
A Cationic Amino Acid Transporter NtCAT1 Promotes Leaf Senescence by the Accumulation of ABA in Nicotiana tabacum
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. RNA Extraction and Quantitative Real-Time PCR Analysis
2.3. Subcellular Localization
2.4. Generation of Transgenic Lines
2.5. Leaf Senescence Experiments
2.6. Assays for Antioxidant Enzymes Activity
2.7. GUS Staining
2.8. Physiological and Biochemical Measurements
2.9. Measurement of ABA Content and Free Amino Acids
2.10. Statistical Analysis
3. Results
3.1. Screening and Identification of NtCAT1 in Nicotiana tabacum
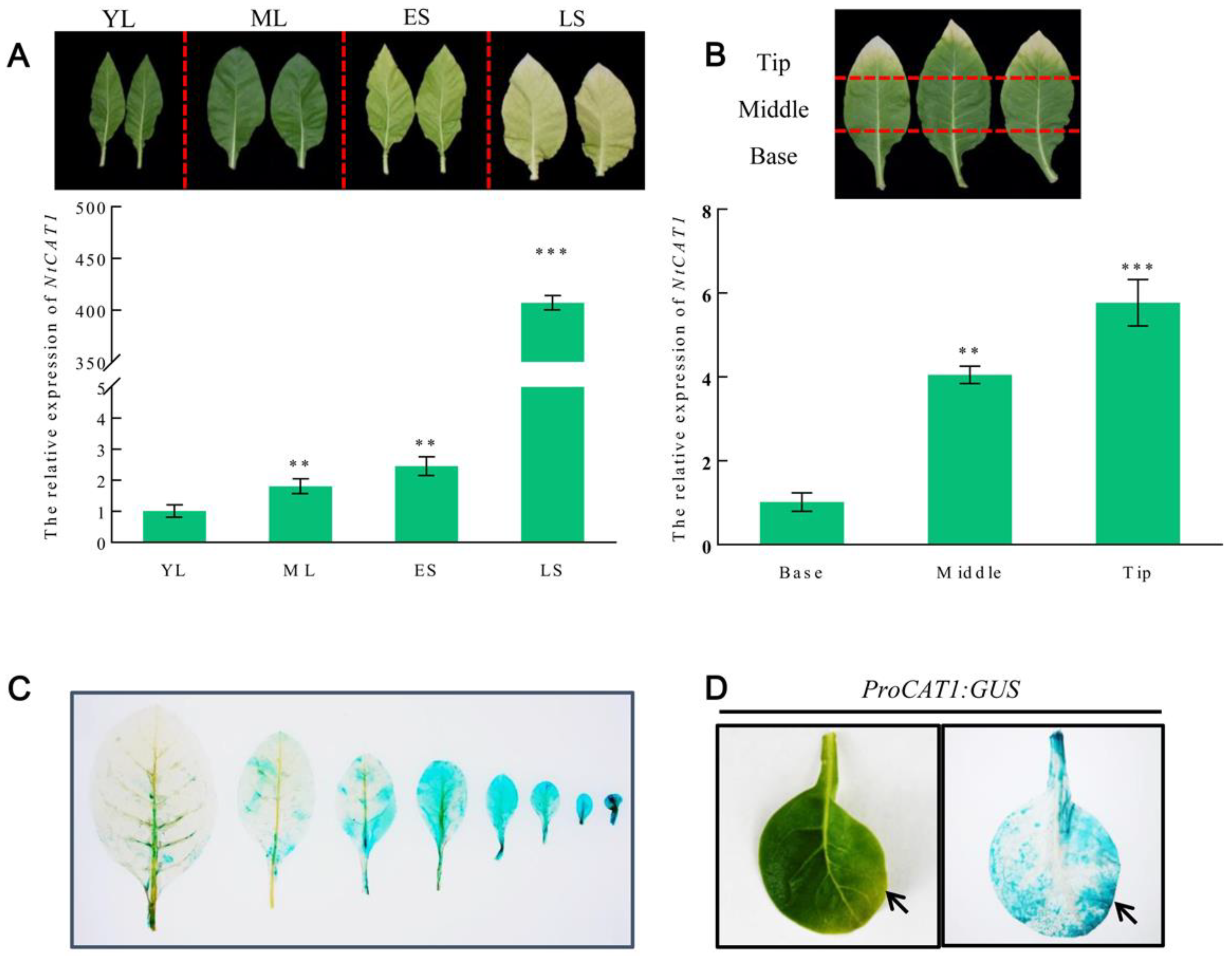
3.2. NtCAT1 Is a Senescence-Associated Gene during Natural Leaf Aging
3.3. Expression of NtCAT1 Is Induced by ABA Treatment
3.4. NtCAT1 Protein Is Localized in the Plasma Membrane
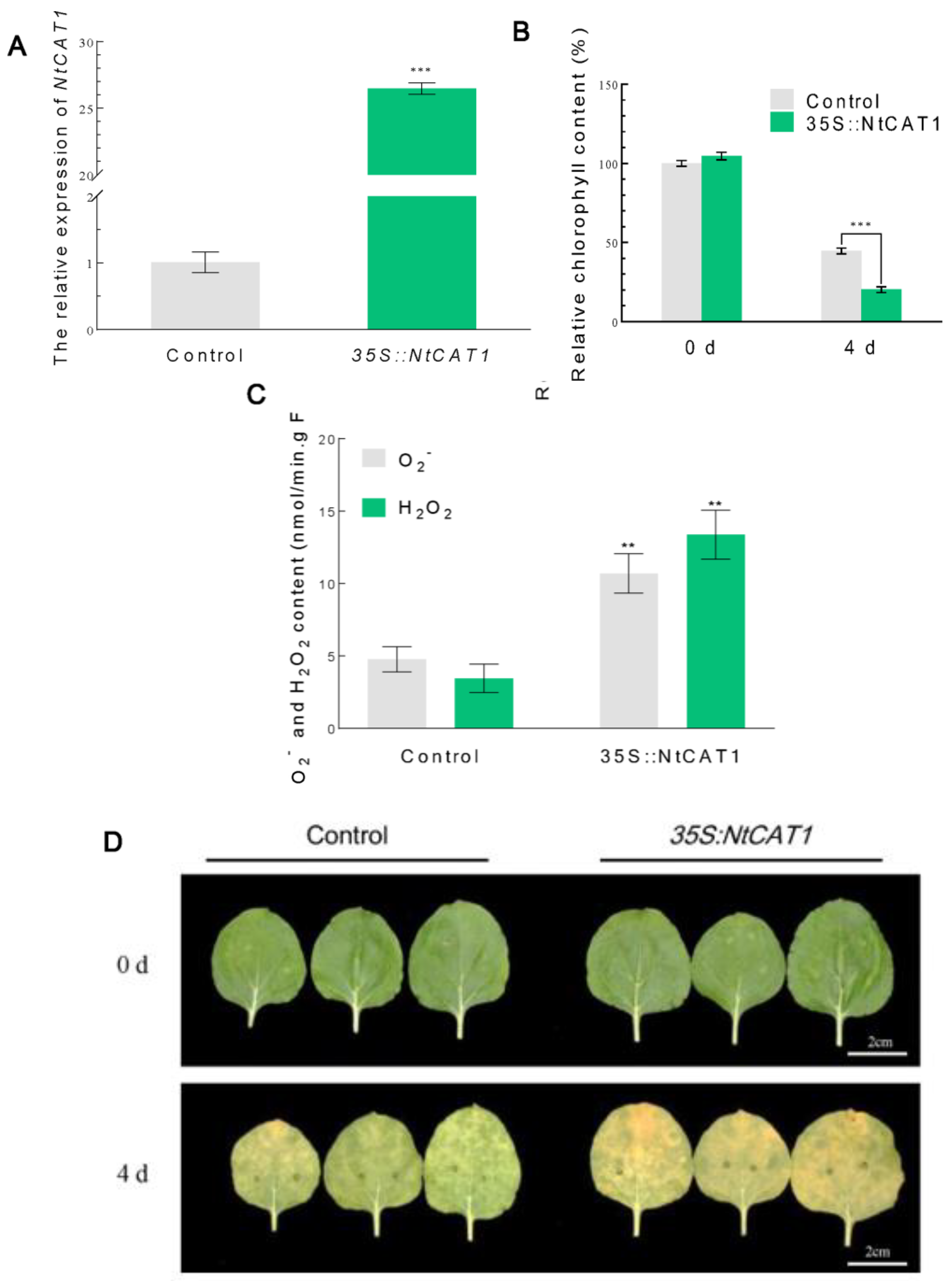
3.5. Transient Expression of NtCAT1 Accelerates Dark-Induced Leaf Senescence
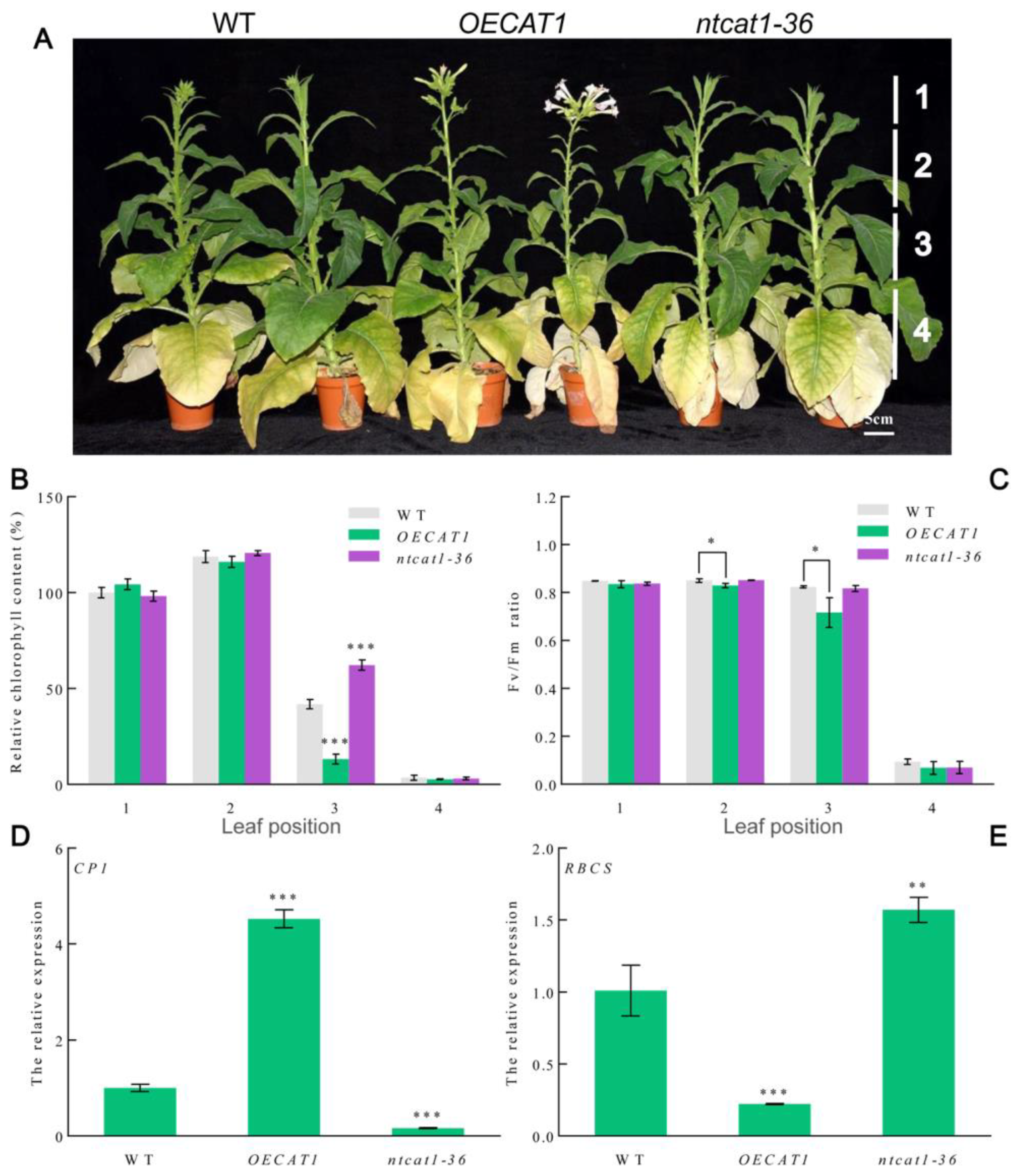
3.6. Overexpression of NtCAT1 Promotes Natural Leaf Senescence
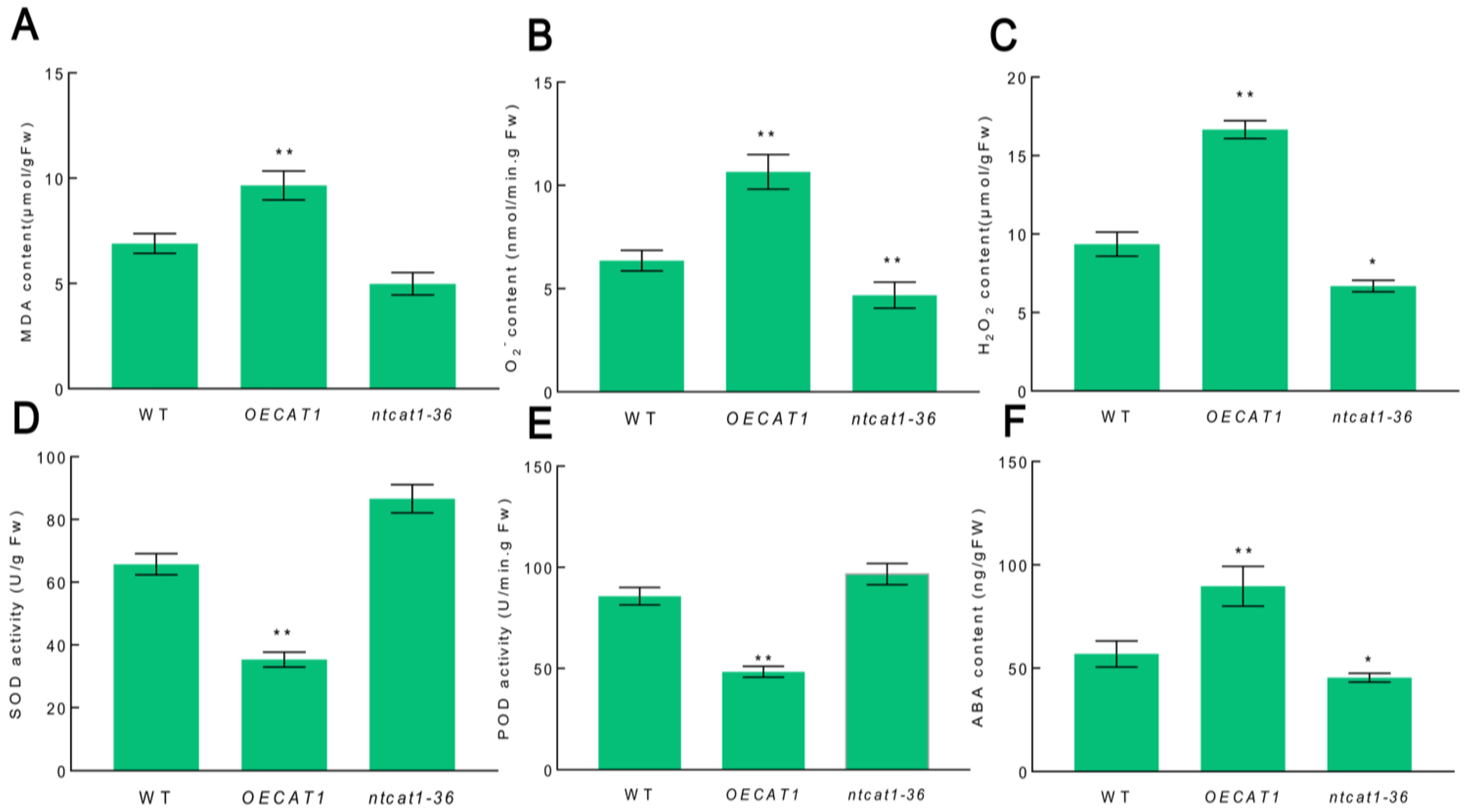
3.7. Overexpression of NtCAT1 Causes the Accumulation of Reactive Oxygen Species and ABA during Leaf Senescence
3.8. Altered Expression of NtCAT1 Influences the Expression of ABA-Associated Genes and TOR Gene during Leaf Senescence
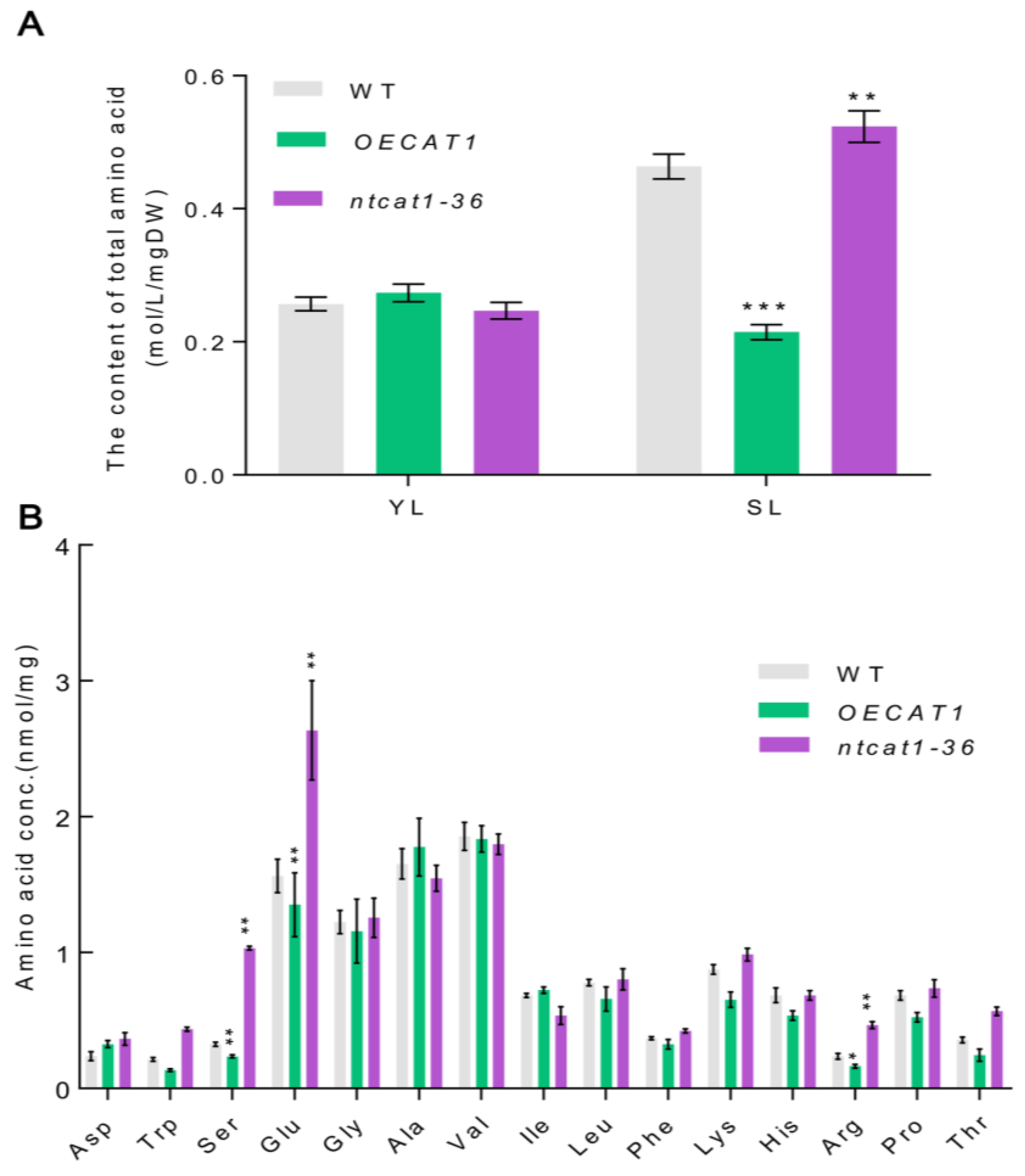
3.9. Overexpression of NtCAT1 Affects the Free Amino Acid Profile during Leaf Senescence
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Quirino, B.F.; Noh, Y.-S.; Himelblau, E.; Amasino, R.M. Molecular Aspects of Leaf Senescence. Trends Plant Sci. 2000, 5, 278–282. [Google Scholar] [CrossRef] [PubMed]
- Mao, C.; Lu, S.; Lv, B.; Zhang, B.; Shen, J.; He, J.; Luo, L.; Xi, D.; Chen, X.; Ming, F. A Rice Nac Transcription Factor Promotes Leaf Senescence via ABA Biosynthesis. Plant Physiol. 2017, 174, 1747–1763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, S.; Guo, Y. Signal Transduction in Leaf Senescence: Progress and Perspective. Plants 2019, 8, 405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Ren, G.; Zhang, K.; Li, Z.; Miao, Y.; Guo, H. Leaf Senescence: Progression, Regulation, and Application. Mol. Hortic. 2021, 1, 5. [Google Scholar] [CrossRef]
- Guo, Y.; Gan, S. Leaf Senescence: Signals, Execution, and Regulation. Curr. Top. Dev. Biol. 2005, 71, 83–112. [Google Scholar] [CrossRef]
- Li, Z.; Zhao, T.; Liu, J.; Li, H.; Liu, B. Shade-Induced Leaf Senescence in Plants. Plants 2023, 12, 1550. [Google Scholar] [CrossRef]
- Perchlik, M.; Tegeder, M. Improving Plant Nitrogen Use Efficiency through Alteration of Amino Acid Transport Processes. Plant Physiol. 2017, 175, 235–247. [Google Scholar] [CrossRef] [Green Version]
- Sanders, A.; Collier, R.; Trethewy, A.; Gould, G.; Sieker, R.; Tegeder, M. AAP1 Regulates Import of Amino Acids into Developing Arabidopsis Embryos. Plant J. 2009, 59, 540–552. [Google Scholar] [CrossRef]
- Perchlik, M.; Tegeder, M. Leaf Amino Acid Supply Affects Photosynthetic and Plant Nitrogen Use Efficiency under Nitrogen Stress. Plant Physiol. 2018, 178, 174–188. [Google Scholar] [CrossRef] [Green Version]
- Santiago, J.P.; Tegeder, M. Connecting Source with Sink: The Role of Arabidopsis AAP8 in Phloem Loading of Amino Acids. Plant Physiol. 2016, 171, 508–521. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Tan, Q.; Lee, R.; Trethewy, A.; Lee, Y.H.; Tegeder, M. Altered Xylem-Phloem Transfer of Amino Acids Affects Metabolism and Leads to Increased Seed Yield and Oil Content in Arabidopsis. Plant Cell 2010, 22, 3603–3620. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Postel, S.; Kemmerling, B.; Ludewig, U. Altered Growth and Improved Resistance of Arabidopsis against Pseudomonas Syringae by Overexpression of the Basic Amino Acid Transporter AtCAT1. Plant Cell Environ. 2014, 37, 1404–1414. [Google Scholar] [CrossRef]
- Lu, K.; Wu, B.; Wang, J.; Zhu, W.; Nie, H.; Qian, J.; Huang, W.; Fang, Z. Blocking Amino Acid Transporter OsAAP3 Improves Grain Yield by Promoting Outgrowth Buds and Increasing Tiller Number in Rice. Plant Biotechnol. J. 2018, 16, 1710–1722. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Wu, B.; Lu, K.; Wei, Q.; Qian, J.; Chen, Y.; Fang, Z. The Amino Acid Permease 5 (Osaap5) Regulates Tiller Number and Grain Yield in Rice. Plant Physiol. 2019, 180, 1031–1045. [Google Scholar] [CrossRef]
- Wei, Q.; Yan, Z.; Xiong, Y.; Fang, Z. Altered Expression of Osaap3 Influences Rice Lesion Mimic and Leaf Senescence by Regulating Arginine Transport and Nitric Oxide Pathway. Int. J. Mol. Sci. 2021, 22, 2181. [Google Scholar] [CrossRef]
- Yang, G.; Wei, X.; Fang, Z. Melatonin Mediates Axillary Bud Outgrowth by Improving Nitrogen Assimilation and Transport in Rice. Front. Plant Sci. 2022, 13, 900262. [Google Scholar] [CrossRef]
- Couturier, J.; Doidy, J.; Guinet, F.; Wipf, D.; Blaudez, D.; Chalot, M. Glutamine, Arginine and the Amino Acid Transporter Pt-CAT11 Play Important Roles during Senescence in Poplar. Ann. Bot. 2010, 105, 1159–1169. [Google Scholar] [CrossRef]
- Zhang, H.; Zhao, Y.; Zhu, J.K. Thriving under Stress: How Plants Balance Growth and the Stress Response. Dev. Cell 2020, 55, 529–543. [Google Scholar] [CrossRef]
- Zhang, H.; Zhu, J.; Gong, Z.; Zhu, J.-K. Abiotic Stress Responses in Plants. Nat. Rev. Genet. 2022, 23, 104–119. [Google Scholar] [CrossRef]
- Yang, J.; Worley, E.; Udvardi, M. A NAP-AAO3 Regulatory Module Promotes Chlorophyll Degradation via Aba Biosynthesis in Arabidopsis Leavesw Open. Plant Cell 2014, 26, 4862–4874. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Ma, S.; Ye, N.; Jiang, M.; Cao, J.; Zhang, J. WRKY Transcription Factors in Plant Responses to Stresses. J. Integr. Plant Biol. 2017, 59, 86–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rong, H.; Tang, Y.; Zhang, H.; Wu, P.; Chen, Y.; Li, M.; Wu, G.; Jiang, H. The Stay-Green Rice like (SGRL) Gene Regulates Chlorophyll Degradation in Rice. J. Plant Physiol. 2013, 170, 1367–1373. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Yu, J.W.; Park, J.S.; Li, J.; Yoo, S.C.; Lee, N.Y.; Lee, S.K.; Jeong, S.W.; Seo, H.S.; Koh, H.J.; et al. The Senescence-Induced Staygreen Protein Regulates Chlorophyll Degradation. Plant Cell 2007, 19, 1649–1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kusaba, M.; Ito, H.; Morita, R.; Iida, S.; Sato, Y.; Fujimoto, M.; Kawasaki, S.; Tanaka, R.; Hirochika, H.; Nishimura, M.; et al. Rice NON-YELLOW COLORING1 Is Involved in Light-Harvesting Complex II and Grana Degradation during Leaf Senescence. Plant Cell 2007, 19, 1362–1375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Xiang, S.; Chen, Y.; Li, D.; Yu, D. Arabidopsis WRKY45 Interacts with the DELLA Protein RGL1 to Positively Regulate Age-Triggered Leaf Senescence. Mol. Plant 2017, 10, 1174–1189. [Google Scholar] [CrossRef] [Green Version]
- Tan, B.C.; Joseph, L.M.; Deng, W.T.; Liu, L.; Li, Q.B.; Cline, K.; McCarty, D.R. Molecular Characterization of the Arabidopsis 9-Cis Epoxycarotenoid Dioxygenase Gene Family. Plant J. 2003, 35, 44–56. [Google Scholar] [CrossRef]
- Oliver, S.N.; Dennis, E.S.; Dolferus, R. ABA Regulates Apoplastic Sugar Transport and Is a Potential Signal for Cold-Induced Pollen Sterility in Rice. Plant Cell Physiol. 2007, 48, 1319–1330. [Google Scholar] [CrossRef] [Green Version]
- Fujii, H.; Zhu, J.K. Arabidopsis Mutant Deficient in 3 Abscisic Acid-Activated Protein Kinases Reveals Critical Roles in Growth, Reproduction, and Stress. Proc. Natl. Acad. Sci. USA 2009, 106, 8380–8385. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Chan, Z.; Gao, J.; Xing, L.; Cao, M.; Yu, C.; Hu, Y.; You, J.; Shi, H.; Zhu, Y.; et al. ABA Receptor PYL9 Promotes Drought Resistance and Leaf Senescence. Proc. Natl. Acad. Sci. USA 2016, 113, 1949–1954. [Google Scholar] [CrossRef] [Green Version]
- Fu, L.; Wang, P.; Xiong, Y. Target of Rapamycin Signaling in Plant Stress Responses. Plant Physiol. 2020, 182, 1613–1623. [Google Scholar] [CrossRef] [Green Version]
- Dobrenel, T.; Caldana, C.; Hanson, J.; Robaglia, C.; Vincentz, M.; Veit, B.; Meyer, C. TOR Signaling and Nutrient Sensing. Annu. Rev. Plant Biol. 2016, 67, 261–285. [Google Scholar] [CrossRef]
- González, A.; Hall, M.N. Nutrient Sensing and TOR Signaling in Yeast and Mammals. EMBO J. 2017, 36, 397–408. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Y.; McCormack, M.; Li, L.; Hall, Q.; Xiang, C.; Sheen, J. Glucose–TOR Signalling Reprograms the Transcriptome and Activates Meristems. Nature 2013, 496, 181–186. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Zhao, Y.; Li, Z.; Hsu, C.-C.; Liu, X.; Fu, L.; Hou, Y.-J.; Du, Y.; Xie, S.; Zhang, C.; et al. Reciprocal Regulation of the TOR Kinase and ABA Receptor Balances Plant Growth and Stress Response. Mol. Cell 2018, 69, 100–112.e6. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Duan, X.; Zhao, X.; Ding, W.; Wang, Y.; Xiong, Y. Diverse Nitrogen Signals Activate Convergent ROP2-TOR Signaling in Arabidopsis. Dev. Cell 2021, 56, 1283–1295.e5. [Google Scholar] [CrossRef]
- Horsch, R.B.; Fry, J.E.; Hoffmann, N.L.; Wallroth, M.; Eichholtz, D.; Rogers, S.G.; Fraley, R.T. A Simple and General Method for Transferring Genes into Plants. Science 1985, 227, 1229–1231. [Google Scholar] [CrossRef]
- Sun, Y.; Song, K.; Guo, M.; Wu, H.; Ji, X.; Hou, L.; Liu, X.; Lu, S. A NAC Transcription Factor from ‘Sea Rice 86′ Enhances Salt Tolerance by Promoting Hydrogen Sulfide Production in Rice Seedlings. Int. J. Mol. Sci. 2022, 23, 6435. [Google Scholar] [CrossRef]
- Chen, X.; Lu, S.; Wang, Y.; Zhang, X.; Lv, B.; Luo, L.; Xi, D.; Shen, J.; Ma, H.; Ming, F. OsNAC2 Encoding a NAC Transcription Factor That Affects Plant Height through Mediating the Gibberellic Acid Pathway in Rice. Plant J. 2015, 82, 302–314. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.; Wang, Z.; Gong, G.; Zhu, Y.; Ye, Q.; Lu, S.; Liu, X. Hydrogen Sulfide Alleviates Manganese Stress in Arabidopsis. Int. J. Mol. Sci. 2022, 23, 5046. [Google Scholar] [CrossRef]
- Daie, J.; Wyse, R. Adaptation of the Enzyme-Linked Immunosorbent Assay (ELISA) to the Quantitative Analysis of Abscisic Acid. Anal. Biochem. 1982, 119, 365–371. [Google Scholar] [CrossRef]
- Gu, L.; Jones, A.D.; Last, R.L. LC−MS/MS Assay for Protein Amino Acids and Metabolically Related Compounds for Large-Scale Screening of Metabolic Phenotypes. Anal. Chem. 2007, 79, 8067–8075. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Seo, P.J.; Lee, H.J.; Park, C.M. A NAC Transcription Factor NTL4 Promotes Reactive Oxygen Species Production during Drought-Induced Leaf Senescence in Arabidopsis. Plant J. 2012, 70, 831–844. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.; Liu, T.; Deng, Z.; Zhang, Z.; Wang, Q.; Wang, W.; Li, W.; Guo, Y. Characterization of NAC Transcription Factor NtNAC028 as a Regulator of Leaf Senescence and Stress Responses. Front. Plant Sci. 2022, 13, 941026. [Google Scholar] [CrossRef] [PubMed]
- Pyung, O.L.; Hyo, J.K.; Hong, G.N. Leaf Senescence. Annu. Rev. Plant Biol. 2007, 58, 115–136. [Google Scholar] [CrossRef] [Green Version]
- Saha, J.; Gupta, S. Molecular Characterization, Evolutionary and Phylogenetic Analyses of Rice ACT/BAT-Type Amino Acid Transporters. Comput. Biol. Chem. 2022, 100, 107745. [Google Scholar] [CrossRef] [PubMed]
- Breeze, E.; Harrison, E.; McHattie, S.; Hughes, L.; Hickman, R.; Hill, C.; Kiddle, S.; Kim, Y.S.; Penfold, C.A.; Jenkins, D.; et al. High-Resolution Temporal Profiling of Transcripts during Arabidopsis Leaf Senescence Reveals a Distinct Chronology of Processes and Regulation. Plant Cell 2011, 23, 873–894. [Google Scholar] [CrossRef] [Green Version]
- Havé, M.; Marmagne, A.; Chardon, F.; Masclaux-Daubresse, C. Nitrogen Remobilization during Leaf Senescence: Lessons from Arabidopsis to Crops. J. Exp. Bot. 2017, 68, 2513–2529. [Google Scholar] [CrossRef]
- Palatnik, J.F.; Carrillo, N.; Valle, E.M. The Role of Photosynthetic Electron Transport in the Oxidative Degradation of Chloroplastic Glutamine Synthetase. Plant Physiol. 1999, 121, 471–478. [Google Scholar] [CrossRef] [Green Version]
- Kunkowska, A.B.; Fontana, F.; Betti, F.; Soeur, R.; Beckers, G.J.M.; Meyer, C.; De Jaeger, G.; Weits, D.A.; Loreti, E.; Perata, P. Target of Rapamycin Signaling Couples Energy to Oxygen Sensing to Modulate Hypoxic Gene Expression in Arabidopsis. Proc. Natl. Acad. Sci. USA 2023, 120, e2212474120. [Google Scholar] [CrossRef]
- Punzo, P.; Ruggiero, A.; Grillo, S.; Batelli, G. TIP41 Network Analysis and Mutant Phenotypes Predict Interactions between the TOR and ABA Pathways. Plant Signal. Behav. 2018, 13, e1537698. [Google Scholar] [CrossRef]
- Belda-Palazón, B.; Costa, M.; Beeckman, T.; Rolland, F.; Baena-González, E. ABA Represses TOR and Root Meristem Activity through Nuclear Exit of the SnRK1 Kinase. Proc. Natl. Acad. Sci. USA 2022, 119, e2204862119. [Google Scholar] [CrossRef]
- Ingargiola, C.; Jéhanno, I.; Forzani, C.; Marmagne, A.; Broutin, J.; Clément, G.; Leprince, A.-S.; Meyer, C. The Arabidopsis Target of Rapamycin Kinase Regulates Ammonium Assimilation and Glutamine Metabolism. Plant Physiol. 2023, kiad216. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, S.; Ji, X.; Zhang, X.; Wu, H.; Sun, Y.; Zhu, Y.; Su, S.; Wei, S.; Liu, X. A Cationic Amino Acid Transporter NtCAT1 Promotes Leaf Senescence by the Accumulation of ABA in Nicotiana tabacum. Agronomy 2023, 13, 1691. https://doi.org/10.3390/agronomy13071691
Lu S, Ji X, Zhang X, Wu H, Sun Y, Zhu Y, Su S, Wei S, Liu X. A Cationic Amino Acid Transporter NtCAT1 Promotes Leaf Senescence by the Accumulation of ABA in Nicotiana tabacum. Agronomy. 2023; 13(7):1691. https://doi.org/10.3390/agronomy13071691
Chicago/Turabian StyleLu, Songchong, Xuan Ji, Xinshuang Zhang, Hao Wu, Yan Sun, Ying Zhu, Shanshan Su, Shumin Wei, and Xin Liu. 2023. "A Cationic Amino Acid Transporter NtCAT1 Promotes Leaf Senescence by the Accumulation of ABA in Nicotiana tabacum" Agronomy 13, no. 7: 1691. https://doi.org/10.3390/agronomy13071691
APA StyleLu, S., Ji, X., Zhang, X., Wu, H., Sun, Y., Zhu, Y., Su, S., Wei, S., & Liu, X. (2023). A Cationic Amino Acid Transporter NtCAT1 Promotes Leaf Senescence by the Accumulation of ABA in Nicotiana tabacum. Agronomy, 13(7), 1691. https://doi.org/10.3390/agronomy13071691