
Contribution of Putrescine and Glutamic Acid on γ-Aminobutyric Acid Accumulation of Malus baccata Borkh. Roots under Suboptimal Low Root-Zone Temperature

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials, Culture Conditions and Treatments
2.2. Determination of γ-Aminobutyric Acid and Glutamic Acid Content
2.3. Determination of Polyamines Contents
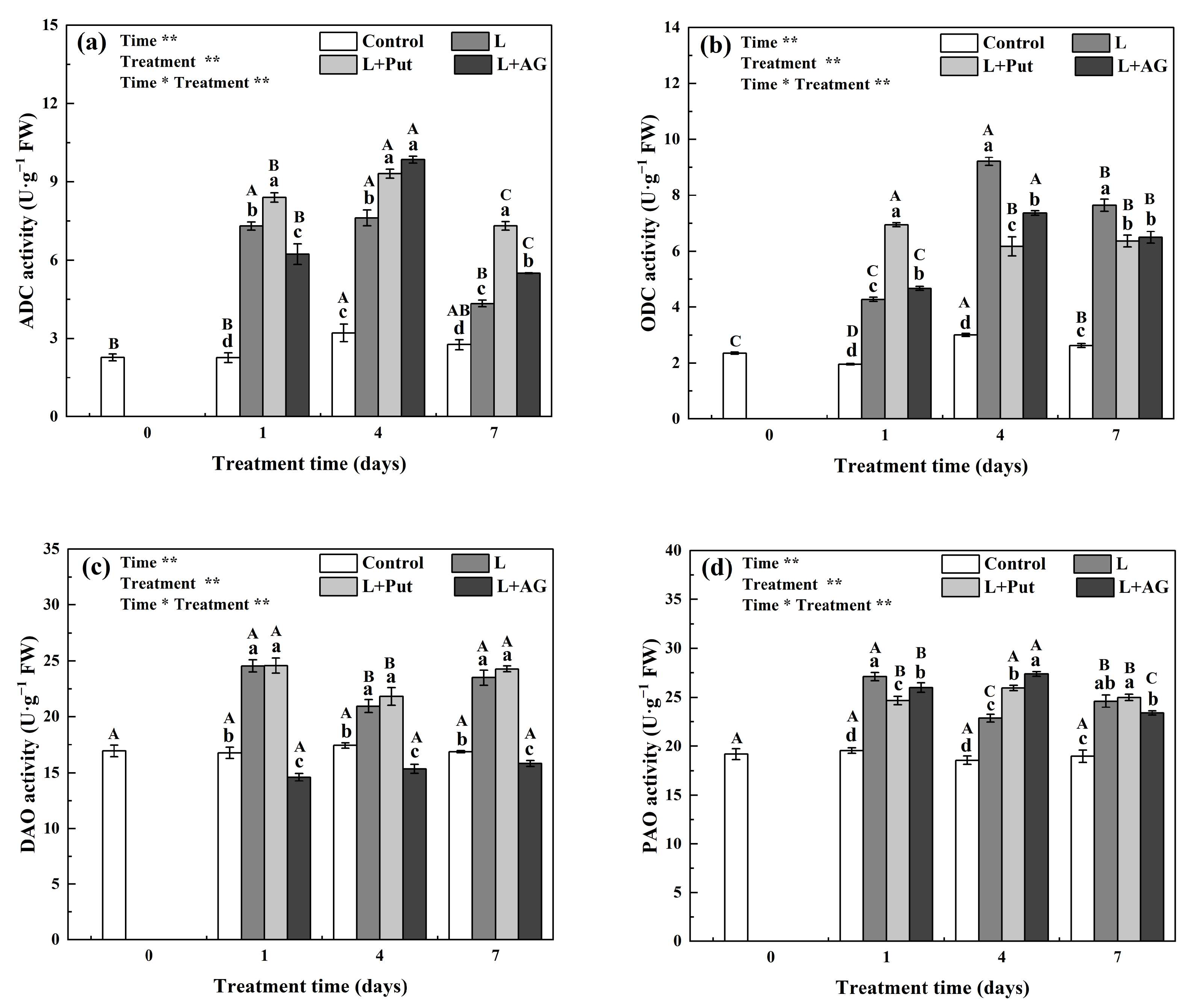
2.4. Determination of Enzyme Activities
2.5. Determination of Hydroperoxide Content, Superoxide Anion Content and Lipid Peroxidation (Malondialdehyde Content)
2.6. Analysis of Antioxidative Enzyme Activities
2.7. Statistical Analysis
3. Results
3.1. γ-Aminobutyric Acid (GABA) Content and GABA Transaminase (GABA-T) Activity
3.2. Glutamic Acid (Glu) Content and Glutamate Decarboxylase (GAD) Activity
3.3. Polyamines (PAs) Contents
3.4. The Enzyme Activities of Putrescine Biosynthetic and Catabolic Pathway
3.5. Contributions of Glu-Derived Pathway and PAs Catabolism Pathway to GABA Accumulation
3.6. The Contents of Hydroperoxide (H2O2), Superoxide Anion (O2∙−), and Malondialdehyde (MDA)
3.7. Antioxidant Enzyme Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Dent, C.E.; Stepka, W.; Steward, F.C. Detection of the free amino-acids of plant cells by partition chromatography. Nature 1947, 160, 682. [Google Scholar] [CrossRef] [PubMed]
- Fait, A.; Fromm, H.; Walter, D.; Galili, G.; Fernie, A.R. Highway or byway: The metabolic role of the GABA shunt in plants. Trends Plant Sci. 2008, 13, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Shelp, B.J.; Bozzo, G.G.; Trobacher, C.P.; Chiu, G.; Bajwa, V.S. Strategies and tools for studying the metabolism and function of γ-aminobutyrate in plants. I. Pathway structure. Botany 2012, 90, 651–668. [Google Scholar] [CrossRef]
- Antonio, C.; Papke, C.; Rocha, M.; Diab, H.; Limami, A.M.; Obata, T.; Fernie, A.R.; van Dongen, J.T. Regulation of primary metabolism in response to low oxygen availability as revealed by carbon and nitrogen isotope redistribution. Plant Physiol. 2016, 170, 43–56. [Google Scholar] [CrossRef]
- Nonaka, S.; Arai, C.; Takayama, M.; Matsukura, C.; Ezura, H. Efficient increase of gamma-aminobutyric acid (GABA) content in tomato fruits by targeted mutagenesis. Sci. Rep. 2017, 7, 7057. [Google Scholar] [CrossRef] [Green Version]
- Scholz, S.S.; Malabarba, J.; Reichelt, M.; Heyer, M.; Ludewig, F.; Mithofer, A. Evidence for GABA-induced systemic GABA accumulation in Arabidopsis upon wounding. Front. Plant Sci. 2017, 8, 388. [Google Scholar] [CrossRef] [Green Version]
- Zarei, A.; Chiu, G.Z.; Yu, G.H.; Trobacher, C.P.; Shelp, B.J. Salinity-regulated expression of genes involved in GABA metabolism and signaling. Botany 2017, 95, 621–627. [Google Scholar] [CrossRef]
- Shelp, B.J.; Bozzo, G.G.; Trobacher, C.P.; Zarei, A.; Deyman, K.L.; Brikis, C.J. Hypothesis/review: Contribution of putrescine to 4-aminobutyrate (GABA) production in response to abiotic stress. Plant Sci. 2012, 193–194, 130–135. [Google Scholar] [CrossRef]
- Wuddineh, W.; Minocha, R.; Minocha, S.C. Polyamines in the context of metabolic networks. In Methods Molecular Biology; Springer: Clifton, NJ, USA, 2018; Volume 1694, pp. 1–23. [Google Scholar]
- Xing, S.G.; Jun, Y.B.; Hau, Z.W.; Liang, L.Y. Higher accumulation of gamma-aminobutyric acid induced by salt stress through stimulating the activity of diamine oxidases in Glycine max (L.) Merr. roots. Plant Physiol. Biochem. 2007, 45, 560–566. [Google Scholar] [CrossRef]
- Liao, J.R.; Wu, X.Y.; Xing, Z.Q.; Li, Q.H.; Duan, Y.; Fang, W.P.; Zhu, X.J. Gamma-aminobutyric acid (GABA) accumulation in tea (Camellia sinensis L.) through the GABA shunt and polyamine degradation pathways under anoxia. J. Agric. Food Chem. 2017, 65, 3013–3018. [Google Scholar] [CrossRef]
- Vera-Sirera, F.; Minguet, E.G.; Singh, S.K.; Ljung, K.; Tuominen, H.; Blazquez, M.A.; Carbonell, J. Role of polyamines in plant vascular development. Plant Physiol. Biochem. 2010, 48, 534–539. [Google Scholar] [CrossRef]
- Romero, F.M.; Maiale, S.J.; Rossi, F.R.; Marina, M.; Ruiz, O.A.; Garriz, A. Polyamine metabolism responses to biotic and abiotic Stress. In Methods in Molecular Biology; Springer: Clifton, NJ, USA, 2018; Volume 1694, pp. 37–49. [Google Scholar]
- Hu, X.; Xu, Z.; Xu, W.; Li, J.; Zhao, N.; Zhou, Y. Application of gamma-aminobutyric acid demonstrates a protective role of polyamine and GABA metabolism in muskmelon seedlings under Ca(NO3)2 stress. Plant Physiol. Biochem. 2015, 92, 1–10. [Google Scholar] [CrossRef]
- Minocha, R.; Majumdar, R.; Minocha, S.C. Polyamines and abiotic stress in plants: A complex relationship. Front. Plant Sci. 2014, 5, 175. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Shao, Q.; Yin, L.; Younis, A.; Zheng, B. Polyamine function in plants: Metabolism, regulation on development, and roles in abiotic stress responses. Front. Plant Sci. 2019, 9, 1945. [Google Scholar] [CrossRef]
- Yong, B.; Xie, H.; Li, Z.; Li, Y.P.; Zhang, Y.; Nie, G.; Zhang, X.Q.; Ma, X.; Huang, L.K.; Yan, Y.H.; et al. Exogenous application of GABA improves PEG-induced drought tolerance positively associated with GABA-shunt, polyamines, and proline metabolism in white clover. Front. Physiol. 2017, 8, 1107. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.J.; Liu, T.; Yang, S.C.; Jin, X.Q.; Qu, F.; Huang, N.; Hu, X.H. Polyamines are involved in GABA-regulated salinity-alkalinity stress tolerance in muskmelon. Environ. Exp. Bot. 2019, 164, 181–189. [Google Scholar] [CrossRef]
- Liu, J.; Yang, R.; Jian, N.; Wei, L.; Ye, L.; Wang, R.; Gao, H.; Zheng, Q. Putrescine metabolism modulates the biphasic effects of brassinosteroids on canola and Arabidopsis salt tolerance. Plant Cell Environ. 2020, 43, 1348–1359. [Google Scholar] [CrossRef]
- Wang, D.; Li, L.; Xu, Y.; Limwachiranon, J.; Li, D.; Ban, Z.; Luo, Z. Effect of exogenous nitro oxide on chilling tolerance, polyamine, proline, and gamma-aminobutyric acid in bamboo shoots (Phyllostachys praecox f. prevernalis). J. Agric. Food Chem. 2017, 65, 5607–5613. [Google Scholar] [CrossRef]
- Pal, M.; Csavas, G.; Szalai, G.; Olah, T.; Khalil, R.; Yordanova, R.; Gell, G.; Birinyi, Z.; Nemeth, E.; Janda, T. Polyamines may influence phytochelatin synthesis during Cd stress in rice. J. Hazard. Mater. 2017, 340, 272–280. [Google Scholar] [CrossRef] [Green Version]
- Mohapatra, S.; Minocha, R.; Long, S.; Minocha, S.C. Putrescine overproduction negatively impacts the oxidative state of poplar cells in culture. Plant Physiol. Biochem. 2009, 47, 262–271. [Google Scholar] [CrossRef]
- Gerlin, L.; Baroukh, C.; Genin, S. Polyamines: Double agents in disease and plant immunity. Trends Plant Sci. 2021, 26, 1061–1071. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Han, Y.; Hao, J.; Qin, X.; Liu, C.; Fan, S. Exogenous spermidine enhances the photosynthesis and ultrastructure of lettuce seedlings under high-temperature stress. Sci. Hortic. 2022, 291, 110570. [Google Scholar] [CrossRef]
- Sen, S.; Ghosh, D.; Mohapatra, S. Modulation of polyamine biosynthesis in Arabidopsis thaliana by a drought mitigating pseudomonas putida strain. Plant Physiol. Biochem. 2018, 129, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.A.; Pang, J.H.; Meng, F.W.; Li, Y.W.; Xu, N.; Yang, C.; Liu, J. Putrescine, spermidine, and spermine play distinct roles in rice salt tolerance. J. Integr. Agr. 2020, 19, 643–655. [Google Scholar] [CrossRef]
- Li, L.J.; Lu, X.C.; Ma, H.Y.; Lyu, D.G. Jasmonic acid regulates the ascorbate-glutathione cycle in Malus baccata Borkh. roots under low root-zone temperature. Acta Physiol. Plant. 2017, 39, 174. [Google Scholar] [CrossRef]
- Li, L.J.; Ma, H.Y.; Lu, X.C.; Lyu, D.G. TMT-Based quantitative proteomic revealed metabolic changes of Jasmonic Acid in regulating the response of Malus baccata (L.) Borkh. roots to low root-zone temperature. J. Plant Growth Regul. 2022, 41, 596–611. [Google Scholar] [CrossRef]
- Lu, X.C.; Li, L.J.; Hu, X.L.; Ma, H.Y.; Lyu, D.G. Effects of exogenous GABA on antioxidant function of Malus Baccata Borkh. roots under suboptimal root-zone temperature. Acta Bot. Boreali-Occident. Sin. 2019, 39, 0285–0293. [Google Scholar]
- Baum, G.; Lev-Yadun, S.; Fridmann, Y.; Arazi, T.; Katsnelson, H.; Zik, M.; Fromm, H. Calmodulin binding to glutamate decarboxylase is required for regulation of glutamate and GABA metabolism and normal development in plants. EMBO J. 1996, 15, 2988–2996. [Google Scholar] [CrossRef]
- Helmond, W.; Puit, M. Amino acid profiling from fingerprints using amide stationary-phase UPLC-MS. In Amino Acid Analysis: Methods and Protocols; Springer: Berlin/Heidelberg, Germany, 2019; Volume 2030, pp. 439–450. [Google Scholar]
- Gong, X.Q.; Liu, J.H. Detection of free polyamines in plants subjected to abiotic stresses by High-Performance Liquid Chromatography (HPLC). In Plant Stress Tolerance: Methods and Protocols; Springer: Berlin/Heidelberg, Germany, 2017; Volume 1631, pp. 305–311. [Google Scholar]
- Tsutsui, H.; Mochizuki, T.; Inoue, K.; Toyama, T.; Yoshimoto, N.; Endo, Y.; Todoroki, K.; Min, J.Z.; Toyo’oka, T. High-throughput LC-MS/MS based simultaneous determination of polyamines including N-acetylated forms in human saliva and the diagnostic approach to breast cancer patients. Anal. Chem. 2013, 85, 11835–11842. [Google Scholar] [CrossRef]
- Takayama, T.; Tsutsui, H.; Shimizu, I.; Toyama, T.; Yoshimoto, N.; Endo, Y.; Inoue, K.; Todoroki, K.; Min, J.Z.; Mizuno, H.; et al. Diagnostic approach to breast cancer patients based on target metabolomics in saliva by liquid chromatography with tandem mass spectrometry. Clin. Chim. Acta 2016, 452, 18–26. [Google Scholar] [CrossRef]
- Patterson, B.D.; MacRae, E.A.; Ferguson, I.B. Estimation of hydrogen peroxide in plant extracts using titanium(IV). Anal. Biochem. 1984, 139, 487–492. [Google Scholar] [CrossRef]
- Verma, S.; Mishra, S.N. Putrescine alleviation of growth in salt stressed Brassica juncea by inducing antioxidative defense system. J. Plant Physiol. 2005, 162, 669–677. [Google Scholar] [CrossRef]
- Liu, Y.J.; Zhao, Z.G.; Si, J.; Di, C.X.; Han, J.; An, L.Z. Brassinosteroids alleviate chilling-induced oxidative damage by enhancing antioxidant defense system in suspension cultured cells of Chorispora bungeana. Plant Growth Regul. 2009, 59, 207–214. [Google Scholar] [CrossRef]
- He, J.L.; Qin, J.J.; Long, L.Y.; Ma, Y.L.; Li, H.; Li, K.; Jiang, X.N.; Liu, T.X.; Polle, A.; Liang, Z.S.; et al. Net cadmium flux and accumulation reveal tissue-specific oxidative stress and detoxification in Populus × canescens. Physiol. Plantarum 2011, 143, 50–63. [Google Scholar] [CrossRef]
- Khan, J.; Malangisha, G.K.; Ali, A.; Mahmoud, A.; Yang, J.; Zhang, M.; Hu, Z. Nitric oxide alleviates lead toxicity by inhibiting lead translocation and regulating root growth in watermelon seedlings. Hortic. Environ. Biote. 2021, 62, 701–714. [Google Scholar] [CrossRef]
- Yang, R.; Chen, H.; Gu, Z. Factors influencing diamine oxidase activity and gamma-aminobutyric acid content of fava bean (Vicia faba L.) during germination. J. Agric. Food Chem. 2011, 59, 11616–11620. [Google Scholar] [CrossRef]
- Shelp, B.J.; Aghdam, M.S.; Flaherty, E.J. γ-Aminobutyrate (GABA) regulated plant defense: Mechanisms and opportunities. Plants 2021, 10, 1939. [Google Scholar] [CrossRef]
- Aghdam, M.S.; Naderi, R.; Jannatizadeh, A.; Babalar, M.; Sarcheshmeh, M.A.; Faradonbe, M.Z. Impact of exogenous GABA treatments on endogenous GABA metabolism in anthurium cut flowers in response to postharvest chilling temperature. Plant Physiol. Biochem. 2016, 106, 11–15. [Google Scholar] [CrossRef]
- Shelp, B.J.; Zarei, A. Subcellular compartmentation of 4-aminobutyrate (GABA) metabolism in Arabidopsis: An update. Plant Signal. Behav. 2017, 12, e1322244. [Google Scholar] [CrossRef] [Green Version]
- Yang, R.Q.; Guo, Q.H.; Gu, Z.X. GABA shunt and polyamine degradation pathway on gamma-aminobutyric acid accumulation in germinating fava bean (Vicia faba L.) under hypoxia. Food Chem. 2013, 136, 152–159. [Google Scholar] [CrossRef]
- You, J.; Chan, Z. ROS regulation during abiotic stress responses in crop plants. Front. Plant Sci. 2015, 6, 1092. [Google Scholar] [CrossRef] [Green Version]
- Puglia, G.D.; Balestrasse, K.; Bustos, J.S.; Huarte, H.R. New insights into the role of alternating temperatures and cyanide in the ROS-mediated cardoon seed dormancy termination. Horticulturae 2022, 8, 960. [Google Scholar] [CrossRef]
- Pospisil, P. Production of reactive oxygen species by photosystem II as a response to light and temperature Stress. Front Plant Sci. 2016, 7, 1950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, B.; Rasmussen, A.; Porter, J.R. Temperatures and the growth and development of maize and rice: A review. Glob. Change Biol. 2014, 20, 408–417. [Google Scholar] [CrossRef]
- Erdal, S.; Genisel, M.; Turk, H.; Dumlupinar, R.; Demir, Y. Modulation of alternative oxidase to enhance tolerance against cold stress of chickpea by chemical treatments. J. Plant Physiol. 2015, 175, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, K.; Sharma, K.D.; Hanumantha Rao, B.; Siddique, K.H.M.; Gaur, P.; Agrawal, S.K.; Nair, R.M.; Nayyar, H. Temperature sensitivity of food legumes: A physiological insight. Acta Physiol. Plant. 2017, 39, 68. [Google Scholar] [CrossRef] [Green Version]
- Akbulut, M.; Cakir, S. The effects of Se phytotoxicity on the antioxidant systems of leaf tissues in barley (Hordeum vulgare L.) seedlings. Plant Physiol. Biochem. 2010, 48, 160–166. [Google Scholar] [CrossRef]
- Sekmen, A.H.; Turkan, I.; Tanyolac, Z.O.; Ozfidan, C.; Dinc, A. Different antioxidant defense responses to salt stress during germination and vegetative stages of endemic halophyte Gypsophila oblanceolata Bark. Environ. Exp. Bot. 2012, 77, 63–76. [Google Scholar] [CrossRef]
- Nouairi, I.; Ben Ammar, W.; Ben Youssef, N.; Ben Miled, D.D.; Ghorbal, M.H.; Zarrouk, M. Antioxidant defense system in leaves of Indian mustard (Brassica juncea) and rape (Brassica napus) under cadmium stress. Acta Physiol. Plant. 2008, 31, 237–247. [Google Scholar] [CrossRef]
- Yan, Q.Y.; Duan, Z.Q.; Mao, J.D.; Li, X.; Dong, F. Low root zone temperature limits nutrient effects on cucumber seedling Growth and induces adversity physiological response. J. Integr. Agric. 2013, 12, 1450–1460. [Google Scholar] [CrossRef]
- Zhang, Y.P.; Jia, F.F.; Zhang, X.M.; Qiao, Y.X.; Shi, K.; Zhou, Y.H.; Yu, J.Q. Temperature effects on the reactive oxygen species formation and antioxidant defence in roots of two cucurbit species with contrasting root zone temperature optima. Acta Physiol. Plant. 2011, 34, 713–720. [Google Scholar] [CrossRef]
- Tiburcio, A.F.; Alcazar, R. Potential applications of polyamines in agriculture and plant biotechnology. In Methods in Mmolecular Biology; Springer: Clifton, NJ, USA, 2018; Volume 1694, pp. 489–508. [Google Scholar]
- Bueno, M.; Cordovilla, M.P. Polyamines in Halophytes. Front. Plant Sci. 2019, 10, 439. [Google Scholar] [CrossRef] [Green Version]
- Kielkowska, A.; Adamus, A. Exogenously applied polyamines reduce reactive oxygen species, enhancing cell division and the shoot regeneration from Brassica oleracea L. var. capitata protoplasts. Agronomy 2021, 11, 735. [Google Scholar] [CrossRef]
- Pal, M.; Majlath, I.; Nemeth, E.; Hamow, K.A.; Szalai, G.; Rudnoy, S.; Balassa, G.; Janda, T. The effects of putrescine are partly overlapping with osmotic stress processes in wheat. Plant Sci. 2018, 268, 67–76. [Google Scholar] [CrossRef]
- Sardar, R.; Ahmed, S.; Yasin, N.A. Role of exogenously applied putrescine in amelioration of cadmium stress in Coriandrum sativum by modulating antioxidant system. Int. J. Phytoremediat. 2022, 24, 955–962. [Google Scholar] [CrossRef]
- Collado-Gonzalez, J.; Pinero, M.C.; Otalora, G.; Lopez-Marin, J.; Amor, F.M.D. The Effect of foliar putrescine application, ammonium exposure, and heat stress on antioxidant compounds in cauliflower waste. Antioxidants 2021, 10, 707. [Google Scholar] [CrossRef]
- Izadi, Z.; Nejad, A.R.; Abadia, J. Foliar applications of thidiazuron and putrescine increase leaf iron and chlorophyll concentrations in iron-deficient pot marigold (Calendula officinalis L.). Acta Physiol. Plant. 2021, 43, 122. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Irrigation (100 mL/pot) | Upper Part | Potted Part |
|---|---|---|---|
| Control | dH2O | 18 °C (14 h)/8 °C (10 h) | 18 °C (14 h)/8 °C (10 h) |
| L | dH2O | 18 °C (14 h)/8 °C (10 h) | 5 ± 0.5 °C (24 h) |
| L + Put | 0.1 mmol·L−1 Put | 18 °C (14 h)/8 °C (10 h) | 5 ± 0.5 °C (24 h) |
| L + AG | 10 mmol·L−1 AG | 18 °C (14 h)/8 °C (10 h) | 5 ± 0.5 °C (24 h) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, X.; Zhao, M.; Zhou, E.; Ma, H.; Lyu, D. Contribution of Putrescine and Glutamic Acid on γ-Aminobutyric Acid Accumulation of Malus baccata Borkh. Roots under Suboptimal Low Root-Zone Temperature. Agronomy 2023, 13, 1989. https://doi.org/10.3390/agronomy13081989
Lu X, Zhao M, Zhou E, Ma H, Lyu D. Contribution of Putrescine and Glutamic Acid on γ-Aminobutyric Acid Accumulation of Malus baccata Borkh. Roots under Suboptimal Low Root-Zone Temperature. Agronomy. 2023; 13(8):1989. https://doi.org/10.3390/agronomy13081989
Chicago/Turabian StyleLu, Xiaochen, Meiqi Zhao, Enda Zhou, Huaiyu Ma, and Deguo Lyu. 2023. "Contribution of Putrescine and Glutamic Acid on γ-Aminobutyric Acid Accumulation of Malus baccata Borkh. Roots under Suboptimal Low Root-Zone Temperature" Agronomy 13, no. 8: 1989. https://doi.org/10.3390/agronomy13081989
APA StyleLu, X., Zhao, M., Zhou, E., Ma, H., & Lyu, D. (2023). Contribution of Putrescine and Glutamic Acid on γ-Aminobutyric Acid Accumulation of Malus baccata Borkh. Roots under Suboptimal Low Root-Zone Temperature. Agronomy, 13(8), 1989. https://doi.org/10.3390/agronomy13081989