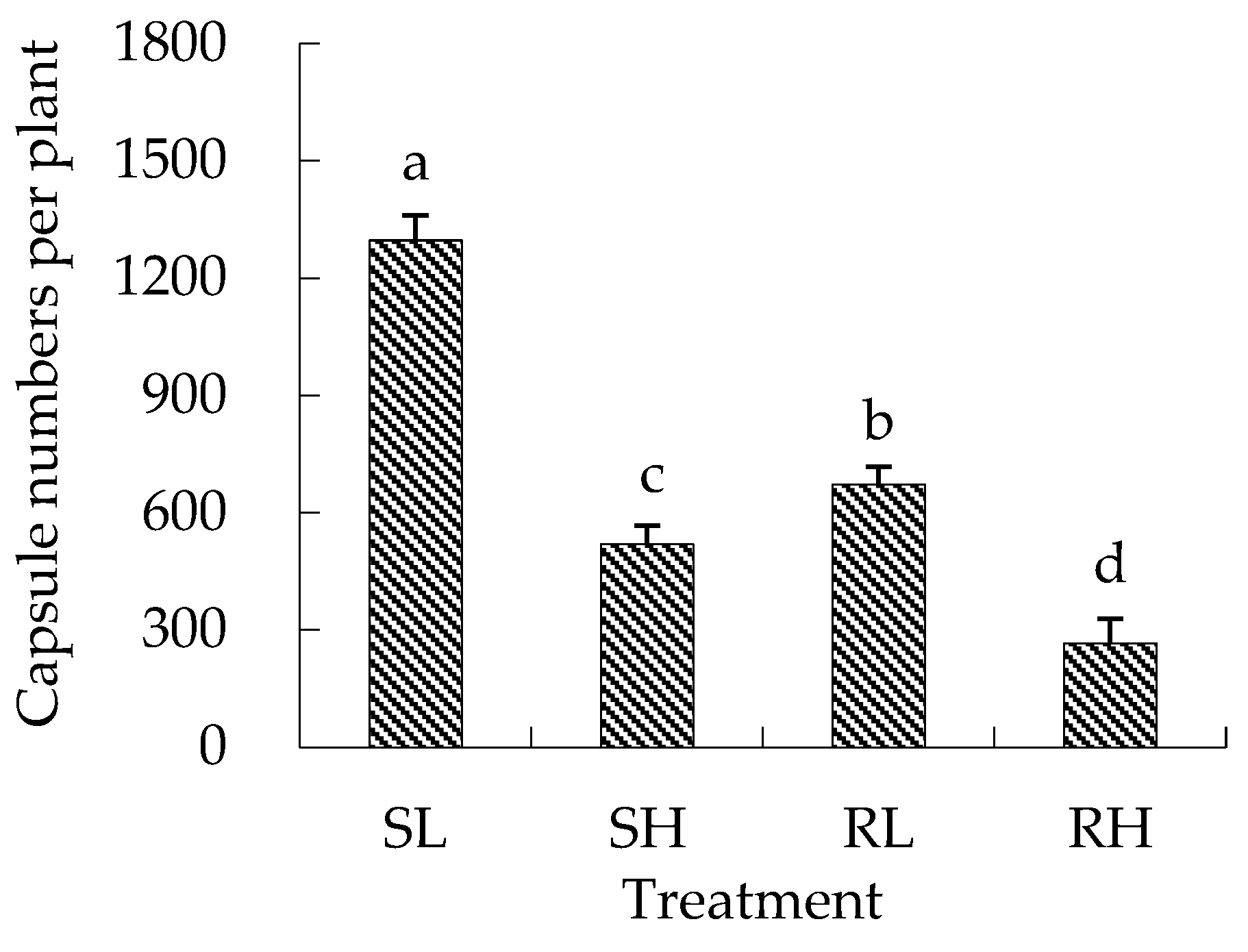
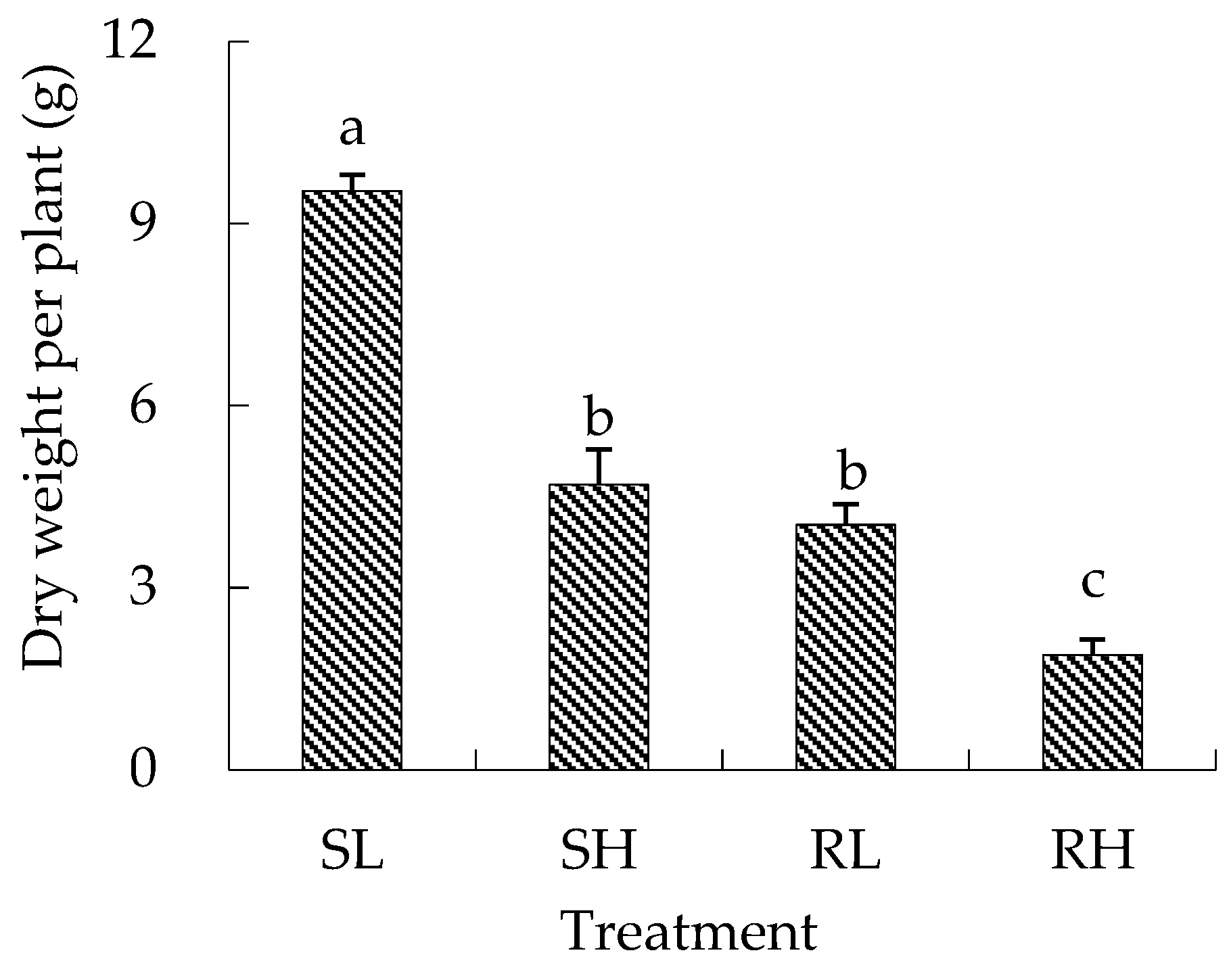
3.1.2. Branches, Capsules, and Dry Weight of Eared Redstem
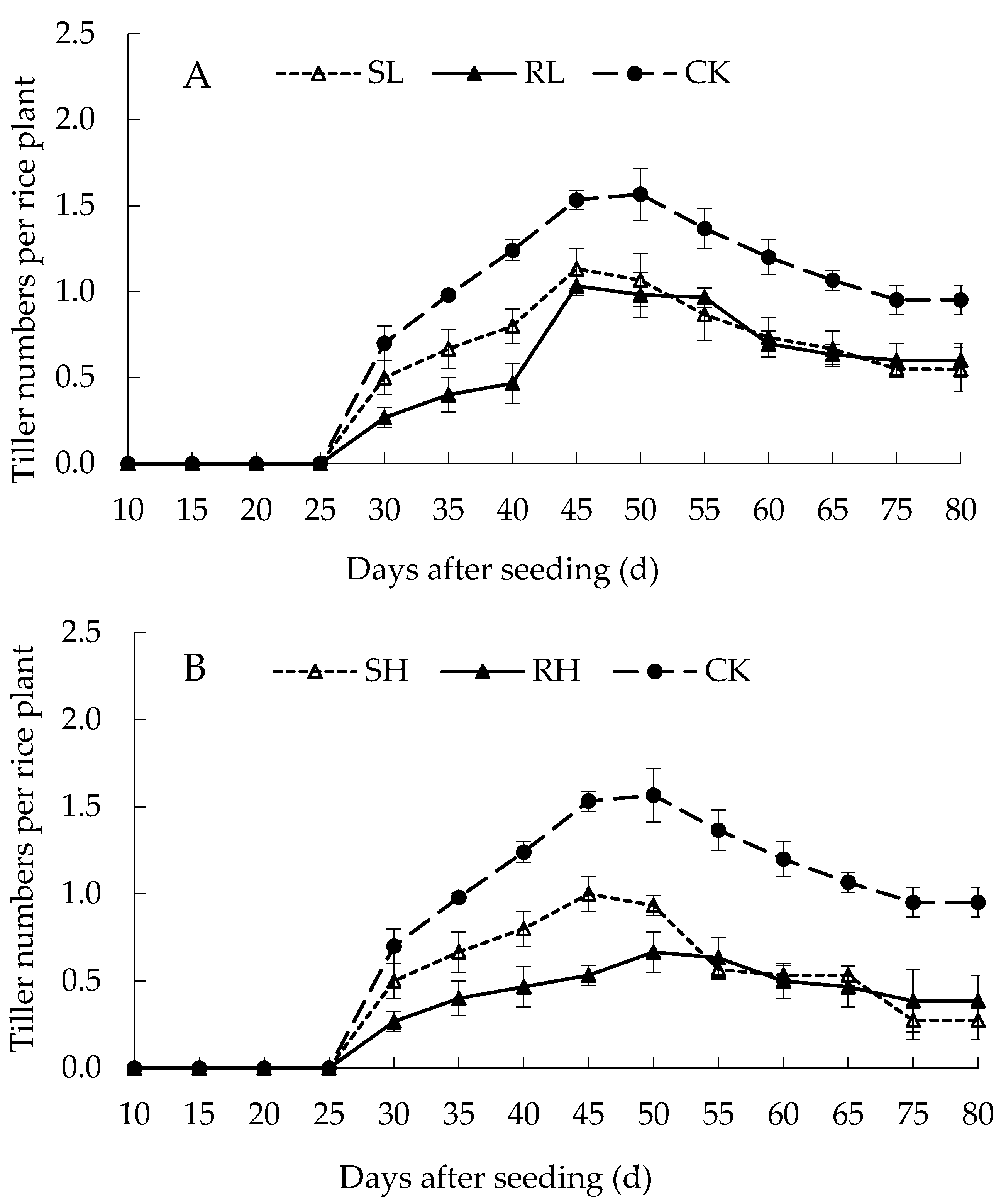
3.1.3. Tillering and Yield of Rice
3.2. Field Experiment
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- NBSC (National Bureau of Statistics of China). Available online: http://data.stats.gov.cn/easyquery.htm?cn=C01 (accessed on 8 November 2021).
- Chen, L.; Yi, Y.; Wang, W.; Zeng, Y.; Zeng, Y. Innovative furrow ridging fertilization under a mechanical direct seeding system improves the grain yield and lodging resistance of early indica rice in South China. Field Crops Res. 2021, 270, 108184. [Google Scholar]
- Sha, W.B.; Chen, F.B.; Mishra, A.K. Adoption of direct seeded rice, land use and enterprise income: Evidence from Chinese rice producers. Land Use Policy 2019, 83, 564–570. [Google Scholar]
- Bajwa, A.A.; Ullah, A.; Farooq, M.; Chauhan, B.S.; Adkins, S. Competition dynamics of Parthenium hysterophorusin direct-seeded aerobic rice fields. Exp. Agr. 2020, 56, 196–203. [Google Scholar]
- Deng, W.; Yang, M.T.; Duan, Z.W.; Peng, C.; Xia, Z.M.; Yuan, S.Z. Molecular basis of resistance to bensulfuron-methyl and cross-resistance patterns to ALS-inhibiting herbicides in Ludwigia prostrata. Weed Technol. 2021, 35, 656–661. [Google Scholar]
- Yamaguchi, T.; Matsumura, K.; Iwakami, S.; Sunohara, Y.; Matsumoto, H. Heterologous expression of CYP81A6 from rice (Oryza sativa) in Escherichia coli and structural analyses of bensulfuron-methyl metabolites. Weed Biol. Manag. 2021, 21, 164–171. [Google Scholar] [CrossRef]
- Heap, I. The International Survey of Herbicide Resistant Weeds. Available online: www.weedscience.org (accessed on 13 June 2019).
- Liu, R.; Zhu, J.W.; Zhou, G.J.; Xu, L.Y.; Wang, X.G.; Liu, Y.G.; Shi, Y.K. Efficacy comparison of five herbicides combinations to direct seeding paddy weeds. J. Zhejiang Agri. Sci. 2012, 318, 363–366. (In Chinese) [Google Scholar]
- Zhu, J.W.; Wang, J.; Ditommaso, A.; Zhang, C.X.; Zheng, G.P.; Liang, W.; Islam, F.; Yang, C.; Chen, X.X.; Zhou, W.J. Weed research status, challenges, and opportunities in China. Crop Prot. 2020, 134, 104449. [Google Scholar]
- Wei, S.; Li, P.; Ji, M.; Dong, Q.; Wang, H. Target-site resistance to bensulfuron-methyl in Sagittaria trifolia L. populations. Pestic. Biochem. Physiol. 2015, 124, 81–85. [Google Scholar]
- Gao, L.S.; Cui, H.L.; Luo, Y.P.; Li, X.J. Sensitivity research of Cyperus difformis to different herbicides. Hubei Agri. Sci. 2015, 54, 2123–2126. (In Chinese) [Google Scholar]
- Zhu, J.W.; Wang, J.; Wang, W.; Yang, S.W.; Lu, Y.L.; Cheng, J.J.; Zhou, W.J. Weeds of Rice and Their Management in China. In Weed Management in Rice in the Asian-Pacific Region; Asian-Pacific Weed Science Society: Hyderabad, India, 2017. [Google Scholar]
- Chaudhary, A.; Chhokar, R.S.; Dhanda, S.; Kaushik, P.; Punia, S.S. Herbicide resistance to metsulfuron-methyl in Rumex dentatus L. in north-west India and its management perspectives for sustainable wheat production. Sustainability 2021, 13, 6947. [Google Scholar]
- Gharde, Y.; Singh, P.K.; Dubey, R.P.; Gupta, P.K. Assessment of yield and economic losses in agriculture due to weeds in India. Crop Prot. 2018, 107, 12–18. [Google Scholar]
- Zimdahl, R.L. Weed-Crop Competition: A Review, 2nd ed.; Blackwell Publishers: Oxford, UK, 2008. [Google Scholar]
- Vila-Aiub, M.M.; Neve, P.; Powles, S.B. Fitness costs associated with evolved herbicide resistance alleles in plants. New Phytol. 2009, 184, 751–767. [Google Scholar] [PubMed]
- Asaduzzaman, M.; Koetz, E.; Wu, H.W.; Hopwood, M.; Shephard, A. Fate and adaptive plasticity of heterogeneous resistant population of Echinochloa colona in response to glyphosate. Sci. Rep. 2021, 11, 14858. [Google Scholar]
- Li, Y.H. Weeds of China; China Agriculture Press: Beijing, China, 1998. (In Chinese) [Google Scholar]
- GBIF (Global Biodiversity Information Facility). Available online: https://www.gbif.org/species/3188714 (accessed on 4 March 2023).
- Liu, B. Study on Resistance to ALS Inhibitors of AMMANNIA ARENARIA H·B·K and the Expression of ALS Gene. Master’s Thesis, Zhejiang University, Zhejiang, China, 2015. (In Chinese). [Google Scholar]
- Caton, B.P.; Foin, T.C.; Hill, J.E. Phenotypic plasticity of Ammannia spp. in competition with rice. Weed Res. 1997, 37, 33–38. [Google Scholar]
- Lu, B.L.; Zhang, J.X.; Wang, Y.X.; Gan, H.H.; Wang, W.P.; Huang, H.Y. A brief report on the control of Ammannia auriculata. Shanghai Agri. Sci. Tech. 2008, 4, 127–128. (In Chinese) [Google Scholar]
- Pappas-Fader, T.T.R.G.; Cook, J.F.; Butler, T.D.; Lana, P.J.; Carriere, M. Resistance monitoring program for aquatic weeds to sulfonylurea herbicides in California rice fields. Proc. Rice Tech. Work Group 1994, 25, 165. [Google Scholar]
- Wang, X.G.; Xu, Q.F.; Zhu, J.W.; Liu, R.; Wang, S.R.; Liu, Y.G.; Lu, Q.; Wang, G.R. Resistance comparison of Ammannia arenaria to bensulfuron-methyl in different paddy rice growing regions of Zhejiang Province. Chin. J. Pestic. Sci. 2013, 15, 52–58. (In Chinese) [Google Scholar]
- Liu, R. Research on Resistance of Ammannia auriculata H·B·K to Bensulfuron-Methyl and Its Biological Characteristics. Master’s Thesis, Zhejiang University, Zhejiang, China, 2012. (In Chinese). [Google Scholar]
- Li, T.; Shen, G.H.; Ping, L.F.; Lu, B.L.; Qian, Z.G.; Wen, G.Y.; Gan, H.H.; Wang, W.P. Occurrence and biological characteristics of Amaranthus auriculata. Plant Prot. 2011, 37, 172–175. (In Chinese) [Google Scholar]
- Gibson, K.D.; Fischer, A.J.; Foin, T.C. Shading and the growth and photosynthetic responses of Ammannia coccinnea. Weed Res. 2001, 41, 59–67. [Google Scholar]
- Tian, Z.H.; Shen, G.H.; Yuan, G.H.; Song, K.; Lu, J.Y.; Da, L.J. Effects of Echinochloa crusgalli and Cyperus difformis on yield and eco-economic thresholds of rice. J. Clean. Prod. 2020, 259, 120807. [Google Scholar]
- Bajwa, A.A.; Ullah, A.; Farooq, M.; Chauhan, B.S.; Adkins, S. Effect of different densities of parthenium weed (Parthenium hysterophorus L.) on the performance of direct-seeded rice under aerobic conditions. Arch. Agron. Soil Sci. 2019, 65, 796–808. [Google Scholar]
- Han, H.P.; Vila-Aiub, M.M.; Jalaludin, A.; Yu, Q.; Powles, S.B. A double EPSPS gene mutation endowing glyphosate resistance shows a remarkably high resistance cost. Plant Cell Environ. 2017, 40, 3031–3042. [Google Scholar] [CrossRef] [PubMed]
- Frenkel, E.; Matzrafi, M.; Rubin, B.; Peleg, Z. Effects of Environmental Conditions on the Fitness Penalty in Herbicide Resistant Brachypodium hybridum. Front. Plant Sci. 2017, 8, 94. [Google Scholar] [CrossRef]
- Hassanpour-Bourkheili, S.; Heravi, M.; Gherekhloo, J.; Alcántara-De La Cruz, R.; De Prado, R. Fitness Cost of Imazamox Resistance in Wild Poinsettia (Euphorbia heterophylla L.). Agronomy 2020, 10, 1859. [Google Scholar] [CrossRef]
- Keshtkar, E.; Abdolshahi, R.; Sasanfar, H.; Zand, E.; Beffa, R.; Dayan, F.E.; Kudsk, P. Assessing fitness costs from a herbicide-resistance management perspective: A review and insight. Weed Sci. 2019, 67, 137–148. [Google Scholar]
- Vila-Aiub, M.M. Fitness of herbicide-resistant weeds: Current knowledge and implications for management. Plants 2019, 8, 469. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Liang, W.; Yang, S.; Wang, H.; Ditommaso, A. Safety of oilseed rape straw mulch of different lengths to rice and its suppressive effects on weeds. Agronomy 2020, 10, 201. [Google Scholar] [CrossRef]
- Zhu, J.W.; Dong, Q.M.; Liu, B.; Lu, L.H.; Zhou, J.; Liu, C.; Zhang, L.Y.; Huang, Y.Z. Effect of soil moisture, submersion depth and soil thickness on seed germination of herbicide resistant Ammannia arenaria. Weed Sci. 2014, 32, 39–41. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Spike Number Per 0.063 m2 | Reduction in Spike Number (%) | Dry Weight of Rice (kg ha−1) | Yield Reduction (%) |
|---|---|---|---|---|
| CK | 18.7 ± 1.8 a | 5275.5 ± 403.2 a | ||
| SL | 11.0 ± 1.5 b | 41.2 | 1419.4 ± 157.7 c | 73.1 |
| SH | 1.8 ± 0.7 d | 90.4 | 88.4 ± 27.7 e | 98.3 |
| RL | 11.2 ± 1.6 b | 40.1 | 2596.9 ± 266.0 b | 50.8 |
| RH | 7.0 ± 0.9 c | 62.6 | 960.5 ± 91.8 d | 81.8 |
| Eared Redstem Density (Plants m−2) | Paddy Rice | |||
|---|---|---|---|---|
| Spike Number Per 0.25 m2 | Reduction in Spike Number (%) | Dry Weight of Rice (kg ha−1) | Yield Reduction (%) | |
| 0 (CK) | 101.7 ± 1.2 a | 8719.9 ± 889.3 a | ||
| 5 | 82.7 ± 3.5 b | 18.7 | 6786.5 ± 398.3 b | 22.2 |
| 10 | 80.0 ± 2.0 b | 21.3 | 6034.2 ± 299.1 c | 30.8 |
| 20 | 78.4 ± 3.1 bc | 22.9 | 5129.7 ± 241.1 d | 41.2 |
| 30 | 73.7 ± 3.6 cd | 27.5 | 4069.6 ± 228.6 e | 53.3 |
| 40 | 71.4 ± 4.3 d | 29.8 | 3385.5 ± 195.2 e | 61.2 |
| 50 | 55.9 ± 1.7 e | 45.0 | 2586.9 ± 180.0 f | 70.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, S.; Liu, J.; Liu, R.; Zhou, G.; Chen, C.; Zhou, W.; Ali, B.; Gui, W.; Zhu, J.; DiTommaso, A. Competitiveness of Bensulfuron-Methyl-Susceptible and -Resistant Biotypes of Ammannia auriculata Willd. in Direct-Seeded Rice. Agronomy 2023, 13, 1152. https://doi.org/10.3390/agronomy13041152
Yang S, Liu J, Liu R, Zhou G, Chen C, Zhou W, Ali B, Gui W, Zhu J, DiTommaso A. Competitiveness of Bensulfuron-Methyl-Susceptible and -Resistant Biotypes of Ammannia auriculata Willd. in Direct-Seeded Rice. Agronomy. 2023; 13(4):1152. https://doi.org/10.3390/agronomy13041152
Chicago/Turabian StyleYang, Siyu, Jie Liu, Rui Liu, Guojun Zhou, Chang Chen, Weijun Zhou, Basharat Ali, Wenjun Gui, Jinwen Zhu, and Antonio DiTommaso. 2023. "Competitiveness of Bensulfuron-Methyl-Susceptible and -Resistant Biotypes of Ammannia auriculata Willd. in Direct-Seeded Rice" Agronomy 13, no. 4: 1152. https://doi.org/10.3390/agronomy13041152
APA StyleYang, S., Liu, J., Liu, R., Zhou, G., Chen, C., Zhou, W., Ali, B., Gui, W., Zhu, J., & DiTommaso, A. (2023). Competitiveness of Bensulfuron-Methyl-Susceptible and -Resistant Biotypes of Ammannia auriculata Willd. in Direct-Seeded Rice. Agronomy, 13(4), 1152. https://doi.org/10.3390/agronomy13041152