
Effects of Iron Nanoparticles Addition on Bacterial Community and Phytotoxicity in Aerobic Compost of Pig Manure

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Synthesis of G-nFe
2.3. Characterization
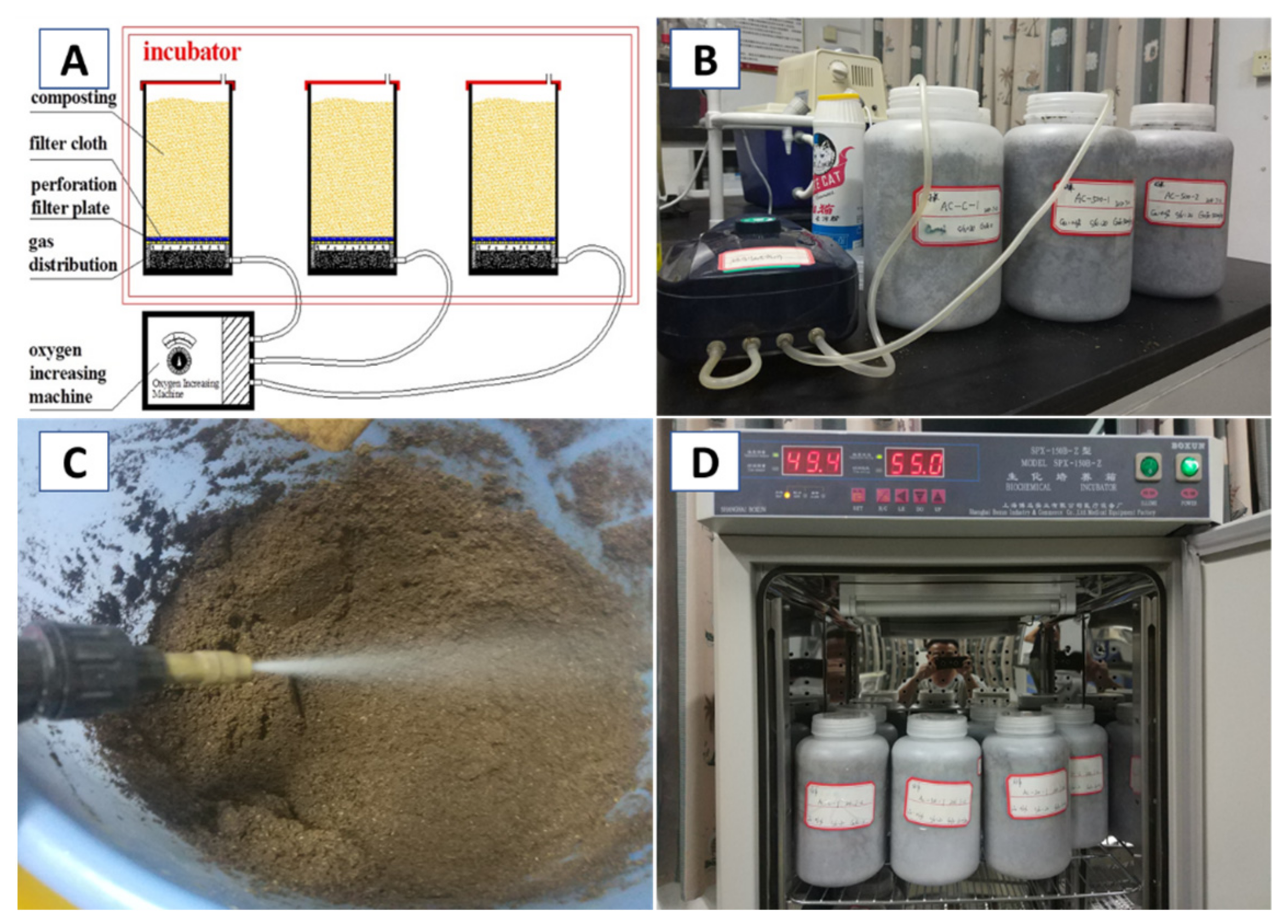
2.4. Thermophilic Aerobic Composting
2.5. Statistical Analysis
3. Results and Discussion
3.1. Characterization of the G-nFe and Compost Mixture
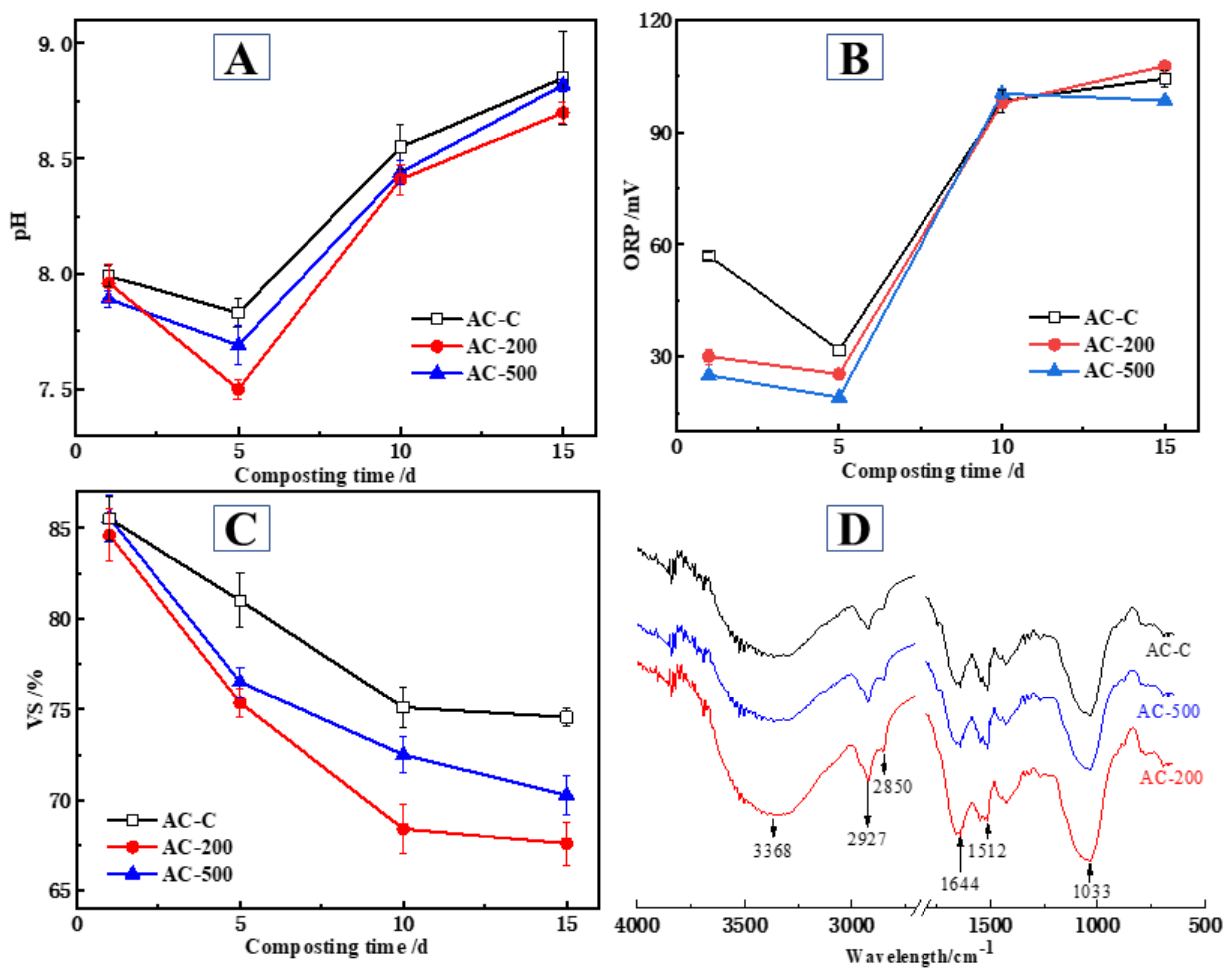
3.2. Temporal Changes in pH, ORP, VS, and FTIR during Composting
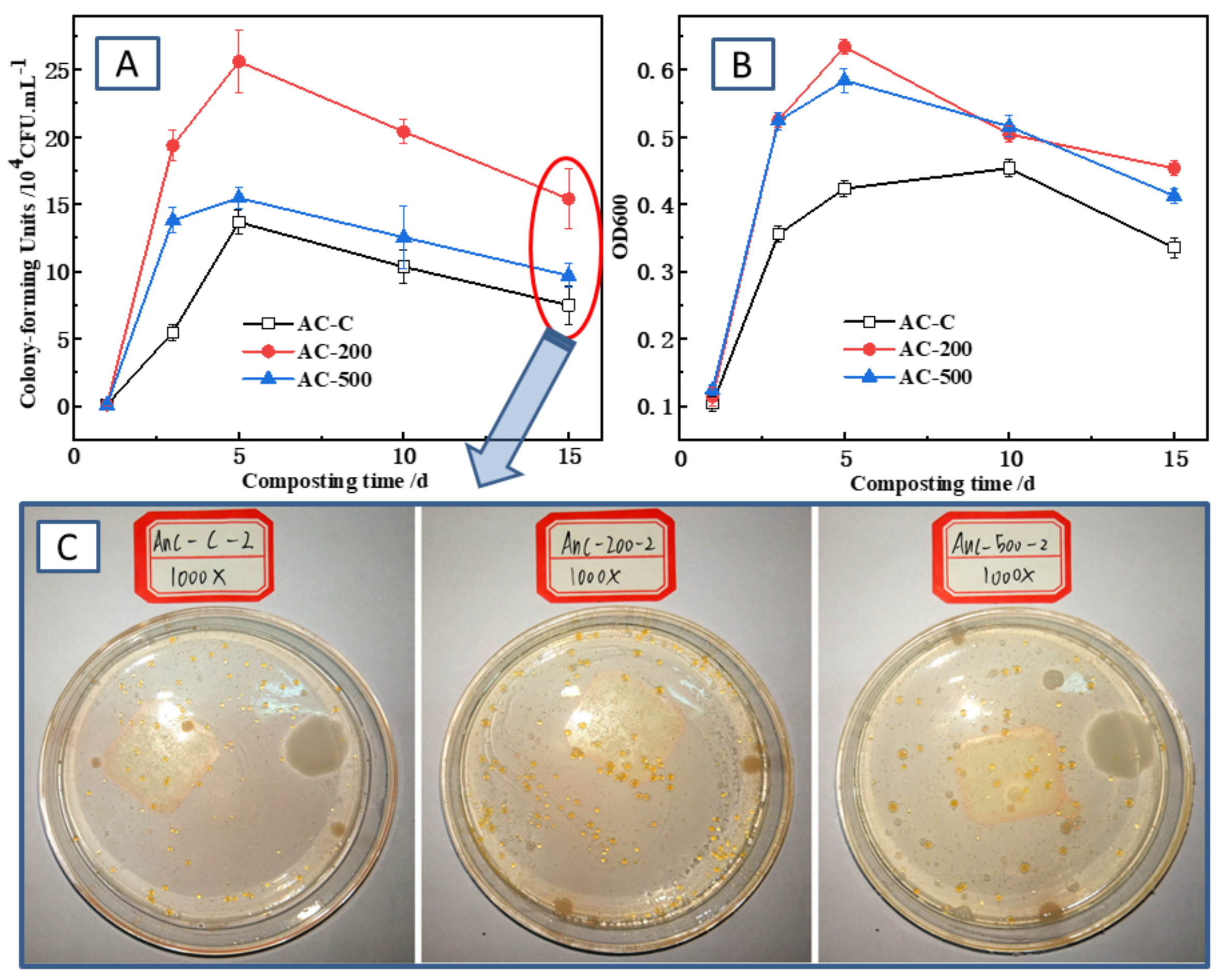
3.3. Temporal Changes in OD600 and CFU during Composting
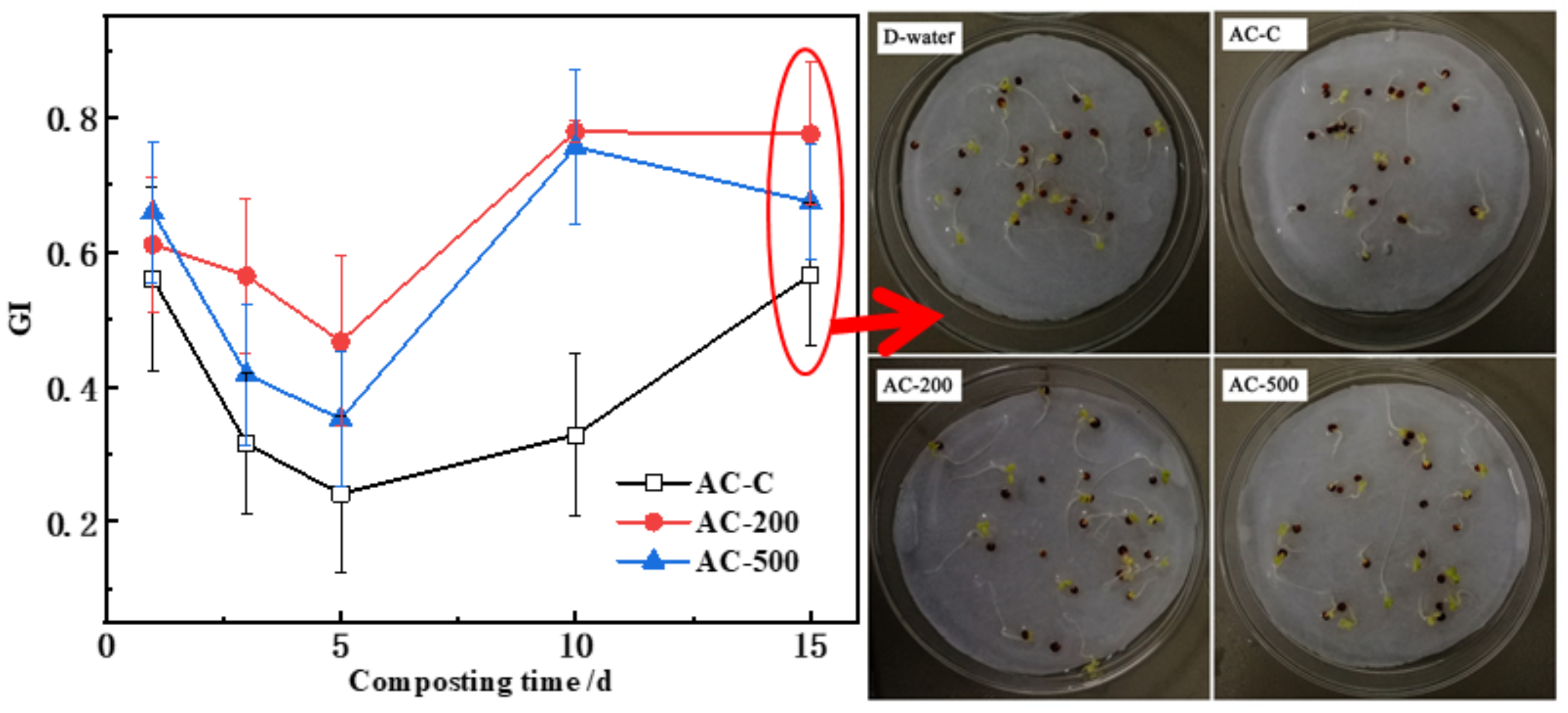
3.4. Temporal Changes in GI during Composting
3.5. Analysis of Bacteriological Changes during Composting
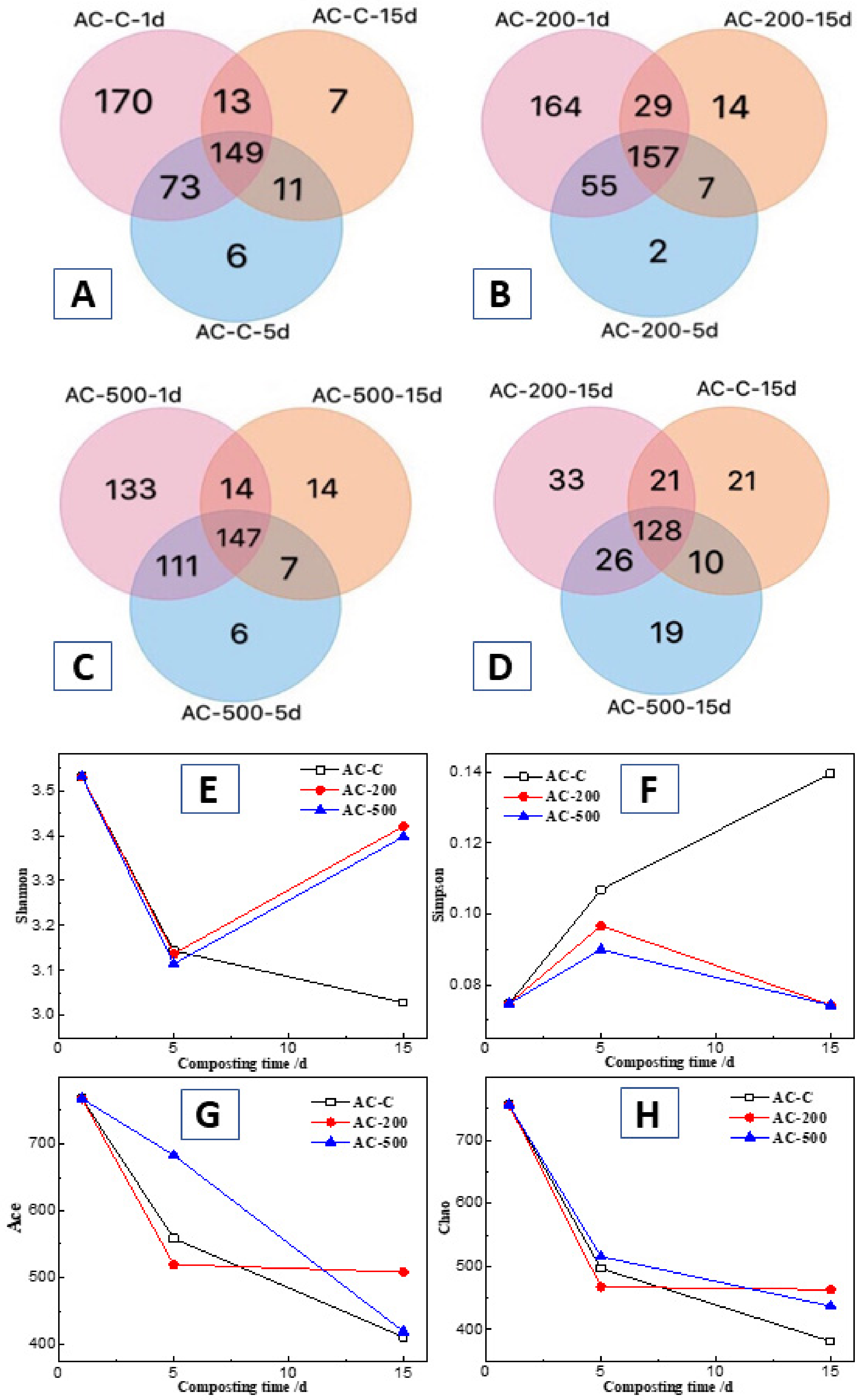
3.5.1. OTUs and Alpha Diversity of Compost Bacterial
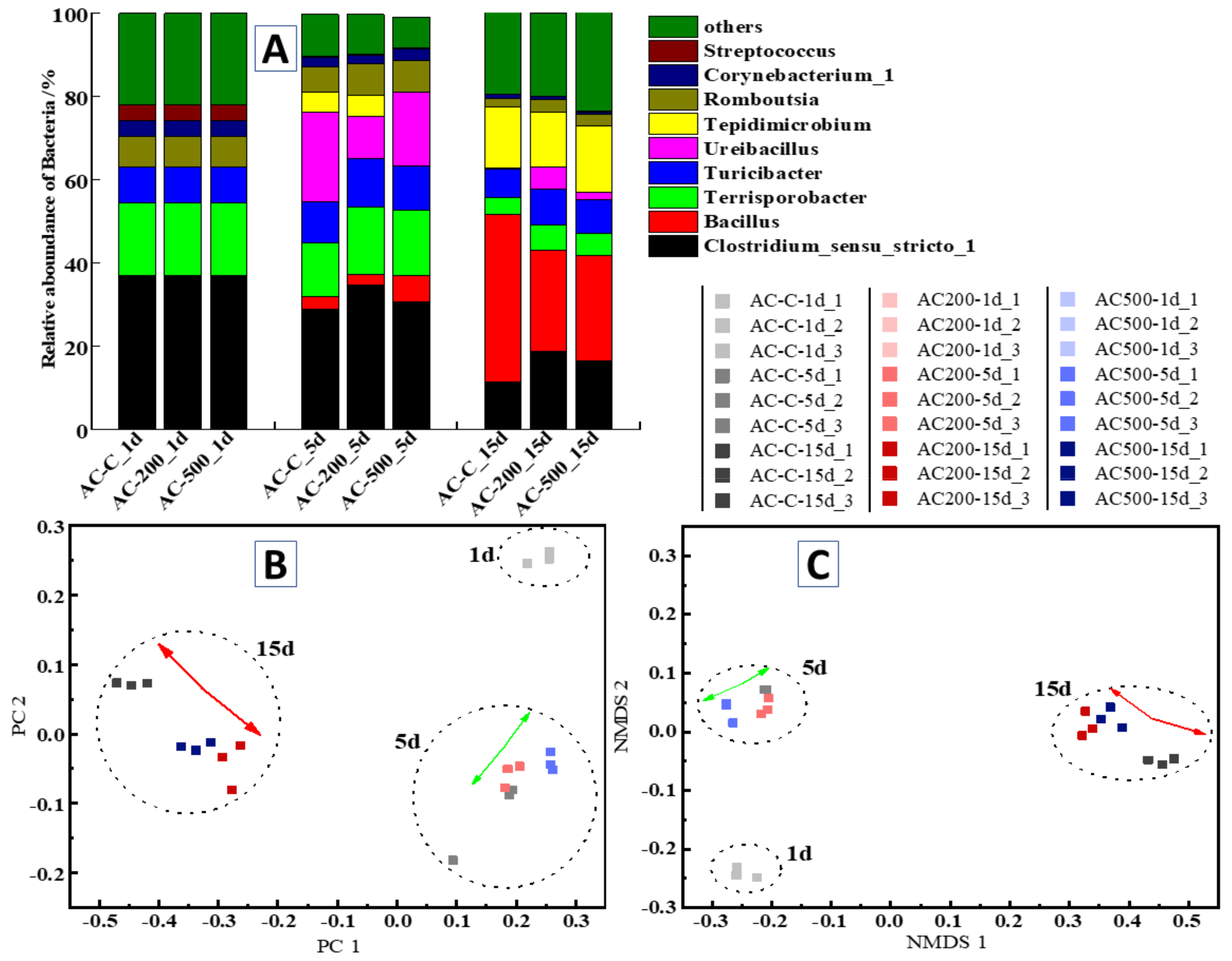
3.5.2. Temporal Variation in the Abundance of Dominant Bacterial Communities
3.5.3. Similarities and Differences between Bacterial Communities
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Stefaniuk, M.; Oleszczuk, P.; Yong, S.O. Review on nano zerovalent iron (nZVI): From synthesis to environmental applications. Chem. Eng. J. 2016, 287, 618–632. [Google Scholar] [CrossRef]
- Yi, Y.; Tu, G.; Tsang, P.; Xiao, S.; Fang, Z. Green synthesis of iron-based nanoparticles from extracts of Nephrolepis auriculata and applications for Cr(VI) removal. Mater. Lett. 2019, 234, 388–391. [Google Scholar] [CrossRef]
- Khatami, M.; Alijani, H.; Fakheri, B.; Mobasseri, M.; Heydarpour, M.; Farahani, Z.; Khan, A. Super-paramagnetic iron oxide nanoparticles (SPIONs): Greener synthesis using Stevia plant and evaluation of its antioxidant properties. J. Clean. Prod. 2019, 208, 1171–1177. [Google Scholar] [CrossRef]
- Luo, F.; Yang, D.; Chen, Z.; Megharaj, M.; Naidu, R. The mechanism for degrading Orange II based on adsorption and reduction by ion-based nanoparticles synthesized by grape leaf extract. J. Hazard. Mater. 2015, 296, 37–45. [Google Scholar] [CrossRef]
- Fang, L.; Chen, Z.; Megharaj, M.; Naidu, R. Simultaneous removal of trichloroethylene and hexavalent chromium by green synthesized agarose-Fe nanoparticles hydrogel. Chem. Eng. J. 2016, 294, 290–297. [Google Scholar]
- Christina, L. Pharmaceuticals. China’s lakes of pig manure spawn antibiotic resistance. Sci. Total Environ. 2015, 347, 704. [Google Scholar]
- Pan, X.; Qiang, Z.; Ben, W.; Chen, M. Residual veterinary antibiotics in swine manure from concentrated animal feeding operations in Shandong Province, China. Chemosphere 2011, 84, 695–700. [Google Scholar] [CrossRef]
- Zitnick, K.K.; Shappell, N.W.; Hakk, H.; Desutter, T.M.; Khan, E.; Casey, F.X.M. Effects of liquid swine manure on dissipation of 17β-estradiol in soil. J. Hazard. Mater. 2011, 186, 1111–1117. [Google Scholar] [CrossRef]
- Li, L.; Xu, Z.; Wu, J.; Tian, G. Bioaccumulation of heavy metals in the earthworm Eisenia fetida in relation to bioavailable metal concentrations in pig manure. Bioresour. Technol. 2010, 101, 3430–3436. [Google Scholar] [CrossRef]
- Ziemer, C.J.; Bonner, J.M.; Cole, D.; Vinjé, J.; Constantini, V.; Goyal, S.; Gramer, M.; Mackie, R.; Meng, X.J.; Myers, G. Fate and transport of zoonotic, bacterial, viral, and parasitic pathogens during swine manure treatment, storage, and land application. J. Anim. Sci. 2010, 88, E84–E94. [Google Scholar] [CrossRef]
- Fang, H.; Han, L.; Zhang, H.; Long, Z.; Cai, L.; Yu, Y. Dissemination of antibiotic resistance genes and human pathogenic bacteria from a pig feedlot to the surrounding stream and agricultural soils. J. Hazard. Mater. 2018, 357, 53. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.; Hao, Y.; Shen, M.; Zhao, Q.; Li, Q.; Hu, J. Impacts of supplementing chemical fertilizers with organic fertilizers manufactured using pig manure as a substrate on the spread of tetracycline resistance genes in soil. Ecotoxicol. Environ. Saf. 2016, 130, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Wan, N.; Hwang, E.Y.; Cheong, J.G.; Choi, J.Y. A Comparative Evaluation of Maturity Parameters for Food Waste Composting. Compos. Sci. Util. 1999, 7, 55–62. [Google Scholar]
- Huang, G.F.; Wong, J.W.C.; Wu, Q.T.; Nagar, B.B. Effect of C/N on composting of pig manure with sawdust. Waste Manag. 2004, 24, 805–813. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Wei, Y.; Zheng, J.; Xin, Z.; Zhong, W. The behavior of tetracyclines and their degradation products during swine manure composting. Bioresour. Technol. 2011, 102, 5924–5931. [Google Scholar] [CrossRef]
- Wang, Q.; Li, R.; Cai, H.; Awasthi, M.K.; Zhang, Z.; Wang, J.J.; Ali, A.; Amanullah, M. Improving pig manure composting efficiency employing Ca-bentonite. Ecol. Eng. 2016, 87, 157–161. [Google Scholar] [CrossRef]
- Ran, X.; Awasthi, M.K.; Li, R.; Park, J.; Pensky, S.M.; Quan, W.; Wang, J.J.; Zhang, Z. Recent developments in biochar utilization as an additive in organic solid waste composting: A review. Bioresour. Technol. 2017, 246, 203–213. [Google Scholar]
- Ye, K.; Yan, Z.; Chen, Z.; Megharaj, M.; Naidu, R. Impact of Fe and Ni/Fe nanoparticles on biodegradation of phenol by the strain Bacillus fusiformis (BFN) at various pH values: Biomass, bioenergy, biowastes, conversion technologies, biotransformations, production technologies. Bioresour. Technol. 2013, 136, 588–594. [Google Scholar]
- Jiang, C.; Liu, Y.; Chen, Z.; Megharaj, M.; Naidu, R. Impact of iron-based nanoparticles on microbial denitrification by Paracoccus sp. strain YF1. Aquat. Toxicol. 2013, 142–143, 329–335. [Google Scholar] [CrossRef]
- Liu, Y.; Li, S.; Chen, Z.; Megharaj, M.; Naidu, R. Influence of zero-valent iron nanoparticles on nitrate removal by Paracoccus sp. Chemosphere 2014, 108, 426–432. [Google Scholar] [CrossRef]
- Yang, W.-q.; Zhuo, Q.; Chen, Q.; Chen, Z. Effect of iron nanoparticles on passivation of cadmium in the pig manure aerobic composting process. Sci. Total Environ. 2019, 690, 900–910. [Google Scholar] [CrossRef] [PubMed]
- Bertoldi, M.D.; Vallini, G.; Pera, A. The biology of composting: A review. Waste Manag. Res. 1983, 1, 157–176. [Google Scholar] [CrossRef]
- Jiang, J.; Huang, H.; Huang, Y.; Liu, X.; Liu, D. Relationship between maturity and microbial communities during pig manure composting by phospholipid fatty acid (PLFA) and correlation analysis. J. Environ. Manag. 2018, 206, 532–539. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Yin, X.; Mao, H.; Chu, C.; Tian, Y. Changes in structure and function of fungal community in cow manure composting. Bioresour. Technol. 2018, 255, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Saludes, R.B.; Iwabuchi, K.; Kayanuma, A.; Shiga, T. Composting of dairy cattle manure using a thermophilic–mesophilic sequence. Biosyst. Eng. 2007, 98, 198–205. [Google Scholar] [CrossRef]
- Baddi, G.A.; Hafidi, M.; Cegarra, J.; Alburquerque, J.A.; Gonzálvez, J.; Gilard, V.; Revel, J.C. Characterization of fulvic acids by elemental and spectroscopic (FTIR and 13 C-NMR) analyses during composting of olive mill wastes plus straw. Bioresour. Technol. 2004, 93, 285–290. [Google Scholar] [CrossRef]
- Kothary, M.H.; Chase, T., Jr.; Macmillan, J.D. Levels of Aspergillus fumigatus in air and in compost at a sewage sludge composting site. Environ. Pollut. 1984, 34, 1–14. [Google Scholar] [CrossRef]
- Changha, L.; Jee Yeon, K.; Won, I.L.; Nelson, K.L.; Jeyong, Y.; Sedlak, D.L. Bactericidal effect of zero-valent iron nanoparticles on Escherichia coli. Environ. Sci. Technol. 2008, 42, 4927–4933. [Google Scholar]
- Wang, Z.; Lee, Y.-H.; Wu, B.; Horst, A.; Kang, Y.; Tang, Y.J.; Chen, D.-R. Anti-microbial activities of aerosolized transition metal oxide nanoparticles. Chemosphere 2010, 80, 525–529. [Google Scholar] [CrossRef]
- Chanyasak, V.; Katayama, A.; Hirai, M.F.; Mori, S.; Kubota, H. Effects of compost maturity on growth of komatsuna (brassica rapa var. pervidis) in neubauer’s pot. Soil Sci. Plant Nutr. 1983, 29, 239–250. [Google Scholar] [CrossRef]
- Kato, K.; Miura, N. Effect of matured compost as a bulking and inoculating agent on the microbial community and maturity of cattle manure compost. Bioresour. Technol. 2008, 99, 3372–3380. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Fang, Z.; Cheng, W.; Tsang, P.E.; Zhao, D. Ageing decreases the phytotoxicity of zero-valent iron nanoparticles in soil cultivated with Oryza sativa. Ecotoxicology 2016, 25, 1202–1210. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Shigeru, S. Understanding oxidative stress and antioxidant functions to enhance photosynthesis. Plant Physiol. 2011, 155, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Wang, Y.; Wen, Q. Effects of chlortetracycline on the fate of multi-antibiotic resistance genes and the microbial community during swine manure composting. Environ. Pollut. 2018, 237, 977–987. [Google Scholar] [CrossRef]
- Snell-Castro, R.; Godon, J.-J.; Delgenès, J.-P.; Dabert, P. Characterisation of the microbial diversity in a pig manure storage pit using small subunit rDNA sequence analysis. FEMS Microbiol. Ecol. 2005, 52, 229–242. [Google Scholar] [CrossRef]
- Yang, G.; Wang, J. Pretreatment of grass waste using combined ionizing radiation-acid treatment for enhancing fermentative hydrogen production. Bioresour. Technol. 2018, 255, 7–15. [Google Scholar] [CrossRef]
- Tang, J.; Li, X.; Zhao, W.; Wang, Y.; Cui, P.; Zeng, R.J.; Yu, L.; Zhou, S. Electric field induces electron flow to simultaneously enhance the maturity of aerobic composting and mitigate greenhouse gas emissions. Bioresour. Technol. 2019, 279, 234–242. [Google Scholar] [CrossRef]
- Zhang, R.; Gu, J.; Wang, X.; Li, Y.; Zhang, K.; Yin, Y.; Zhang, X. Contributions of the microbial community and environmental variables to antibiotic resistance genes during co-composting with swine manure and cotton stalks. J. Hazard. Mater. 2018, 358, 82–91. [Google Scholar] [CrossRef]
- Bernal, M.P.; Sommer, S.G.; Chadwick, D.; Qing, C.; Guoxue, L.; Michel, F.C. Chapter Three—Current Approaches and Future Trends in Compost Quality Criteria for Agronomic, Environmental, and Human Health Benefits. In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press: Cambridge, MA, USA, 2017; Volume 144, pp. 143–233. [Google Scholar]
- Arif, M.S.; Riaz, M.; Shahzad, S.M.; Yasmeen, T.; Ali, S.; Akhtar, M.J. Phosphorus-Mobilizing Rhizobacterial Strain Bacillus cereus GS6 Improves Symbiotic Efficiency of Soybean on an Aridisol Amended with Phosphorus-Enriched Compost. Pedosphere 2017, 27, 1049–1061. [Google Scholar] [CrossRef]
- Nakasaki, K.; Tran, L.T.H.; Idemoto, Y.; Abe, M.; Rollon, A.P. Comparison of organic matter degradation and microbial community during thermophilic composting of two different types of anaerobic sludge. Bioresour. Technol. 2009, 100, 676–682. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test | FCP | G-nFe (v/d w) | Aeration (Times/Day) | Aeration Time | Stirring Frequency /Artificial |
|---|---|---|---|---|---|
| AC-C | Yes | None added | 5 times/day | 5 min/times | 5 times/day |
| AC-200 | Yes | 200 mL/kg | 5 times/day | 5 min/times | 5 times/day |
| AC-500 | Yes | 500 mL/kg | 5 times/day | 5 min/times | 5 times/day |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, W.; Zhuo, Q.; Zhong, Y.; Chen, Q.; Chen, Z. Effects of Iron Nanoparticles Addition on Bacterial Community and Phytotoxicity in Aerobic Compost of Pig Manure. Agronomy 2023, 13, 1239. https://doi.org/10.3390/agronomy13051239
Yang W, Zhuo Q, Zhong Y, Chen Q, Chen Z. Effects of Iron Nanoparticles Addition on Bacterial Community and Phytotoxicity in Aerobic Compost of Pig Manure. Agronomy. 2023; 13(5):1239. https://doi.org/10.3390/agronomy13051239
Chicago/Turabian StyleYang, Wenqing, Qian Zhuo, Yuanping Zhong, Qinghua Chen, and Zuliang Chen. 2023. "Effects of Iron Nanoparticles Addition on Bacterial Community and Phytotoxicity in Aerobic Compost of Pig Manure" Agronomy 13, no. 5: 1239. https://doi.org/10.3390/agronomy13051239
APA StyleYang, W., Zhuo, Q., Zhong, Y., Chen, Q., & Chen, Z. (2023). Effects of Iron Nanoparticles Addition on Bacterial Community and Phytotoxicity in Aerobic Compost of Pig Manure. Agronomy, 13(5), 1239. https://doi.org/10.3390/agronomy13051239