1. Introduction
Indian mustard (
Brassica juncea L.), belonging to the family brassicaceae, is one of the important oilseed crops, and is currently ranked as the world’s third-most important oil seed crop in terms of production and area [
1]. In India, among the nine major oilseed crops, soybean (36%), groundnut (28%), and rapeseed and mustard (28% each) contribute to more than 90% of the total oilseed production in the country [
2]. Out of the total area under rapeseed and mustard production, more than 80% of the acreage is located in Rajasthan, Uttar Pradesh, Haryana, Madhya Pradesh, and Gujarat [
3]. Although India has a large land area (61.24 lakh ha) under oilseed
Brassica cultivation, with a total production of 10.11 million metric tons, it still imports 13.35 metric tons of oilseeds from other countries to meet its domestic requirements [
4,
5]. A major bottleneck in the productivity enhancement of Indian mustard is recurrent yield loss due to a range of biotic and abiotic stress factors. Most of the rapeseed and mustard crops in India are vulnerable to biotic constraints such as white rust, downy mildew, Alternaria blight, Sclerotinia rot, and powdery mildew [
6]. Out of these, white rust disease, incited by
Albugo candida (Pers. Ex. Lev.) Kuntze, is one of the most destructive diseases affecting brassicaceae crops globally, and has a wide host range of about 400 plant species around the world. Considering the significant yield losses in
Brassica, it is one of the top ten Oomycete pathogens [
7]. To date, around 17 distinct physiological races of
A. candida have been reported globally. Often the virulence of races significantly differs, making some of the races devastating for a particular species; for example, race 2 in
B. juncea, race 9 in
B. oleracea, race 7 in
B. rapa, race 1 in
Raphanus sativus, and race 5 in
Sisymbrium officinale [
8,
9,
10,
11]. The disease results in both localized and systemic infections that affect all aerial parts of the plant, including the cotyledons, leaves, stem, and inflorescence. The localized infection is characterized by the development of white to cream-colored zoosporangial pustules. On the other hand, systemic infection on meristems and inflorescences causes “stagheads”, or hypertrophied or malformed racemes, leading to no seed production [
10]. Stagheads (inflorescence nerves) may also appear during the latter part of the growing season as a result of meristematic host tissue contamination [
12]. In India, Australia, and Canada, the pathogen has been linked to yield losses in
B. juncea ranging from 20 to 60% [
13,
14]. The disease has the greatest impact in India because of the susceptibility of nearly all of the newly released commercially grown cultivars [
13]. While many chemicals and cultural techniques have been proposed to control this disease [
15,
16,
17], the alternative approach, such as the breeding of genetic resistance into the cultivars, is regarded as one of the most cost-effective and environmentally friendly methods for disease management.
The process of domestication and continuous selection for yield has resulted in a narrow genetic base for the
B. juncea cultivars. However, previous studies have identified resistance sources in the gene pool of oil brassica including the Indian mustard. Arora et al. reported resistance against six isolates collected from the northern regions of India in the Indian mustard variety Donskaja-IV and identified a single CC-NB-LRR protein-coding R gene (BjuWRR1) as providing resistance against white rust in the European variety [
18]. Another gene WRR12 has also been identified in
A. thaliana which confers resistance to
A. candida race 9 that infects
B. oleracea [
19]. Even though there are, at present, sources available for white rust resistance, the existing sources are often ineffective against new races of pathogens or multiple races of the same pathogen. To create new variability in
Brassica sp. for genetic resistance, three major approaches include: resynthesis, which is the process of incorporating diversity from a progenitor species; introgressing variation from a related species; and mutagenesis through physical or chemical methods [
20,
21]. The goal is to create novel genetic or phenotypic variation due to intergenomic recombination between the parent species’ chromosomes and other polyploidy-related outcomes [
22]. In addition, analysis using allozymes and genetic markers has further demonstrated that resynthesized genotypes are excellent diversity conduits for the
Brassica crop [
23,
24]. Prior to this, Hasan and Rahman (2018) [
25] created clubroot-resistant
B. juncea through resynthesis by mating two susceptible
B. nigra lines with a resistant genotype of
B. rapa. This resulted in the formation of the clubroot-resistant line of
B. juncea. Additionally, due to an infection brought on by
A. brassicae, resistance to the
Brassica leaf blight has been transferred from
B. hirta to
B. juncea [
26]. The identification of R-genes is the first and foremost step in developing disease-resistant varieties through breeding programs. Previous studies have shown that the white rust resistance in different
Brassica species can be governed by a single dominant gene, one or two dominant genes with epistatic effects, an additive dominant gene with epistatic effects, or a partial resistant single recessive gene
wpr [
27]. As a result of environmental and pathogenic variation, some R-genes interact differently with
Brassica genotypes in different environments. Previous work has identified a few genotypes with resistance, but dynamic changes in pathogen race composition and scanty screening with a small number of isolates have frequently resulted in short-lived host resistance in improved varieties, necessitating the identification and characterization of novel sources of white rust resistance from diverse genetic backgrounds. Therefore, understanding the interactions of
Brassica genotypes with
A. candida is essential in determining resistant genotypes with specific abilities to adapt and exploit for further improvements. Understanding the variability of responses of different germplasm to
A. candida infections is critical, as is strengthening the breeding programme to incorporate resistance genes against the white rust pathogen in oilseed
Brassica.
Field evaluation is the most commonly used method for identifying resistant sources; however, it is resource intensive and requires repeated testing to confirm the consistency of reactions, as there is a risk of disease escape due to low or poor inoculum concentration or potential, disease pressure, and other factors. Considering these problems, the current study aimed to identify and validate resistant sources in advanced introgressed, mutant, and resynthesized lines of B. juncea against the most prevalent virulent pathotypes of A. candida collected from major mustard growing hotspot locations across India under artificially inoculated conditions. Simultaneously, rigorous multilocational field testing was conducted in open fields to identify potent sources of resistance against the highly virulent isolates and races of A. candida. These sources can be employed as resistant donors in the backcross transfer of white rust resistance in popular mustard varieties for a successful mustard breeding program.
3. Results and Discussion
In the present study, a total of 194 introgression lines (ERJ) and 9 mutants, as well as 90 resynthesized
B. juncea lines, were screened at the cotyledonary leaf (CL) and true leaf (TL) growth stages under artificial conditions and at the adult plant stage in the field under natural conditions. As the expression of host resistance is reflected through the severity of white rust disease, based on the percent disease index of white rust, promising germplasm exhibiting immune or highly resistant reactions against thirteen isolates of
A. candida was identified. Among the tested germplasm (194 ILs, 90 RBJs, and 9 mutant lines), a wide range of reactions were observed, which varied from being immune or fully free from the disease with the NN type interaction phenotype (no infection and no sign of pustules on either side of the leaf surface) to being highly susceptible (
Table 1).
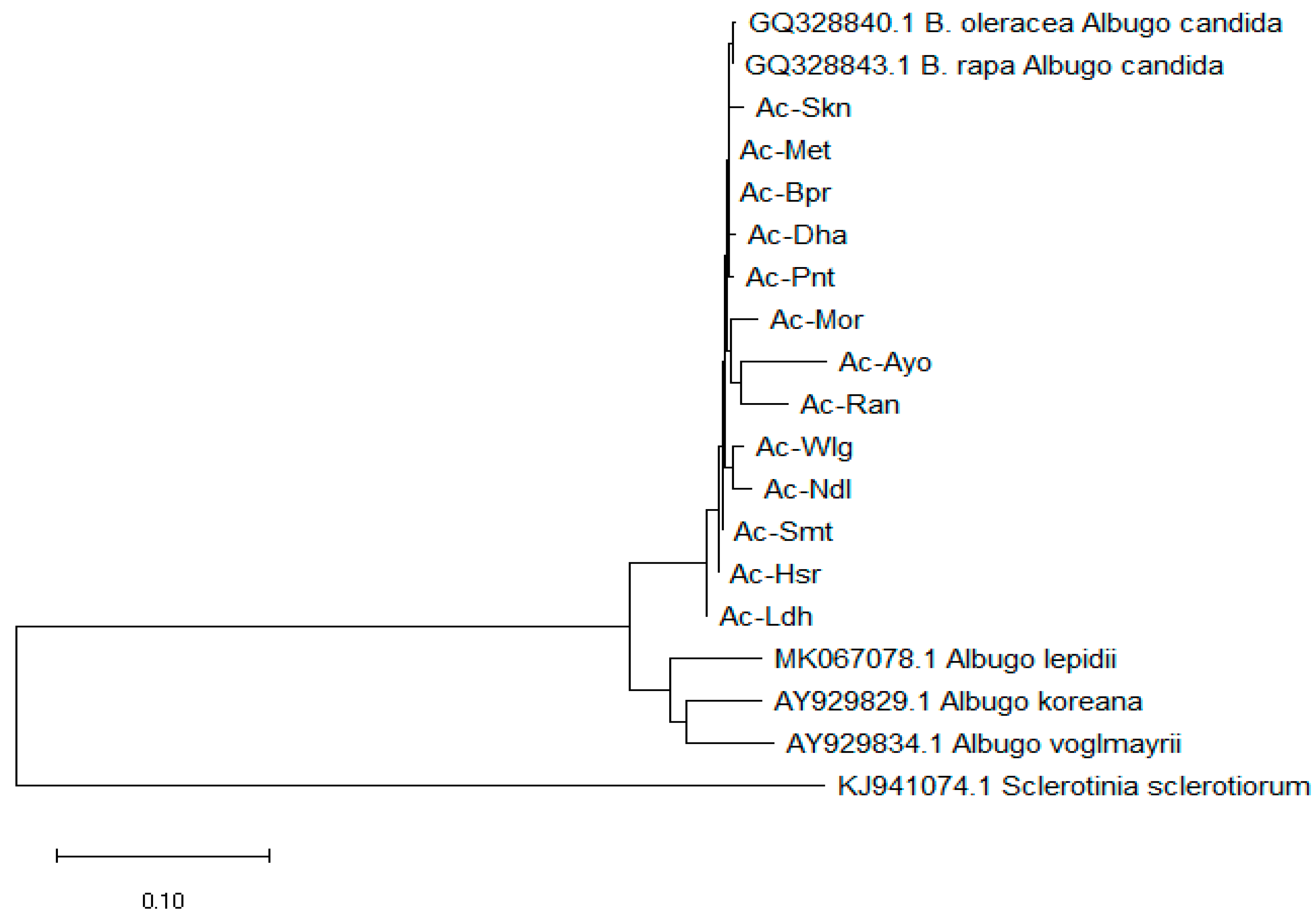
3.1. Genetic Variability of the Selcted Isolates
ITS sequences of thirteen isolates from major mustard growing locations with a nucleotide length of 542 bases were used for sequence alignments. The evolutionary history was inferred by using the Neighbour Joining method based on the Tamura-Nei model [
35]. Based on the phylogenetic tree Ac-Ayo, Ac-Ran and Ac-Mor were clubbed together to form a monophyletic group (
Figure 3). Similarly, Ac-Wlg and Ac-Ndl were clustered together in a monophyletic group and formed another cluster. However, Ac-Skn, Ac-Bpr, Ac-Met, Ac-Smt, Ac-Pnt, Ac-Hsr, Ac-Ldh and Ac-Dha formed different clusters. According to the phylogenetic tree, Ac-Ldh was found out to be the most diffferent as compared to the other clusters that have derived from the same node. The molecular characterization of these 13 A. candida isolates with the ITS gene clearly showed significant variability among different selected isolates. This also indicates association of geographical regions and the variability amid A. candida isolates. Such variability in the population might be the result of different selection pressure on pathogen due to availability of a host species for survival and infection and difference in agro-climatic conditions of 13 diverse mustard growing regions of India. Therefore, identification of the genetic diversity provides a good resolution in the differentiation of A. candida isolates.
3.2. Screening of Introgressed Lines
Introgressed lines of
B. juncea were developed through interspecific-crossing between the wild plant,
Diplotaxis erucoides (D
eD
e, 2n = 14) which shows immunity against multiple isolates of
A. candida and therefore is used as the donor parent, and the highly susceptible cultivar, RLM 198 (AABB, 2n = 36) as the recurrent parent (
Table 2). Among the 194 ILs of
B. juncea, ERJ 39, ERJ 12, and ERJ 15 showed significant potential for white rust resistance against the highest number of isolates of the pathogen (
Figure 4). Both ERJ 39 and ERJ 12 demonstrated an immune response (PDI = 0) characterized by NN type interaction phenotypes (no infection and no sign of pustules on either side of the leaf surface) across all the growth phases, including the cotyledonary and true leaf stages under artificial infection and the adult plant stage in field trials, against five isolates of
A. candida:
Ac-Ndl,
Bpr,
Pnt,
Ldh, and
Ran (
Table 3 and
Table 4). Because white rust resistance is governed by a single gene, such genetic resistance at all the growth stages is practically ideal for breeding approaches, which is highly desirable for further introgression and commercial cultivation of
B. juncea L. varieties in India.
Introgressed line ERJ 39 also expressed an immune response against
Ac-Hsr and
Ac-Met at both cotyledonary and true leaf stages. For
Ac-Mor and
Skn isolates, immunity was only observed for the cotyledonary phase, and moderate resistance was observed with PDI values of 7.2 and 7.5 at true leaf stage, respectively (
Table 3). Furthermore, immunity to
Ac-Smt was also found in ERJ 39 at the cotyledonary and adult plant stages but not at the true leaf stage, where moderate resistance was seen (PDI = 8.7%). Such an immune response in the cotyledonary phase to
A. candida infection is highly desired because the plants can avoid systemic spread of the disease and staghead formations caused by hypertrophy or hyperplasia, which can lead to significant yield losses [
13]. This introgressed line showed immunity to resistance responses at all plant growth stages to a range of isolates obtained from different agro-climatic zones, thus making it superior to all of the other introgression lines evaluated.
In the case of ERJ-12, immunity was recorded at the cotyledonary and true leaf stages against
Ac-Met and
Ac-Mor when tested artificially. However, for
Ac-Hsr,
Ayo,
Dha,
Smt, and
Skn, immunity was only seen at the cotyledonary stage, but high (PDI = 1–5%) to moderate resistance (PDI = 6–10%) was demonstrated at the true leaf and mature plant stages. This type of shift in resistance to plant diseases linked to the transition from the juvenile to the adult phase is demonstrated in
A. thaliana as well [
36]. Coelhoe et al. [
37] made similar observations in resistance at different growth stages in
B. oleracea against downy mildew, suggesting that the age of the plant is also one of the factors on which resistance levels can depend based on the ability of the test pathogen to infect.
ERJ 15, another potential germplasm, expressed a complete immune reaction against four isolates, Ac-Bpr, Pnt, Ldh, and Ran, at all three growth stages of the plant, i.e., cotyledonary, true leaf, and adult plant stages.
This germplasm also demonstrated immune responses for isolates
Ac-Met,
Hsr, and
Ayo in artificial trials at both the cotyledonary and true leaf stages. In the case of
Ac-Smt and
Ac-Ndl isolates, immunity was observed at the cotyledonary stage. However, at the true leaf stage, the same lines reacted as highly resistant (PDI = 1–5%). Under field experiments, however, PDI scores of 0 (immune) and 3.3 (high resistance) were reported for
Ac-Smt and
Ndl, respectively. For
Ac-Dha, which was sourced from the southern part of India, a susceptible reaction was seen at the cotyledonary stage (PDI = 11.2) and moderate resistance (PDI = 9.3) at the true leaf stage. Moreover, in the case of
Ac-Wlg, susceptible to moderately susceptible responses were observed across all the plant growth stages (
Table 3 and
Table 4). This illustrates the contrast in virulence between the northern and southern Indian races of the test pathogen.
In addition to the aforementioned introgression lines, which displayed total immunity against the majority of the
A. candida isolates at all growth stages, 35 genotypes were discovered to produce an immune resistance level of responses against specific pathotypes of white rust under artificial as well as natural field conditions (
Table 5). These genotypes can be exploited in mustard improvement programs for developing resistance cultivars against white rust disease. Among these 35 genotypes, ERJ 108, and ERJ 157 showed immunity against
Ac-Ldh specifically; furthermore, ERJ 159, ERJ 13, and ERJ 32 were also shown to be immune against
Ac-Ran,
Pnt, and
Bpr, respectively (
Table 5). Such a response to the disease can be used to study the differences between host and pathogen and to comprehend the biological specialization that exists among the isolates collected from diverse plant hosts in various places.
Screening available ILs for resistance revealed a higher percentage of susceptibility to the white rust pathogen. Out of 194 germplasm samples tested under
B. juncea, 39.69% expressed a highly susceptible reaction at the cotyledonary stage, while 40.48% and 40.08% showed high susceptibility at the true leaf and adult plant stages, respectively. Only 5.37% of the total ERJ displayed immunity at the cotyledonary stage, 3.68% at the true leaf stage, and 3.41% at the mature plant stage. Under artificial inoculation, high resistance (PDI = 1–5%) was observed in 7.09% of total ILs at the cotyledonary stage and 5.37% at the true leaf stage, while only 6.52% of ILs demonstrated high resistance at the adult stage in fields. Additionally, restricted sporulation occasionally supplemented with necrosis or chlorosis with a FN interaction phenotype was found in 7.52% of ERJs at the cotyledonary stage, 7.05% at the true leaf stage, and 6.83% at the adult plant stage (
Figure 5), indicating moderate resistance (PDI 6–10%) in the introgressed lines.
This section may be divided by subheadings. It should provide a concise and precise description of the experimental results, their interpretation, as well as the experimental conclusions that can be drawn.
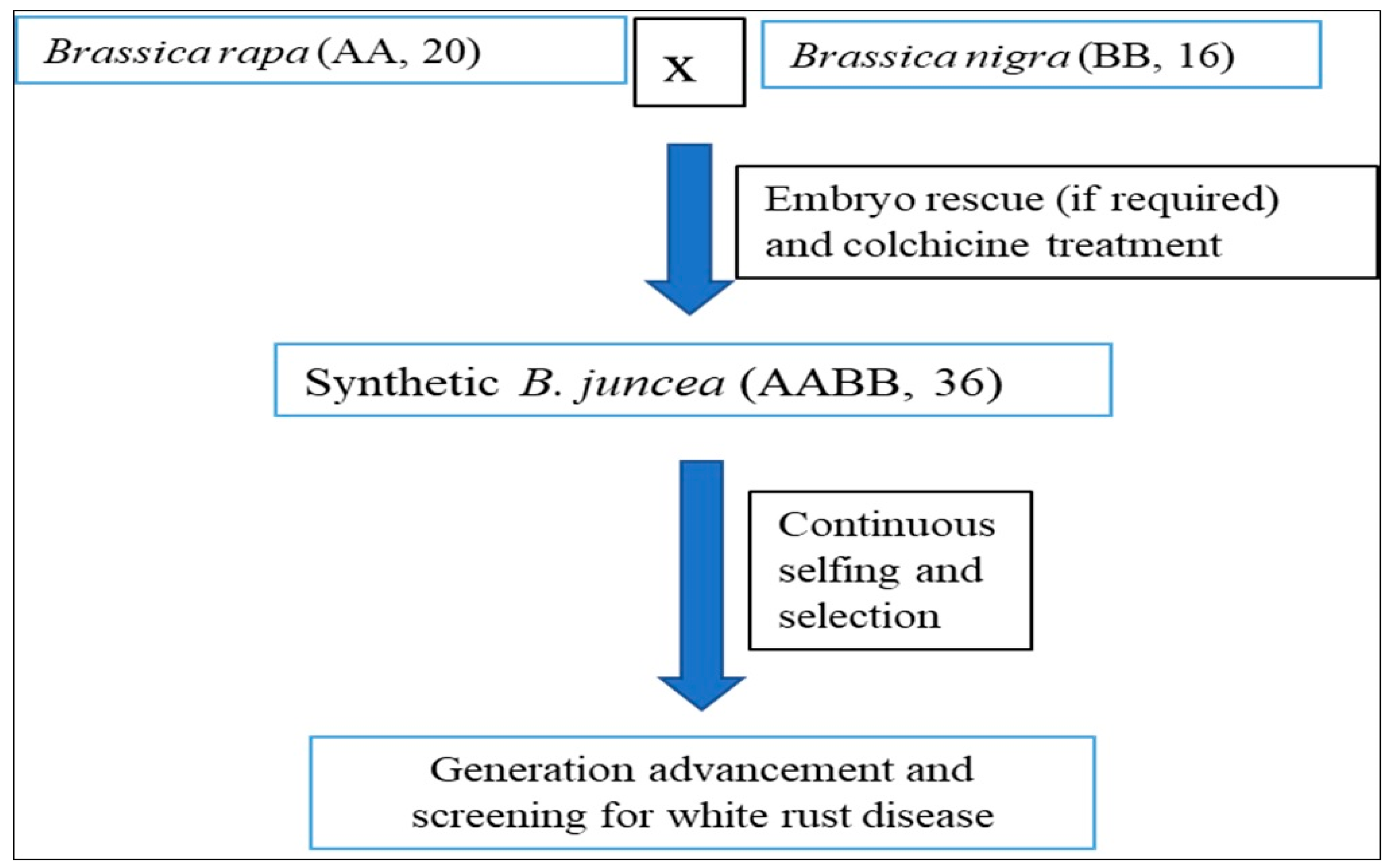
3.3. Screening of Mutant and Resynthesized B. juncea Lines
In the present study, 90 resynthesized and 9 advanced mutant lines of
B. juncea developed at ICAR-NIPB, New Delhi, and ICAR-DRMR, Bharatpur, were also evaluated for identifying novel host resistance sources against the white rust (
A. candida) pathogen. The resynthesized lines were developed by crossing
B. rapa (AA, 2n = 20) and
B. nigra (BB, 2n = 16), which showed various levels of resistance for different
A. candida isolates, followed by the colchicine treatment to develop amphidiploid
B. juncea (AABB, 2n = 36) (
Table 2). Meanwhile, mutant lines were derived from the Indian mustard cultivar RH 749 following treatment with different doses of gamma rays and ethyl methane sulfonate (EMS). Among the tested germplasms, the mutant lines DRMR 18-36-12 and DRMR 18-37-13 and the resynthesized line RBJ 18 were found to express immunity and showed no sign of disease against multiple races of the
A. candida pathogen (
Figure 6).
DRMR 18-36-12 showed complete immunity to four isolates,
Ac-Ndl,
Bpr,
Pnt, and
Ran, at all three growth stages of plants, including cotyledonary, true leaf, and adult plant growth stages. Additionally, under artificial inoculation with
A. candida, no immune reaction (PDI = 0%) was observed for
Ac-Met,
Hsr, and
Mor isolates at both cotyledonary and true leaf stages. For the isolate
Ac-Ldh, immunity was expressed at the cotyledonary stage, while the true leaf and adult plant stages both had high resistance, with PDI scores of 3.2 and 3.8, respectively. This was in contrast to isolates
Ac-Smt and
Skn, where immunity was observed at the true leaf and adult plant stages but not at the cotyledonary stage, as PDI scores of 4.3% and 4.0% were observed in these genotypes, respectively (
Table 6 and
Table 7). Another mutant line, DRMR 18-37-13, also expressed complete immunity against
Ac-Ndl,
Bpr,
Ldh, and
Ran at cotyledonary, true leaf, and adult plant stages. Moreover, under controlled conditions, an immune response was observed against
Ac-Met,
Hsr, and
Mor. For isolates
Ac-Pnt,
Ayo,
Smt, and
Skn, immunity (PDI = 0) was also found at true leaf stage but not at cotyledonary stage, as high to moderate resistance was expressed at this phase of seedling growth. This germplasm, on the other hand, demonstrated susceptible (formation of numerous pustules on the lower surface of leaves covering 11–25% of the leaf area) to moderately susceptible (coalescing large, scattered pustules on the lower surface of leaves covering 26–50% of the leaf area) reactions for
Ac-Wlg and
Dha at all the three stages of plant development.
Under both in vitro and in vivo field trials, one of the most promising resynthesized lines, RBJ 18, demonstrated total immunity to six isolates of A. candida: Ac-Ndl, Bpr, Pnt, Ldh, Smt, and Ran. This line demonstrated an immune response with Ac-Met, Mor, and Skn at true leaf stage but a high (1–5%) to moderate (6–10%) resistance response when tested at cotyledonary leaf stage.
Moreover, under natural field conditions, RBJ 18 also displayed an immune response at the adult plant stage against
Ac-Skn. For isolates
Ac-Dha and
Ac-Wlg, a highly resistant response was observed at the cotyledonary stage; however, at the true leaf stage, moderate resistance (PDI = 9.3) and a susceptible reaction (PDI = 17.8) were found, respectively (
Table 6). Resynthesized lines such as RBJ 38 and RBJ 90 expressed immunity specifically against
Ac-Smt, while mutant line DRMRSJ 1 was found immune only to
Ac-Pnt at the cotyledonary, true leaf, and adult plant stages (
Table 8). Such germplasm can be explored for resistance exclusive to one race and for host differential-related studies. Unexpectedly, none of the mutant and RBJ lines showed complete immunity against
Ac-Wlg and
Dha, which shows a limited variability in resistance of these resynthesized lines to varying degrees of virulence as shown by different isolates across India. A total of 29 genotypes were discovered among the resynthesized lines of
B. juncea that displayed good levels of resistance, of which the immune reaction was represented by only 4.55% of the total germplasm in the field, 5.05% at true leaf stage, and 5.67% at cotyledonary stage. Immunity aside, 11.97% of germplasm exhibited high resistance (PDI = 5%) at the cotyledonary stage and 9.17% at the true leaf stage under artificial inoculation. However, at the adult stage in fields, 10.98% of the germplasm was found to be highly resistant. In addition, 12.59% of genotypes were categorized as moderately resistant (PDI = 6–10%) at the cotyledonary stage, 10.80% at the true leaf stage, and 11.62% at the adult plant stage (
Figure 7).
3.4. Relation between Cotyledonary, True Leaf Stage, and Adult Plant Resistance
In the trial run under artificial inoculation in the growth chamber and under natural field conditions, the relationship between cotyledonary, true leaf, and adult plant resistance was also investigated. Therefore, to ascertain this relation between resistance at the cotyledonary, true leaf, and adult growth phases of plants, a correlation analysis was conducted. Cotyledonary stage resistance is highly valued in
A. candida infections, as plants can become infected during the early stages of growth, which ultimately results in systemic infections that lead to hypertrophy, hyperplasia, extensive distortion of the affected tissues, and staghead formations [
14]. However, cotyledon resistance and adult plant resistance are not well correlated and vary with germplasm and the screening method used [
38].
In the current study, 17 genotypes out of a total of 194 introgressed lines expressed complete immunity against one or more isolates at all three plant growth stages, including cotyledonary, true leaf, and mature plant. This form of resistance at all stages is typically ideal in plant breeding because the individual genes provide high levels of resistance against a larger range of races and have a tendency to be robust [
39]. Despite the similar trend between the resistance levels at the three growth stages, four ILs, ERJ 5, ERJ 13, ERJ 40, and ERJ 110, showed moderate (PDI = 6–10%) to high resistance responses (PDI = 1–5%) at the adult stage under field conditions, which is a slightly lower level of resistance as compared to the seedling stage in greenhouse conditions, where they exhibited immunity against different isolates of the pathogen (
Table 3). This might be because a larger mixed inoculum builds up occurs in outdoor settings as opposed to growth chambers, where tests are conducted with a single isolate under controlled environmental circumstances. It is therefore advisable to test the resistance in the field as well as in controlled conditions with prominently virulent isolates of the pathogen to identify the robust sources of host resistance. Four ILs, namely, ERJ 15, ERJ 38, ERJ 39, and ERJ 109, were found to express immunity at the cotyledonary stage and adult plant stage but not at the true leaf stage. On the contrary, eight germplasms, ERJ 7, ERJ 16, ERJ 38, ERJ 40, ERJ 90, ERJ 103, ERJ 109, and ERJ 110, were identified to express immunity (PDI = 0) at true leaf and adult plant stages but not at cotyledonary stages (
Table 3 and
Table 4). This shows there is a considerably larger association between resistance at the true leaf and adult plant stages than there is between resistance at the cotyledonary stages. Correlation studies have further supported this, as the true leaf stage and adult growth stage of plants in introgression lines showed a significantly strong correlation for disease resistance against
A. candida (r = 0.734, n = 912,
p < 0.001), followed by the correlation between adult plant stage and cotyledonary stage (r = 0.444, n = 912,
p < 0.001) and the true leaf stage and cotyledonary stage (r = 0.402, n = 912,
p < 0.001).
Among mutant and resynthesized lines, nine genotypes were shown to exhibit complete immunity at all three plant growth stages, including cotyledonary, true leaf, and mature plant. Only one mutant line, DRMRSJ 1, was found to express high resistance (PDI = 2.7%) at the adult stage under field conditions; however, under greenhouse conditions, it exhibited immunity against the various isolates of the pathogen. Immunity (PDI = 0) was found in 10 genotypes, including three mutant lines (DRMR 18-36-12, DRMRDJ 1, and DRMRSJ 4) and seven resynthesized lines (RBJ 18, RBJ 10A, RBJ 38, RBJ 19, RBJ 10C, RBJ 17, and RBJ 89), at true leaf and adult plant stages, while high resistance (PDI = 1–5%) to susceptible reaction (PDI = 11–25%) was observed at cotyledonary stage (
Table 6 and
Table 7). There was no genotype that expressed immunity at the cotyledonary and adult plant stages but not at the true leaf stage, implying that most genotypes that were immune at the adult, cotyledonary or both stages were also immune at the true leaf stage. This probably suggests a strong correlation between the true leaf stage and the adult plant stage. This was further confirmed by the correlation studies, as a significantly strong correlation for disease resistance against
A. candida was found for true leaf stage and adult plant stage (r = 0.797, n = 696,
p < 0.001), followed by correlation between the adult plant stage and cotyledonary stage (r = 0.441, n = 696,
p < 0.001) and the true leaf stage and cotyledonary stage (r = 0.399, n = 696,
p < 0.001).
It could therefore be concluded from the correlation analysis of ERJ, mutant, and RBJ lines that the genotype responses during the early growth stages, particularly the true leaf stage of the young seedling, could be dependably employed as a quick assay for determining genetic resistance against A. candida pathogenesis. Although this makes it possible to lower the expense of multi-location trials and eliminates the possibility of weather-related variability when screening adult plant resistance in the field, it is not reliable all the time, as depending on the conditions, there are always chances for disease escape. Moreover, a shift in plant developmental stage from juvenile to adult might also trigger a different kind of resistance response. Therefore, for a conclusive result, rigorous testing of the genotypes under both controlled and natural conditions at all growth stages is preferred.
3.5. Analysis of Variance
Under both field and in vitro conditions, the fit test results for the GGE model for
B. juncea revealed a very significant main impact of environment (E) and genotype (G) and genotype by environment interaction (GEI) (
p < 0.001). (
Table 9 and
Table 10). An analysis of variance (ANOVA) for introgressed lines indicated that under controlled conditions, 15.8% of the total sum of squares (SS) was explained by the effect of genotype and 14.8% and 69.4% was attributable to the environment (E) main effects and genotype by environment interaction (GEI), whereas under field conditions, 18.6% was represented by genotype, 17.7% by environment, and 63.7% of the total SS by GEI. Meanwhile, for mutants and resynthesized
B. juncea under artificial conditions, 28.2% of the total SS was explained by genotype, while 16.6% and 55.2% were represented by environment and the interaction between genotype and environment, respectively. Similarly, under field trials, 21.4% of total SS was represented by genotype, and the rest, 11.9% and 66.8%, were denoted by environment and GEI, respectively. These total SS show the variation in genotypes for the white rust disease index across isolates and locations. While the environmental component (locations) coupled with weather conditions influence the genotypes’ performance at various locations, variation due to G or GE interactions is a measure for the strains’ response across the environments and locations. The higher percentage of GE shows that adaptabilities are preferred by the genotype. Therefore, because there is significantly more refinement in the variances for G and GE than in a single location, multi-environment trials (METs) realize the virtue of germplasm for both temporal and geographic stability [
40].
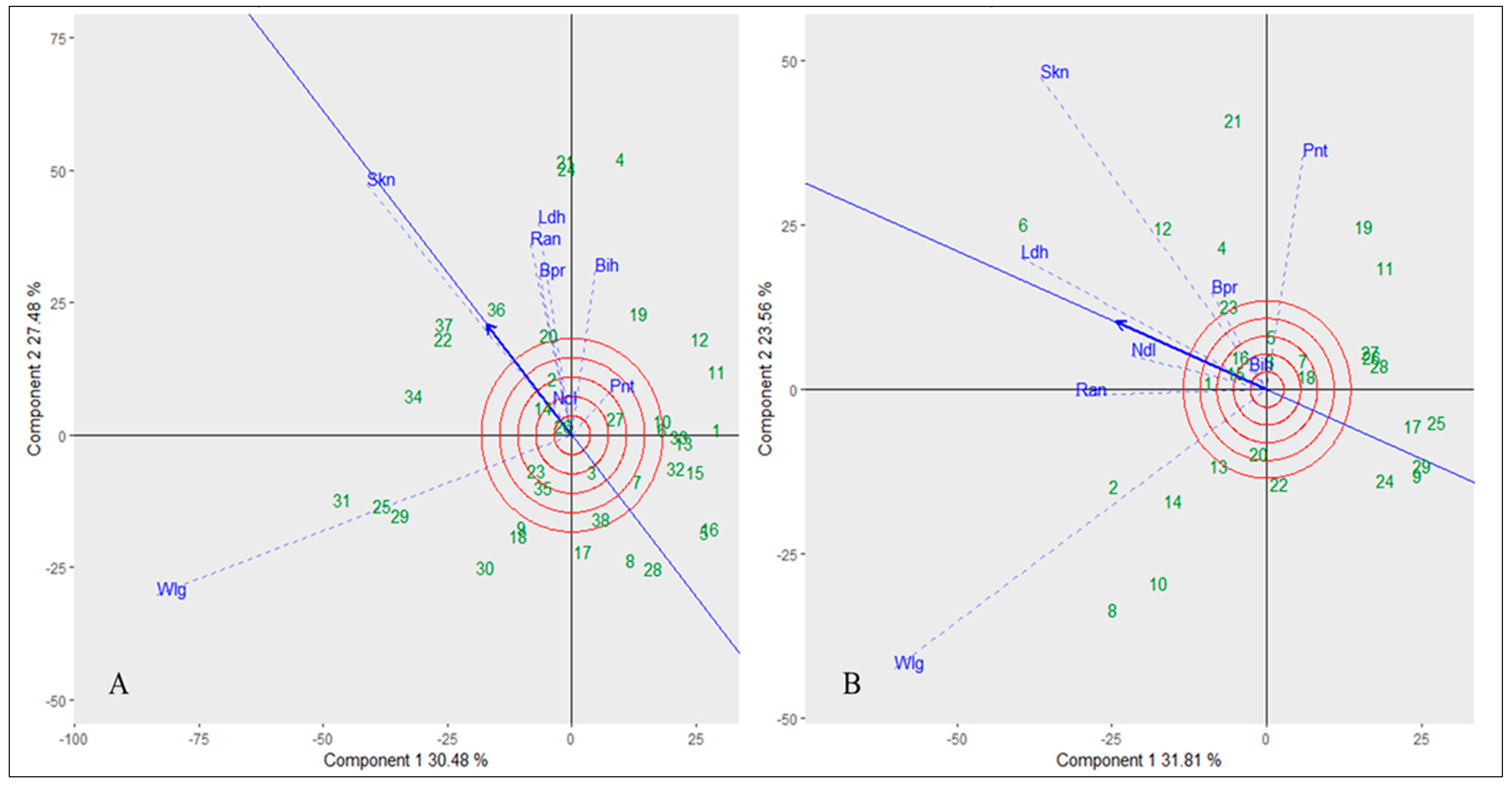
3.6. Interaction Studies between Genotypes and Hotspot Locations
Principal component analysis (PCA) was carried out for the multi-locational trials to comprehend the impact of location-specific environments in terms of the resistance testing against
A. candida isolates exclusively for the introgression, mutant, and resynthesized lines of
B. juncea. The pattern of environments in connection to genotypes for white rust severity was visualized using GGE biplots based on symmetric scaling of genotype and environment. The GGE biplot graph used to evaluate white rust severity in the present study accounted for 57.9% of the total variation due to G + GE for ERJ and 55.3% for RBJ. In GGE biplot, environment vector lines connect the plot origin and markers for the environments. The correlation coefficient between two environments or genotypes is related to the angle between their vectors. While a perpendicular angle denotes no association and an obtuse angle demonstrates a negative correlation, an acute cosine angle suggests a positive correlation between the environment(s) or genotype(s). Therefore, according to the cosine of angles of environment vectors for ERJ, Wellington and S. K. Nagar (SKN) showed a positive correlation, while Wellington and Pantnagar were negatively correlated, suggesting they both have differing agro-climatic conditions for the development of the white rust disease (
Figure 8A). However, in the case of RBJ and mutant lines, no correlation could be seen between Wellington and SKN as the environment vectors in this case formed a right cosine angle; instead, a positive correlation was seen between Ranchi and Wellington, while, similarly to ERJ, a negative correlation was found between Wellington and Pantnagar (
Figure 8B).
In order to choose widely adaptable genotypes, an ideal habitat should have both the power of discrimination (the capacity of the environment to discriminate among genotypes) and representativeness (how well the location represents the mega-environment). The discriminating power of the location is directly correlated with the length of the location vector, whereas the power of representativeness is directly correlated with the angle between the location vector and AEC abscissa, with smaller angles being preferred. As indicated in
Figure 7, Wellington and SKN were the most discriminating environments for both introgression and resynthesized lines, whereas Delhi and Bihar were the least discriminating for ERJ, mutants, and RBJ. In terms of representativeness, Ludhiana was the most representative of the test environment for mutants and RBJ, while SKN formed the smallest angle with the AEC abscissa, thus representing the mega environment for ERJ. Usually for genotypes, the interpretation in the GGE biplot is that the performance of a genotype in an environment is better than average if the angle between its vector and the environment’s vector is <90°; it is poorer than average if the angle is >90°; and it is near average if the angle is about 90°. However, in this study, the opposite trend was followed, as lower values were considered to be resistant because a higher disease score indicates greater susceptibility. Therefore, in the case of introgressed lines, genotypes 5, 8, 16, and 28 representing ERJ 12, ERJ 15, ERJ 39 and ERJ 109 were the most resistant germplasms that performed better as compared to other genotypes and showed better stability across the environments, especially ERJ 39 which showed multiple resistance across the trials. Meanwhile for resynthesized lines, genotypes 9, 17, 25 representing RBJ 18, RBJ 40, DRMRSJ 4 and mutants 24 and 29 represented by DRMR 18-37-13 and DRMR 18-36-12 were found to be more stable and perform well against
A. candida isolates at diverse locations, especially resynthesized and mutant line RBJ 18 and DRMR 18-36-12, respectively, which showed complete immunity and stability across multiple isolates and locations (
Figure 8).
4. Conclusions
Albugo candida is known to be a notorious biotroph responsible for significant economical and yield losses in B. juncea L. throughout the world. Recommended management strategies, such as employing systemic fungicides to combat this disease, not only seem to be ineffective against this disease but also affect the environment negatively. As a result, identifying host resistance may be the best alternative approach to combating this major pathogenic threat. Resistant sources can be further utilized to introgress the resistant genes and produce durable resistance. Mutagen-induced novel variations also add to the scope of screening for resistance. The present investigation was focused on the generation of putative resistance sources that would be suitable for Indian conditions among introgressed, resynthesized, and mutant B. juncea L. lines against prominently virulent isolates of A. candida. To the best of our knowledge, introgressed lines (ERJ 39, ERJ 12, and ERJ 15), resynthesized line RBJ 18, and mutant lines (DRMR 18-36-12 and DRMR 18-37-13) have all been identified as novel sources of resistance against multiple isolates of A. candida at all plant growth stages, including cotyledonary, true leaf, and adult plant stages. Among the others, ILs such as ERJ 108, ERJ 157, ERJ 159, ERJ 13, and ERJ 32 specifically showed resistance against single isolates. Similarly, mutant lines DRMRSJ 1 and RBJ, RBJ 38, and RBJ 90 expressed specific immunity to a single race of A. candida. The study also revealed a positive correlation between true leaf and adult plant stages; however, with the cotyledonary stage, both true leaf and adult plant stages showed a weak correlation. Moreover, one cannot rely solely on artificial or field testing for a conducive result as there is sometimes the possibility of disease escape and, therefore, the germplasm might show resistant reactions. GGE biplot analysis in this study allowed efficient assessment of the resistance of ERJ, mutants, and RBJ lines to white rust disease across environments. The method allowed for the selection of ideal genotypes based on their adaptability and stability to various agro-climatic zones and environments. The value of such genetic resistances in terms of yield advantage under varying environmental conditions, which could have the yield penalty caused by white rust, is very high. More such studies on the tripartite interaction between genotype, isolate, and environment will be much more useful in minimizing yield loss due to white rust in a particular area. The potential sources of resistance among Brassica germplasms identified in our study will have a practical impact on further identification and molecular mapping of the resistance gene(s) or QTLs and their marker-assisted incorporation into the leading cultivars. These lines also could be introgressed into commercial cultivars which have already broken down their resistance and sustainable management of white rust in Indian mustard.


 ,
, 













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}