
Construction and Functional Evaluation of CRISPR/Cas9 Multiple Knockout Vectors of the FAD2 Gene Family


Abstract
:1. Introduction
2. Results
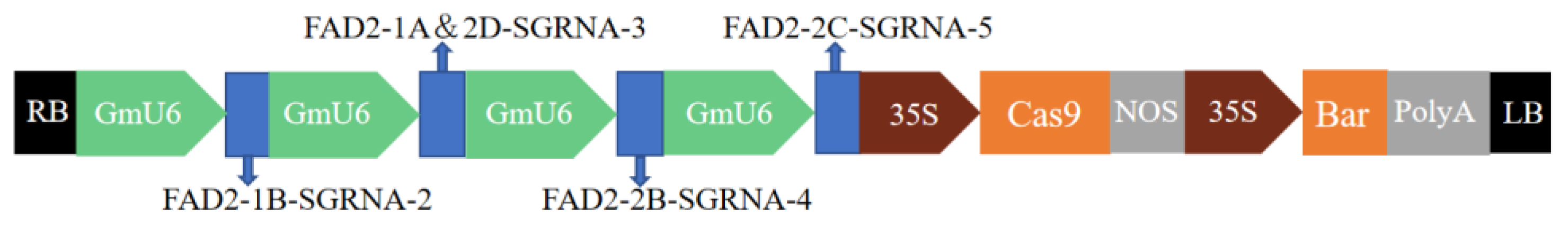
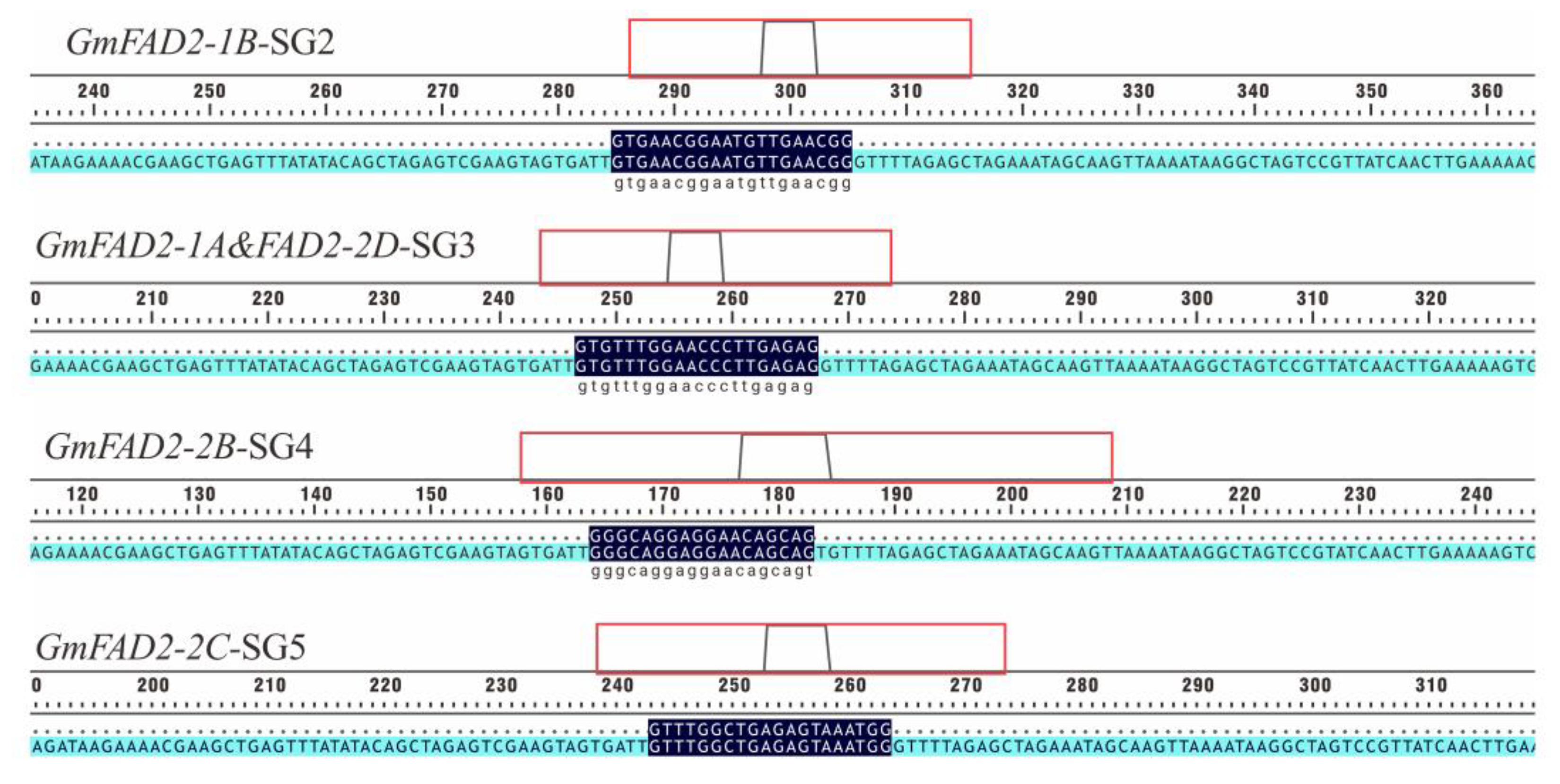
2.1. Construction and Validation of CRISPR/Cas9 Multiplex Knockout Expression Vector
2.2. Genetic Transformation and Identification of Vectors
2.2.1. Agrobacterium-Mediated Soybean Cotyledonary Nodes
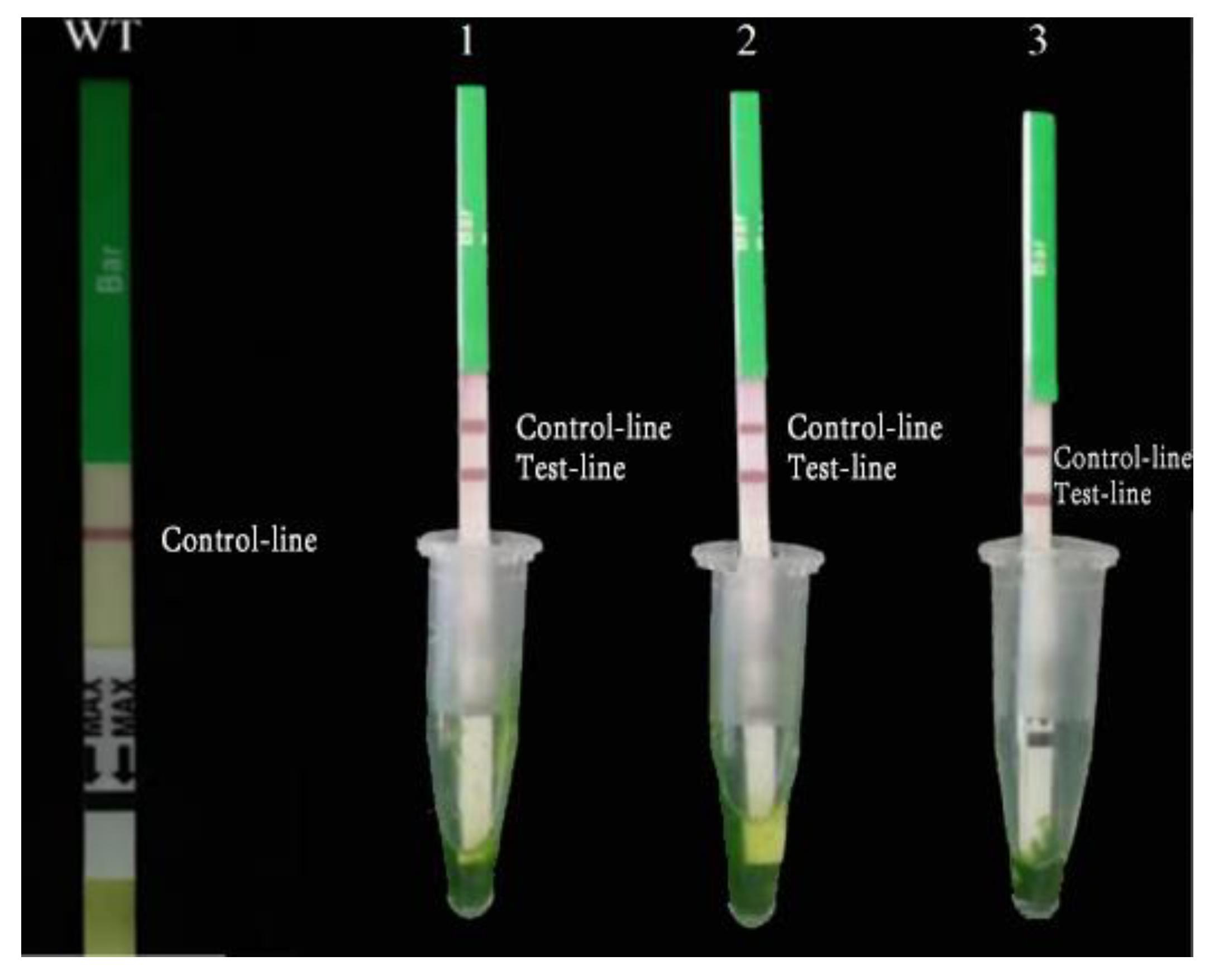
2.2.2. Detection of Positive Soybean
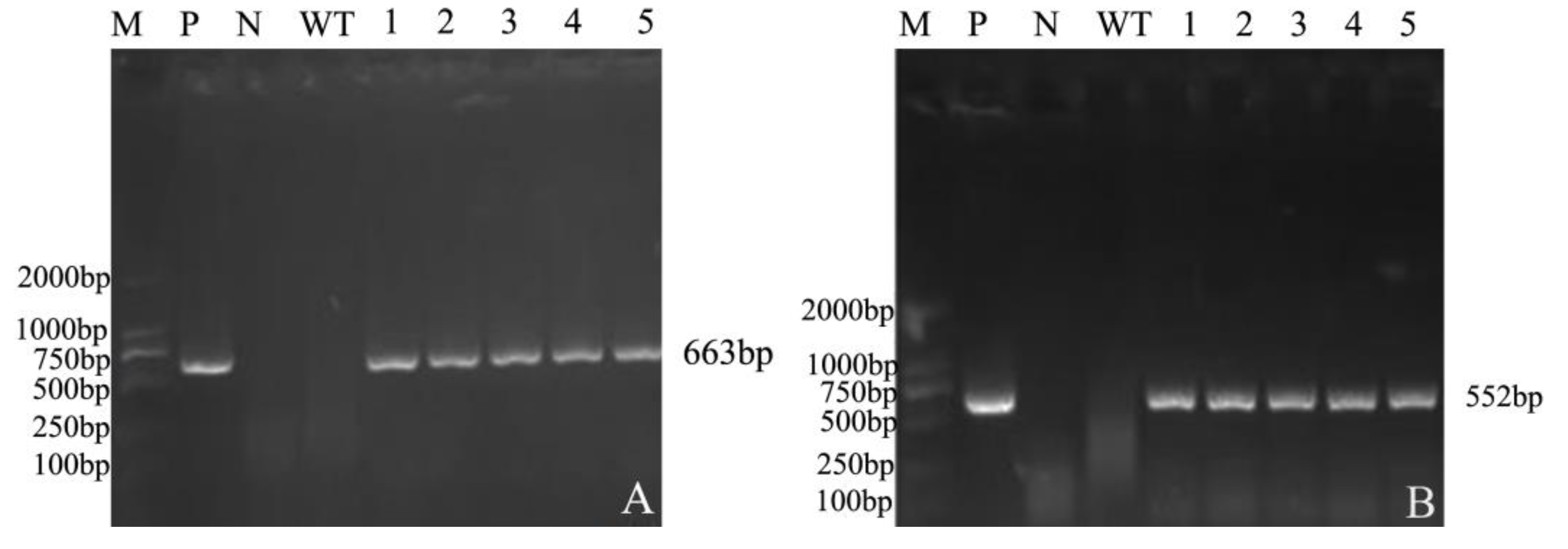
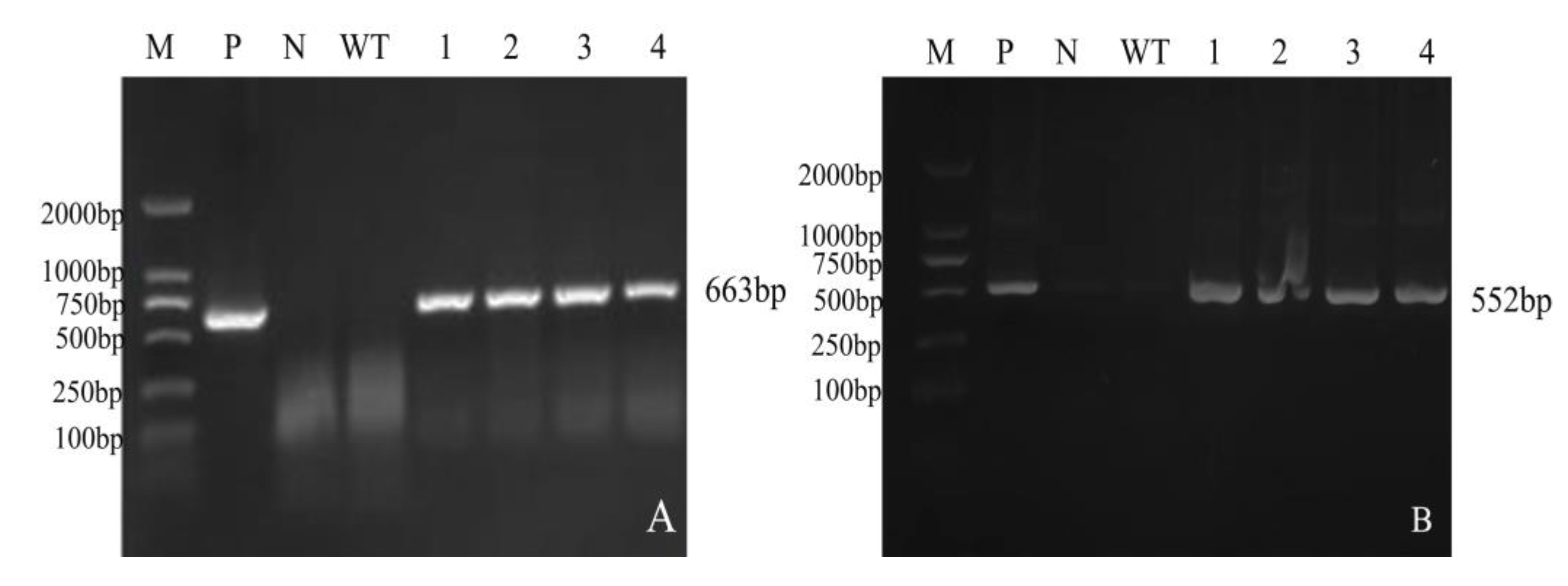
2.2.3. Detection of T1 Generation Mutant Plants of CRISPR/Cas9 Multiple Knockout Vector
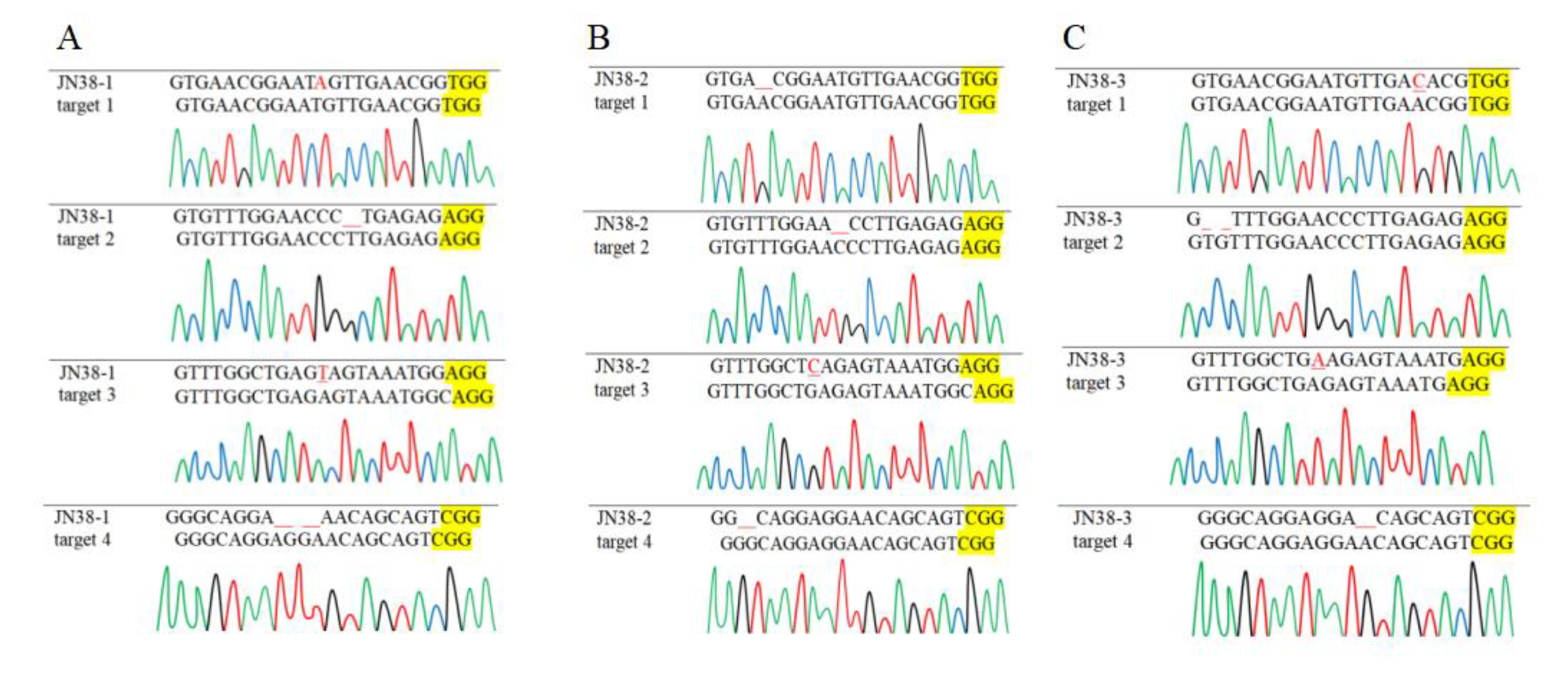
2.2.4. PCR Detection of Mutant Plants
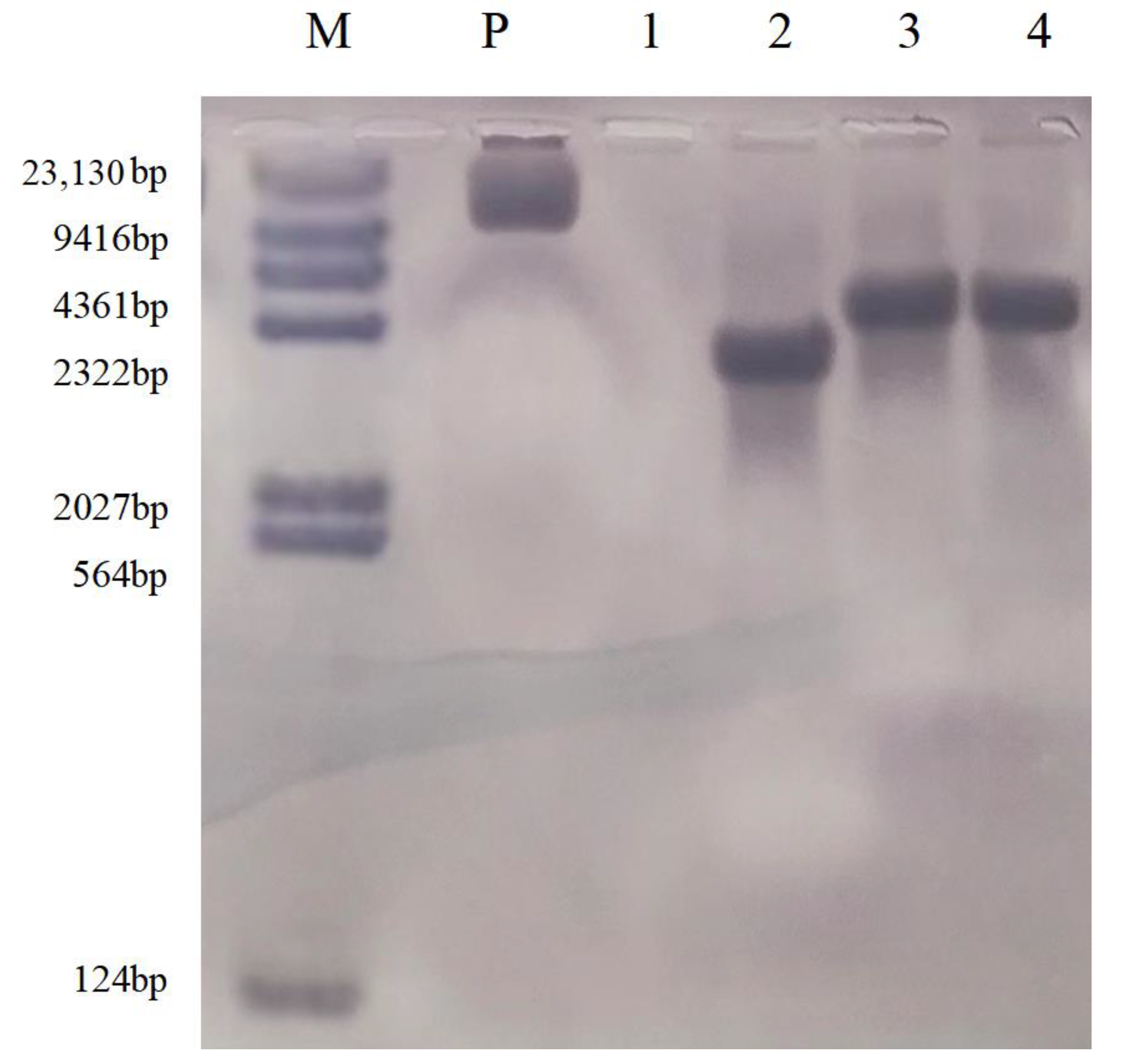
2.2.5. Southern Blot Analysis of T2 Transgenic Plants
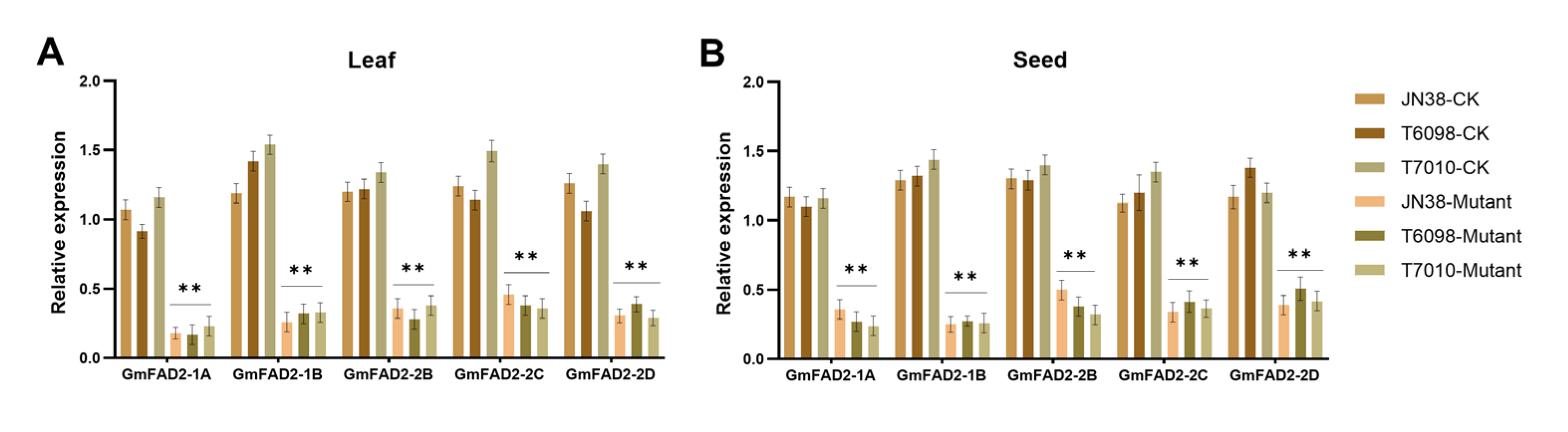
2.3. RT-qPCR Assay of Gene-Edited Progeny
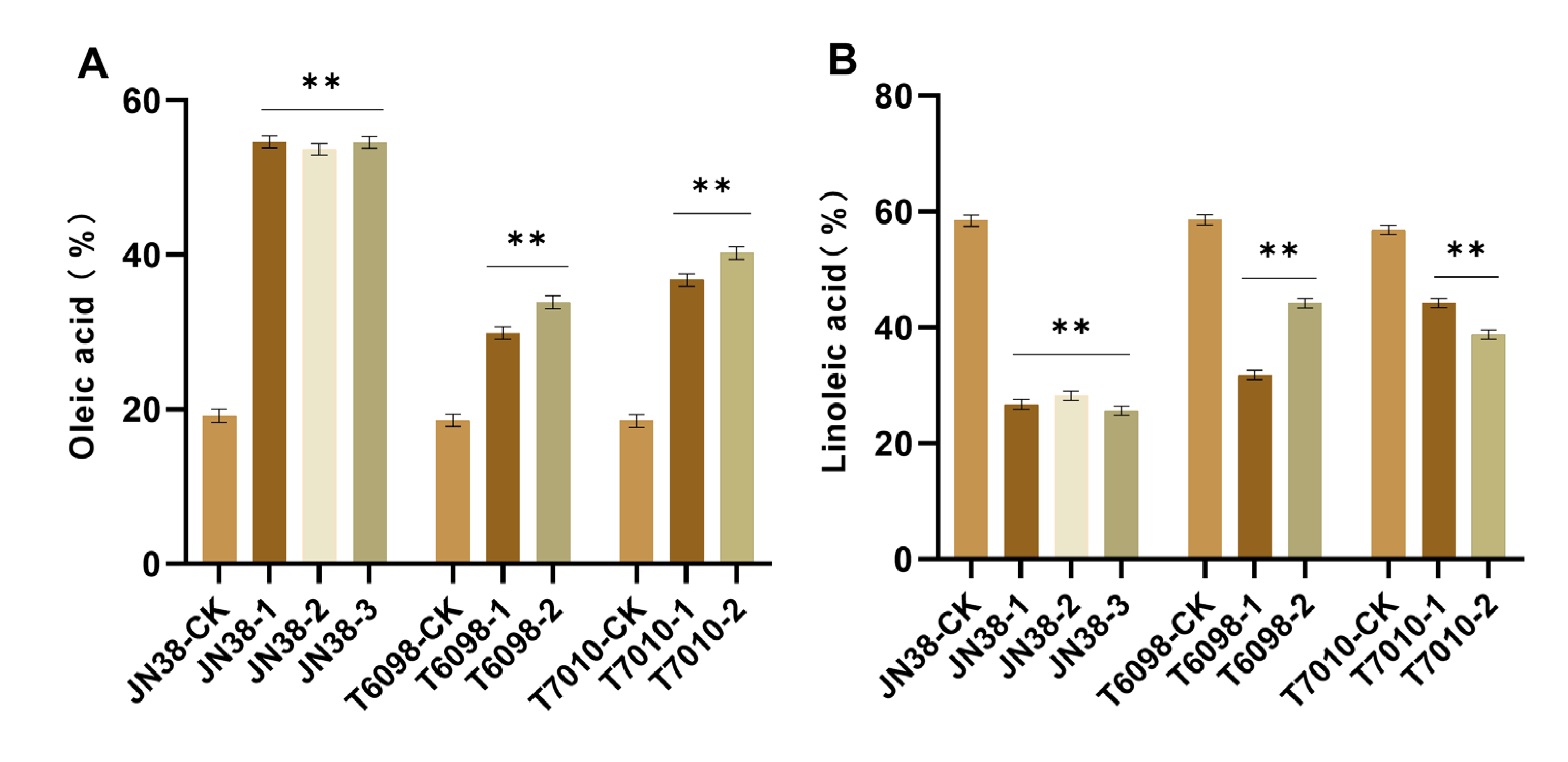
2.4. Determination of Fatty Acid Content of Different Editing Vectors in T2 Plants
2.5. Comparison of Agronomic Traits
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Construction of the CRISPR/Cas9 Vector
4.3. Genetic Transformation
4.4. Bar-Positive Plant Detection Test Strip
4.5. Evaluation of Transformation Efficiency and Gene-Editing Efficiency
4.6. Southern Blot Analysis of the T2 Transgenic Plants
4.7. Real-Time Quantitative PCR (RT-qPCR)
4.8. Determination of Main Oil Content
4.9. Analysis of the Main Agronomic Traits
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ren, D.; Yang, H.; Zhou, L.; Yang, Y.; Liu, W.; Hao, X.; Pan, P. The Land-Water-Food-Environment Nexus in the Context of China’s Soybean Import. Adv. Water Resour. 2021, 151, 103892. [Google Scholar] [CrossRef]
- Ghassemi-Golezani, K.; Farhangi-Abriz, S. Changes in Oil Accumulation and Fatty Acid Composition of Soybean Seeds under Salt Stress in Response to Salicylic Acid and Jasmonic Acid. Russ. J. Plant Physiol. 2018, 65, 229–236. [Google Scholar] [CrossRef]
- Gangadhara, K.; Nadaf, H.L. Genetic Analysis of Oleic Acid and Linoleic Acid Content in Relation to Oil Quality in Groundnut. Electron. J. Plant Breed. 2018, 9, 283. [Google Scholar] [CrossRef]
- Jones, P.J.H.; Jew, S.; AbuMweis, S. The Effect of Dietary Oleic, Linoleic, and Linolenic Acids on Fat Oxidation and Energy Expenditure in Healthy Men. Metabolism 2008, 57, 1198–1203. [Google Scholar] [CrossRef] [PubMed]
- Demorest, Z.L.; Coffman, A.; Baltes, N.J.; Stoddard, T.J.; Clasen, B.M.; Luo, S.; Retterath, A.; Yabandith, A.; Gamo, M.E.; Bissen, J.; et al. Direct Stacking of Sequence-Specific Nuclease-Induced Mutations to Produce High Oleic and Low Linolenic Soybean Oil. BMC Plant Biol. 2016, 16, 225. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.-S.; Liu, T.-T. Optimization of Production of Conjugated Linoleic Acid from Soybean Oil. J. Agric. Food Chem. 2004, 52, 5079–5084. [Google Scholar] [CrossRef] [PubMed]
- Elagizi, A.; Lavie, C.J.; O’Keefe, E.; Marshall, K.; O’Keefe, J.H.; Milani, R.V. An Update on Omega-3 Polyunsaturated Fatty Acids and Cardiovascular Health. Nutrients 2021, 13, 204. [Google Scholar] [CrossRef] [PubMed]
- Otyama, P.I.; Chamberlin, K.; Ozias-Akins, P.; Graham, M.A.; Cannon, E.K.S.; Cannon, S.B.; MacDonald, G.E.; Anglin, N.L. Genome-Wide Approaches Delineate the Additive, Epistatic, and Pleiotropic Nature of Variants Controlling Fatty Acid Composition in Peanut (Arachis hypogaea L.). G3 Genes Genomes Genet. 2022, 12, jkab382. [Google Scholar] [CrossRef]
- Monteros, M.J.; Burton, J.W.; Boerma, H.R. Molecular Mapping and Confirmation of QTLs Associated with Oleic Acid Content in N00-3350 Soybean. Crop. Sci. 2008, 48, 2223–2234. [Google Scholar] [CrossRef]
- Dar, A.A.; Choudhury, A.R.; Kancharla, P.K.; Arumugam, N. The FAD2 Gene in Plants: Occurrence, Regulation, and Role. Front. Plant Sci. 2017, 8, 1789. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.-H.; Li, Q.-Q.; Wang, T.-T.; Hu, Q.; Deng, W.-H.; Xia, X.-L.; Gao, H.-B. XsFAD2 Gene Encodes the Enzyme Responsible for the High Linoleic Acid Content in Oil Accumulated in Xanthoceras sorbifolia Seeds: XsFAD2 Is Responsible for the High Linoleic Acid Content in X. sorbifolia Seeds. J. Sci. Food Agric. 2014, 94, 482–488. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Ruan, J.; Tian, H.; Shan, L.; Meng, J.; Guo, F.; Zhang, Z.; Ding, H.; Wan, S.; Li, X. The Family of Peanut Fatty Acid Desaturase Genes and a Functional Analysis of Four ω-3 AhFAD3 Members. Plant Mol. Biol. Rep. 2020, 38, 209–221. [Google Scholar] [CrossRef] [Green Version]
- Heppard, E.P.; Kinney, A.J.; Stecca, K.L.; Miao, G.H. Developmental and Growth Temperature Regulation of Two Different Microsomal [Omega]-6 Desaturase Genes in Soybeans. Plant Physiol. 1996, 110, 311–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.-R.; Jeon, I.; Yu, H.; Kim, S.-G.; Kim, H.-S.; Ahn, S.-J.; Lee, J.; Lee, S.-K.; Kim, H.U. Increasing Monounsaturated Fatty Acid Contents in Hexaploid Camelina Sativa Seed Oil by FAD2 Gene Knockout Using CRISPR-Cas9. Front. Plant Sci. 2021, 12, 702930. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yang, X.; Zhang, Y.; Yang, J.; Qi, G.; Guo, D.; Xing, G.; Yao, Y.; Xu, W.; Li, H.; et al. Changes in Oleic Acid Content of Transgenic Soybeans by Antisense RNA Mediated Posttranscriptional Gene Silencing. Int. J. Genom. 2014, 2014, 92195. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Z.; Jin, Y.; Zhang, Q.; Lamboro, A.; Dong, B.; Yang, Z.; Wang, P. Construction and Functional Analysis of CRISPR/Cas9 Vector of FAD2 Gene Family in Soybean. Phyton 2022, 91, 349–361. [Google Scholar] [CrossRef]
- Bao, A.; Chen, H.; Chen, L.; Chen, S.; Hao, Q.; Guo, W.; Qiu, D.; Shan, Z.; Yang, Z.; Yuan, S.; et al. CRISPR/Cas9-Mediated Targeted Mutagenesis of GmSPL9 Genes Alters Plant Architecture in Soybean. BMC Plant Biol. 2019, 19, 131. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Nguyen, V.; Liu, J.; Fu, W.; Chen, C.; Yu, K.; Cui, Y. Mutagenesis of Seed Storage Protein Genes in Soybean Using CRISPR/Cas9. BMC Res. Notes 2019, 12, 176. [Google Scholar] [CrossRef] [Green Version]
- Wu, N.; Lu, Q.; Wang, P.; Zhang, Q.; Zhang, J.; Qu, J.; Wang, N. Construction and Analysis of GmFAD2-1A and GmFAD2-2A Soybean Fatty Acid Desaturase Mutants Based on CRISPR/Cas9 Technology. Int. J. Mol. Sci. 2020, 21, 1104. [Google Scholar] [CrossRef] [Green Version]
- Neelakandan, A.K.; Wright, D.A.; Traore, S.M.; Chen, X.; Spalding, M.H.; He, G. CRISPR/Cas9 Based Site-Specific Modification of FAD2 Cis-Regulatory Motifs in Peanut (Arachis hypogaea L). Front. Genet. 2022, 13, 849961. [Google Scholar] [CrossRef]
- Ben Ayed, R.; Chirmade, T.; Hanana, M.; Khamassi, K.; Ercisli, S.; Choudhary, R.; Kadoo, N.; Karunakaran, R. Comparative Analysis and Structural Modeling of Elaeis Oleifera FAD2, a Fatty Acid Desaturase Involved in Unsaturated Fatty Acid Composition of American Oil Palm. Biology 2022, 11, 529. [Google Scholar] [CrossRef] [PubMed]
- Turquetti-Moraes, D.K.; Moharana, K.C.; Almeida-Silva, F.; Pedrosa-Silva, F.; Venancio, T.M. Integrating Omics Approaches to Discover and Prioritize Candidate Genes Involved in Oil Biosynthesis in Soybean. Gene 2022, 808, 145976. [Google Scholar] [CrossRef]
- Dehghan Nayeri, F.; Yarizade, K. Bioinformatics Study of Delta-12 Fatty Acid Desaturase 2 (FAD2) Gene in Oilseeds. Mol. Biol. Rep. 2014, 41, 5077–5087. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wang, X.; Gai, J.; Yu, D. Isolation and Characterization of a Seed-Specific Isoform of Microsomal Omega-6 Fatty Acid Desaturase Gene (FAD2-1B) from Soybean. DNA Seq. 2008, 19, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Gaskin, E.L.; Carrero-Colón, M.; Hudson, K.A. Combination of the Elevated Stearic Acid Trait with Other Fatty Acid Traits in Soybean. J. Am. Oil Chem. Soc. 2021, 98, 221–226. [Google Scholar] [CrossRef]
- Ma, J.; Sun, S.; Whelan, J.; Shou, H. CRISPR/Cas9-Mediated Knockout of GmFATB1 Significantly Reduced the Amount of Saturated Fatty Acids in Soybean Seeds. Int. J. Mol. Sci. 2021, 22, 3877. [Google Scholar] [CrossRef]
- Lakhssassi, N.; Zhou, Z.; Liu, S.; Colantonio, V.; AbuGhazaleh, A.; Meksem, K. Characterization of the FAD2 Gene Family in Soybean Reveals the Limitations of Gel-Based TILLING in Genes with High Copy Number. Front. Plant Sci. 2017, 8, 324. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Song, K.; Cai, Y.; Li, W.; Liu, B.; Liu, L. Genetic Modification of Soybean with a Novel Grafting Technique: Downregulating the FAD2-1 Gene Increases Oleic Acid Content. Plant Mol. Biol. Rep. 2011, 29, 866–874. [Google Scholar] [CrossRef]
- Jo, H.; Woo, C.; Norah, N.; Song, J.T.; Lee, J.-D. Novel Allele of FAD2-1A from an EMS-Induced Mutant Soybean Line (PE529) Produces Elevated Levels of Oleic Acid in Soybean Oil. Agronomy 2022, 12, 2115. [Google Scholar] [CrossRef]
- Cao, S.; Zhang, J.; Cheng, H.; Aslam, M.; Lv, H.; Dong, W.; Hu, A.; Guo, M.; Liu, Q.; Qin, Y. Identification and Evolutionary Analysis of FAD2 Gene Family in Green Plants. Trop. Plant Biol. 2021, 14, 239–250. [Google Scholar] [CrossRef]
- Song, Y.; Zhang, X.; Wang, P.; Liu, Z.; Dong, H.; Ma, J.; Fu, Y.; Zhang, Z. Characterization of Transformed Soybean Strains Expressing GbNPR1 and HrpZpsg12 Genes for Disease Resistance. Euphytica 2016, 211, 369–377. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Target Sequence + PAM | SG Sequence |
|---|---|---|
| GmFAD2-1B | ATGAACGGAATGTTGAACGGTGG | GTGAACGGAATGTTGAACGG |
| GmFAD21A&2D | GTGTTTGGAACCCTTGAGAGAGG | GTGTTTGGAACCCTTGAGAG |
| GmFAD2-2B | ATTTGGCTGAGAGTAAATGGAGG | GTTTGGCTGAGAGTAAATGG |
| GmFAD2-2C | TGGCAGGAGGAACAGCAGTTCGG | GGGCAGGAGGAACAGCAGT |
| Plant Genotype | Target Spot | Target Sequence | Mutation Type |
|---|---|---|---|
| WT | target 1 | GTGAACGGAATGTTGAACGGTGG | |
| target 2 | GTGTTTGGAACCCTTGAGAGAGG | ||
| target 3 | GTTTGGCTGAGAGTAAATGGCAGG | ||
| target 4 | GGGCAGGAGGAACAGCAGTCGG | ||
| JN38-FAD2-1A-1B-2B-2C-2D -mutant-1 | target 1 | GTGAACGGAATAGTTGAACGGTGG | Insertion |
| target 2 | GTGTTTGGAACCC TGAGAGAGG | Deletion | |
| target 3 | GTTTGGCTGAGTAGTAAATGGAGG | Insertion | |
| target 4 | GGGCAGGA AACAGCAGTCGG | Deletion | |
| JN38-FAD2-1A-1B-2B-2C-2D -mutant-2 | target 1 | GTGA CGGAATGTTGAACGGTGG | Deletion |
| target 2 | GTGTTTGGAA CCTTGAGAGAGG | Deletion | |
| target 3 | GTTTGGCTCAGAGTAAATGGAGG | Substitution | |
| target 4 | GG CAGGAGGAACAGCAGTCGG | Deletion | |
| JN38-FAD2-1A-1B-2B-2C-2D -mutant-3 | target 1 | GTGAACGGAATGTTGACACGTGG | Insertion |
| target 2 | G TTTGGAACCCTTGAGAGAGG | Deletion | |
| target 3 | GTTTGGCTGAAGAGTAAATGAGG | Insertion | |
| target 4 | GGGCAGGAGGA CAGCAGTCGG | Deletion | |
| T6098-FAD2-1A-1B-1B-2C-2D -mutant-1 | target 1 | GTGAACGGAGATGTTGAAGGTGG | Insertion |
| target 2 | GTGTTAGGAACCCTTGAGAGAGG | Substitution | |
| target 3 | GTTTGGCTGAGAGTAAA GGAGG | Deletion | |
| target 4 | GGGC GGAGGAACAGCAGTCGG | Deletion | |
| T6098-FAD2-1A-1B-1B-2C-2D -mutant-2 | target 1 | GTGAACGGACTATGTTGAACGTGG | Insertion |
| target 2 | GTGTTTGGAACCCTT AGAGAGG | Deletion | |
| target 3 | GTTTGGCTGAGAG AAATGGAGG | Deletion | |
| target 4 | GGGCGAGGAGGAACAGCAGTCGG | Insertion | |
| T7010-FAD2-1A-1B-1B-2C-2D -mutant-1 | Target 1 | GTGAACGTAATGTTGAACGGTGG | Substitution |
| Target 2 | GTGTCTTGGAACCCTTGAGAGAGG | Insertion | |
| Target 3 | GTTT GCTGAGAGTAAATGGAGG | Deletion | |
| Target 4 | GGGCAGGAGGAACAG AGTCGG | Deletion | |
| T7010-FAD2-1A-1B-1B-2C-2D -mutant-1 | Target 1 | GTGAACGG TGTTGAACGGTGG | Deletion |
| Target 2 | GTGTTGTGGAACCCTTGAGAGAGG | Insertion | |
| Target 3 | GTTTGGCTGAGAGT AATGGAGG | Deletion | |
| Target 4 | GGGCAGGAGGAAC GCAGTCGG | Deletion |
| Cultivar | Palmitic Acid (%) | Stearic Acid (%) | Oleic Acid (%) | Linoleic Acid (%) | Linolenic Acid (%) | Fat Content (%) |
|---|---|---|---|---|---|---|
| JN38 (CK) | 10.23 ± 0.11 a | 3.67 ± 0.06 b | 18.58 ± 0.23 b | 57.79 ± 0.32 a | 9.73 ± 0.19 a | 21.52 ± 0.12 a |
| JN38-FAD2-1A-1B-2B-2C-2D-mutant-1 | 7.69 ± 0.08 b | 4.40 ± 0.13 a | 54.07 ± 0.15 a | 26.17 ± 0.17 b | 7.67 ± 0.13 b | 21.79 ± 0.11 b |
| JN38-FAD2-1A-1B-2B-2C-2D-mutant-2 | 6.39 ± 0.07 b | 4.74 ± 0.05 b | 53.09 ± 0.12 a | 27.61 ± 0.19 a | 7.82 ± 0.12 a | 22.01 ± 0.12 b |
| JN38-FAD2-1A-1B-2B-2C-2D-mutant-3 | 8.02 ± 0.11 a | 4.13 ± 0.07 a | 54.01 ± 0.14 a | 25.09 ± 0.12 a | 8.13 ± 0.11 b | 21.93 ± 0.13 b |
| T6098 (CK) | 9.87 ± 0.17 a | 2.69 ± 0.12 b | 18.00 ± 0.13 b | 57.98 ± 0.21 a | 11.46 ± 0.09 a | 19.68 ± 0.12 a |
| T6098-FAD2-1A-2B-1B-2C-2D-mutant-1 | 8.31 ± 0.12 a | 2.46 ± 0.08 a | 29.31 ± 0.12 a | 31.24 ± 0.12 a | 9.71 ± 0.12 a | 19.75 ± 0.11 b |
| T6098-FAD2-1A-2B-1B-2C-2D-mutant-2 | 7.94 ± 0.21 b | 6.44 ± 0.18 a | 33.21 ± 0.24 a | 43.57 ± 0.15 b | 8.84 ± 0.17 b | 20.02 ± 0.13 b |
| T7010 (CK) | 11.05 ± 0.15 a | 3.31 ± 0.07 b | 17.92 ± 0.16 b | 56.26 ± 0.14 a | 11.46 ± 0.12 a | 18.95 ± 0.12 a |
| T7010-FAD2-1A-2B-1B-2C-2D-mutant-1 | 6.77 ± 0.08 b | 5.12 ± 0.14 a | 36.21 ± 0.11 a | 43.61 ± 0.11 b | 8.29 ± 0.11 b | 19.01 ± 0.13 |
| T7010-FAD2-1A-2B-1B-2C-2D-mutant-2 | 7.76 ± 0.14 b | 4.49 ± 0.14 a | 39.65 ± 0.13 a | 38.21 ± 0.13 a | 9.04 ± 0.13 a | 19.13 ± 0.11 b |
| Agronomic Trait | JN38 WT) | JN38-FAD2-1A-1B-2B-2C-2D -mutant | T6098 (WT) | T6098-FAD2-1A-1B-1B-2C-2D -mutant | T7010 (WT) | T7010-FAD2-1A-1B-1B-2C-2D -mutant |
| Plant height (cm) | 90.33 ± 4.03 b | 108.67 ± 4.68 a | 104.67 ± 3.56 a | 104.83 ± 3.43 a | 108.83 ± 4.36 a | 109.33 ± 4.08 a |
| Main stem nodes | 14 ± 2 a | 14 ± 2 a | 16 ± 2 a | 16 ± 2 a | 16 ± 2 a | 16 ± 2 a |
| Number of single branches | 2 ± 1 a | 2 ± 1 a | 2 ± 1 a | 2 ± 1 a | 3 ± 1 a | 3 ± 1 a |
| Leaf shape | roundleaf | roundleaf | lobus apicalis | lobus apicalis | lobus apicalis | lobus apicalis |
| Flower color | purple | purple | purple | purple | white | white |
| Grain color | yellow | yellow | black | black | yellow | yellow |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Q.; Liu, L.; Xiao, Z.; Sun, Y.; Xi, Y.; Sun, T.; Wang, J.; Wang, P. Construction and Functional Evaluation of CRISPR/Cas9 Multiple Knockout Vectors of the FAD2 Gene Family. Agronomy 2023, 13, 1737. https://doi.org/10.3390/agronomy13071737
Zhang Q, Liu L, Xiao Z, Sun Y, Xi Y, Sun T, Wang J, Wang P. Construction and Functional Evaluation of CRISPR/Cas9 Multiple Knockout Vectors of the FAD2 Gene Family. Agronomy. 2023; 13(7):1737. https://doi.org/10.3390/agronomy13071737
Chicago/Turabian StyleZhang, Qi, Lu Liu, Zhifeng Xiao, Yuwei Sun, Yongjing Xi, Tingting Sun, Jiabao Wang, and Piwu Wang. 2023. "Construction and Functional Evaluation of CRISPR/Cas9 Multiple Knockout Vectors of the FAD2 Gene Family" Agronomy 13, no. 7: 1737. https://doi.org/10.3390/agronomy13071737
APA StyleZhang, Q., Liu, L., Xiao, Z., Sun, Y., Xi, Y., Sun, T., Wang, J., & Wang, P. (2023). Construction and Functional Evaluation of CRISPR/Cas9 Multiple Knockout Vectors of the FAD2 Gene Family. Agronomy, 13(7), 1737. https://doi.org/10.3390/agronomy13071737