


Effects of Pruning on Tea Tree Growth, Soil Enzyme Activity and Microbial Diversity

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site and Soil Sample Collection
2.2. Determination of the Growth Index of Tea Trees
2.3. Soil Sample Collection and Determination of Physicochemical Indexes
2.4. Determination of Soil Enzyme Activity
2.5. Determination of Functional Diversity of Microorganisms
2.6. Data Analysis
3. Results
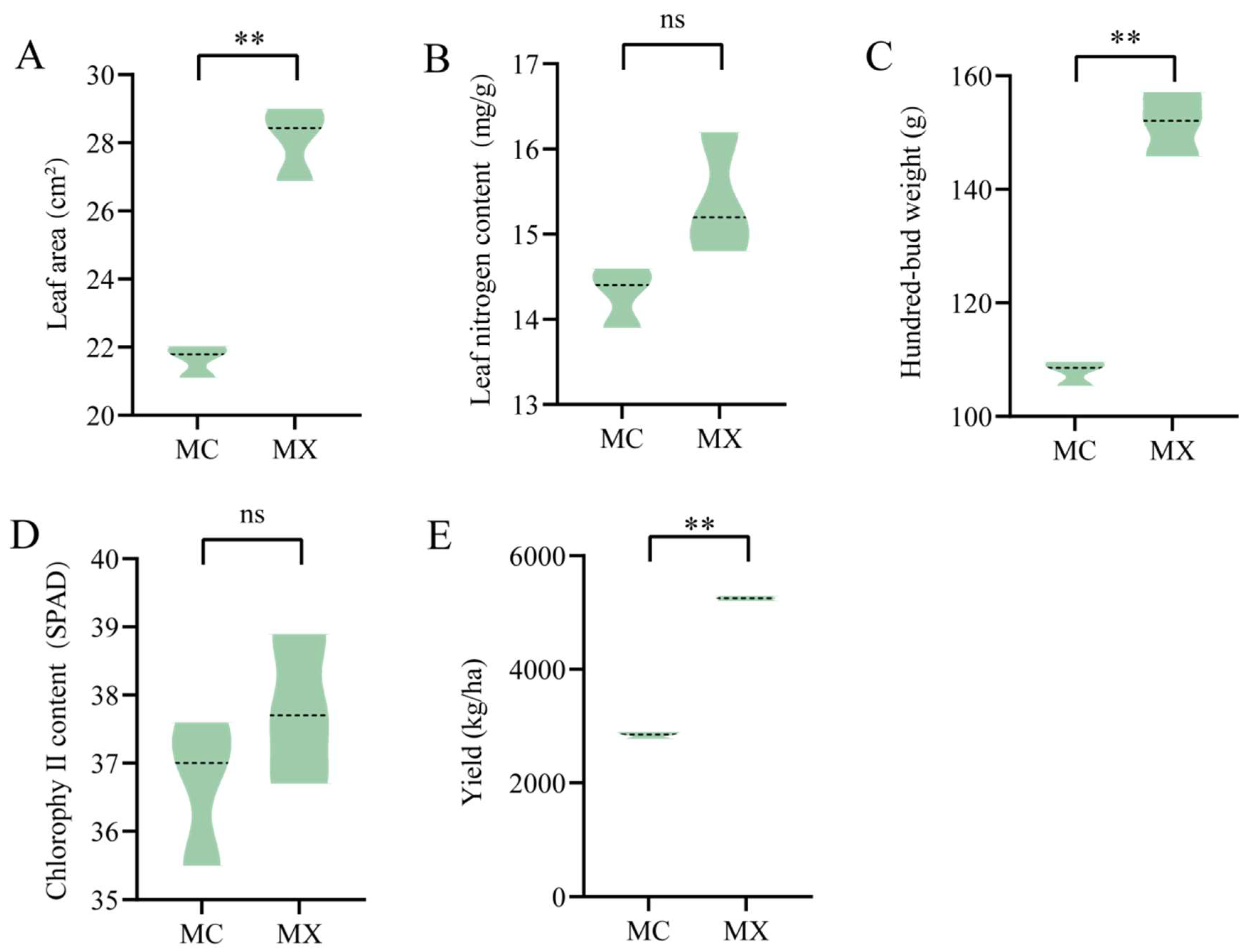
3.1. Analysis of Tea Tree Growth Indexes
3.2. Analysis of Soil Physicochemical Index
3.3. Analysis of Soil Enzyme Activity
3.4. Correlation Analysis
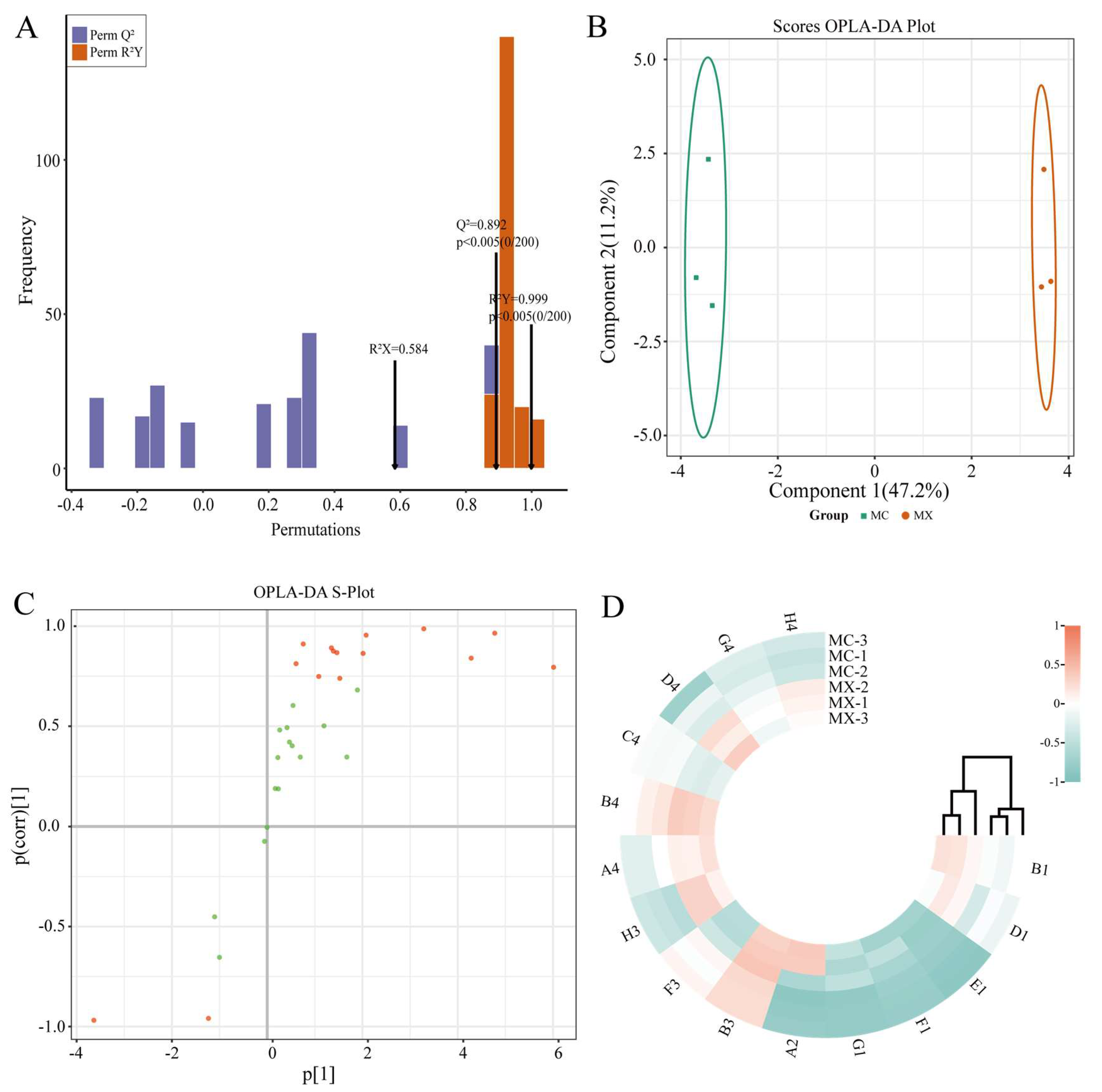
3.5. Analysis of Microbial Functional Diversity in Rhizosphere Soil Microorganisms of Tea Trees
3.6. Carbon Source Screening for Microbial Differences in Rhizosphere Soil of Tea Trees
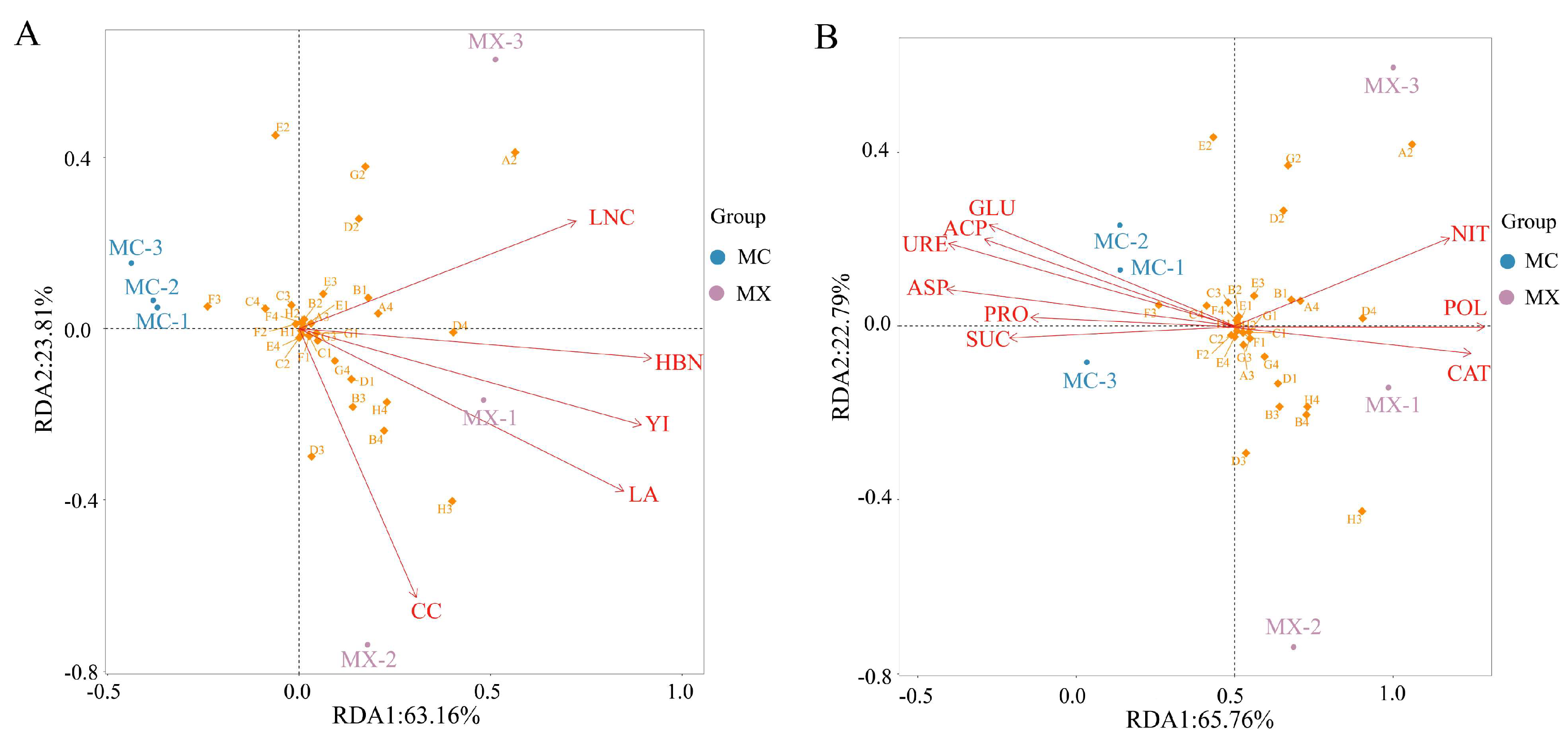
3.7. Redundancy Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wijeratn, M.A. Planting and cultivation of tea. In Global Tea Science: Current Status and Future Needs, 1st ed.; Sharma, Gunasekare, K., Eds.; Burleigh Dodds Science Publishing: London, UK, 2018; p. 32. [Google Scholar]
- Tian, R.Q.; Lv, R.Q.; Fang, L.; Shi, Y.Z. Study on the influence of different pruning methods on tree crown formation in tea plantation with high quality tea machine. Chin. Tea 2015, 37, 16–19. [Google Scholar]
- Zhang, L.; Li, M.; Li, X.; Yan, P.; Zhang, L.; Han, W. Summer pruning improves the branch growth and tea quality of tea trees (Camellia sinensis). Acta Physiol. Plant. 2021, 43, 65. [Google Scholar] [CrossRef]
- Mamun, M.; Hoque, M.M.; Ahmed, M. Effects of different pruning operations on the incidence of red spider mite of tea in bangladesh. Bangl. J. Bot. 2016, 45, 247–251. [Google Scholar]
- Lu, L.; Luo, W.; Zheng, Y.; Jin, J.; Liu, R.; Lv, Y.; Ye, Y.; Ye, J. Effect of different pruning operations on the plant growth, phytohormones and transcriptome profiles of the following spring tea shoots. Beverage Plant Res. 2022, 2, 12. [Google Scholar] [CrossRef]
- Kandiah, S. Studies on the physiology of pruning tea 1. turnover of resources in relation to pruning. Tea Quart. 1971, 42, 89–100. [Google Scholar]
- Arkorful, E.; Hu, S.; Zou, Z.; Yu, Y.; Chen, X.; Li, X. Metabolomic Analyses Provide New Insights into Signaling Mechanisms for Nutrient Uptake by Lateral Roots of Pruned Tea Plant (Camellia sinensis). J. Agric. Food Chem. 2020, 68, 7890–7903. [Google Scholar] [CrossRef]
- Sun, X.; Ye, Y.; Ma, Q.; Guan, Q.; Jones, D.L. Variation in enzyme activities involved in carbon and nitrogen cycling in rhizosphere and bulk soil after organic mulching. Rhizosphere 2021, 19, 100376. [Google Scholar] [CrossRef]
- Salam, A.K.; Katayama, A.; Kimura, M. Activities of some soil enzymes in different land use systems after deforestation in hilly areas of West Lampung, South Sumatra, Indonesia. Soil Sci. Plant Nutr. 1998, 44, 93–103. [Google Scholar] [CrossRef]
- Elkhlifi, Z.; Iftikhar, J.; Sarraf, M.; Ali, B.; Saleem, M.H.; Ibranshahib, I.; Bispo, M.D.; Meili, L.; Ercisli, S.; Kayabasi, E.T.; et al. Potential Role of Biochar on Capturing Soil Nutrients, Carbon Sequestration and Managing Environmental Challenges: A Review. Sustainability 2023, 15, 2527. [Google Scholar] [CrossRef]
- Arafat, Y.; Din, I.U.; Tayyab, M.; Jiang, Y.; Chen, T.; Cai, Z.; Zhao, H.; Lin, X.; Lin, W.; Lin, S. Soil Sickness in Aged Tea Plantation Is Associated With a Shift in Microbial Communities as a Result of Plant Polyphenol Accumulation in the Tea Gardens. Front. Plant Sci. 2020, 11, 601. [Google Scholar] [CrossRef]
- Nacke, H.; Thürmer, A.; Wollherr, A.; Will, C.; HoDac, L.; Herold, N.; Schöning, I.; Schrumpf, M.; Daniel, R. Pyrosequencing-Based Assessment of Bacterial Community Structure Along Different Management Types in German Forest and Grassland Soils. PLoS ONE 2011, 6, e17000. [Google Scholar] [CrossRef] [PubMed]
- Romaniuk, R.; Giuffré, L.; Costantini, A.; Nannipieri, P. Assessment of soil microbial diversity measurements as indicators of soil functioning in organic and conventional horticulture systems. Ecol. Indic. 2011, 11, 1345–1353. [Google Scholar] [CrossRef]
- Lan, Y.; Wang, S.; Zhang, H.; He, Y.; Jiang, C.; Ye, S. Intercropping and nitrogen enhance eucalyptus productivity through the positive interaction between soil fertility factors and bacterial communities along with the maintenance of soil enzyme activities. Land Degrad. Dev. 2023. [Google Scholar] [CrossRef]
- Huang, X.-F.; Chaparro, J.M.; Reardon, K.F.; Zhang, R.; Shen, Q.; Vivanco, J.M. Rhizosphere interactions: Root exudates, microbes, and microbial communities. Botany 2014, 92, 267–275. [Google Scholar] [CrossRef]
- Bag, S.; Mondal, A.; Banik, A. Exploring tea (Camellia sinensis) microbiome: Insights into the functional characteristics and their impact on tea growth promotion. Microbiol. Res. 2022, 254, 126890. [Google Scholar] [CrossRef]
- Li, Y.; Li, Z.; Li, Z.; Jiang, Y.; Weng, B.; Lin, W. Variations of rhizosphere bacterial communities in tea (Camellia sinensis L.) continuous cropping soil by high-throughput pyrosequencing approach. J. Appl. Microbiol. 2016, 121, 787–799. [Google Scholar] [CrossRef]
- Pramanik, P.; Phukan, M.; Ghosh, S.; Goswami, A.J. Pruned tea bushes secrete more root exudates to influence microbiological properties in soil. Arch. Agron. Soil Sci. 2018, 64, 1172–1180. [Google Scholar] [CrossRef]
- Jiang, Y.; Lin, X.; Khan, M.U.; Jiang, W.; Xu, Y.; Li, Z.; Lin, W. Tea pruning for the umbrella-shaped canopy can alleviate rhizosphere soil degradation and improve the ecosystem functioning of tea orchards. Catena 2023, 222, 106885. [Google Scholar] [CrossRef]
- Ye, J.H.; Hu, W.W.; Zhang, Q.; Wang, H.B.; Jia, X.L.; He, H.B. Correlation between soil characteristics of tea plantations and the growth and fresh leaf quality of wuyi tea (Camellia sinensis cv. Shuixian). Chin. J. Trop Crop 2020, 41, 1838–1846. [Google Scholar]
- Lu, K. Methods of Soil Agricultural Chemical Analysis; China Agricultural Science and Technology Press: Beijing, China, 2000. [Google Scholar]
- Li, Z.G.; Luo, Y.M.; Teng, Y. Soil and Environmental Microbiology Research Method; Science Press: Beijing, China, 2008. [Google Scholar]
- Choi, K.H.; Dobbs, F.C. Comparison of two kinds of Biolog microplates (GN and ECO) in their ability to distinguish among aquatic microbial communities. J. Microbiol. Meth. 1999, 36, 203–213. [Google Scholar] [CrossRef]
- Barua, D.N. Seasonal Dormancy in Tea (Camellia sinensis L.). Nature 1969, 224, 514. [Google Scholar] [CrossRef]
- Kumar, R.; Bisen, J.S.; Singh, M.; Bera, B. Effect of pruning and skiffing on growth and productivity of Darjeeling tea (Camellia sinensis L.). Int. J. Tech. Res. Appl. 2015, 3, 28–34. [Google Scholar]
- Yin, L.Q.; Xiao, X.; Luo, Q.X.; Xia, L.F.; Yang, Q.; Liang, M.Z. Effect of light pruning in different period on tea yield and quality of yunnan big-leaf varieties. Southwest China J. Agric. Sci. 2019, 32, 1034–1038. [Google Scholar]
- Tang, W.; Yang, H.; Wang, W.; Wang, C.; Pang, Y.; Chen, D.; Hu, X. Effects of Living Grass Mulch on Soil Properties and Assessment of Soil Quality in Chinese Apple Orchards: A Meta-Analysis. Agronomy 2022, 12, 1974. [Google Scholar] [CrossRef]
- Meddich, A.; Oufdou, K.; Boutasknit, A.; Raklami, A.; Tahiri, A.; Ben-Laouane, R.; Ait-El-Mokhtar, M.; Anli, M.; Mitsui, T.; Wahbi, S.; et al. Use of Organic and Biological Fertilizers as Strategies to Improve Crop Biomass, Yields and Physicochemical Parameters of Soil. In Nutrient Dynamics for Sustainable Crop Production; Meena, R., Ed.; Springer: Singapore, 2020; pp. 247–288. [Google Scholar]
- Cao, Y.; Cui, X.M.; Luo, X.Y. Effects of combine trimming with fertilizing on tea quality of yield and low performing tea garden. J. Anhui Agric. Sci. 2012, 40, 13311–13312. [Google Scholar]
- Zuber, S.M.; Villamil, M.B. Meta-analysis approach to assess effect of tillage on microbial biomass and enzyme activities. Soil Biol. Biochem. 2016, 97, 176–187. [Google Scholar] [CrossRef]
- Zielewicz, W.; Swędrzyński, A.; Dobrzyński, J.; Swędrzyńska, D.; Kulkova, I.; Wierzchowski, P.S.; Wróbel, B. Effect of Forage Plant Mixture and Biostimulants Application on the Yield, Changes of Botanical Composition, and Microbiological Soil Activity. Agronomy 2021, 11, 1786. [Google Scholar] [CrossRef]
- Sun, H.; Zhang, J.F.; Hu, Y.; Wang, L.; Chen, G.C. Response of soil catalase to cover of different stands. Bull. Soil Sci. 2016, 47, 605–610. [Google Scholar]
- Mn, N.; Winding, A. Microorganisms as Indicators of Soil Health; National Environmental Research Institute: Aarhus, Denmark, 2002. [Google Scholar]
- Badri, D.V.; Vivanco, J.M. Regulation and function of root exudates. Plant Cell Environ. 2009, 32, 666–681. [Google Scholar] [CrossRef]
- Bora, S.S.; Hazarika, D.J.; Gogoi, R.; Dullah, S.; Gogoi, M.; Barooah, M. Long-term pruning modulates microbial community structure and their functional potential in Tea (Camellia sinensis L.) soils. Appl. Soil Ecol. 2022, 176, 104483. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | MC | MX | Index | MC | MX |
|---|---|---|---|---|---|
| pH | 4.02 ± 0.01 b | 4.46 ± 0.01 a | TN (g/kg) | 1.15 ± 0.02 a | 1.10 ± 0.04 a |
| AN (mg/kg) | 122.10 ± 1.03 a | 114.59 ± 1.82 b | TP (g/kg) | 1.03 ± 0.03 a | 0.73 ± 0.02 b |
| AP (mg/kg) | 13.24 ± 0.48 b | 15.52 ± 0.62 a | TK (g/kg) | 5.62 ± 0.33 a | 5.84 ± 0.47 a |
| AK (mg/kg) | 101.41 ± 2.81 b | 111.29 ± 1.10 a | OM (g/kg) | 20.01 ± 0.02 b | 21.14 ± 0.09 a |
| Treatment | Simpson Index | Shannon Index | Chao 1 |
|---|---|---|---|
| MC | 0.999 ± 0.001 a | 4.525 ± 0.009 a | 0.913 ± 0.002 a |
| MX | 0.983 ± 0.001 b | 4.510 ± 0.028 a | 0.910 ± 0.006 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Q.; Zhang, Y.; Miao, P.; Chen, M.; Du, M.; Pang, X.; Ye, J.; Wang, H.; Jia, X. Effects of Pruning on Tea Tree Growth, Soil Enzyme Activity and Microbial Diversity. Agronomy 2023, 13, 1214. https://doi.org/10.3390/agronomy13051214
Zhang Q, Zhang Y, Miao P, Chen M, Du M, Pang X, Ye J, Wang H, Jia X. Effects of Pruning on Tea Tree Growth, Soil Enzyme Activity and Microbial Diversity. Agronomy. 2023; 13(5):1214. https://doi.org/10.3390/agronomy13051214
Chicago/Turabian StyleZhang, Qi, Ying Zhang, Pengyao Miao, Meihui Chen, Mengru Du, Xiaomin Pang, Jianghua Ye, Haibin Wang, and Xiaoli Jia. 2023. "Effects of Pruning on Tea Tree Growth, Soil Enzyme Activity and Microbial Diversity" Agronomy 13, no. 5: 1214. https://doi.org/10.3390/agronomy13051214
APA StyleZhang, Q., Zhang, Y., Miao, P., Chen, M., Du, M., Pang, X., Ye, J., Wang, H., & Jia, X. (2023). Effects of Pruning on Tea Tree Growth, Soil Enzyme Activity and Microbial Diversity. Agronomy, 13(5), 1214. https://doi.org/10.3390/agronomy13051214