
Modern Plant Biotechnology: An Antidote against Global Food Insecurity

 ,
,  , ,
, ,
Abstract
:1. Introduction
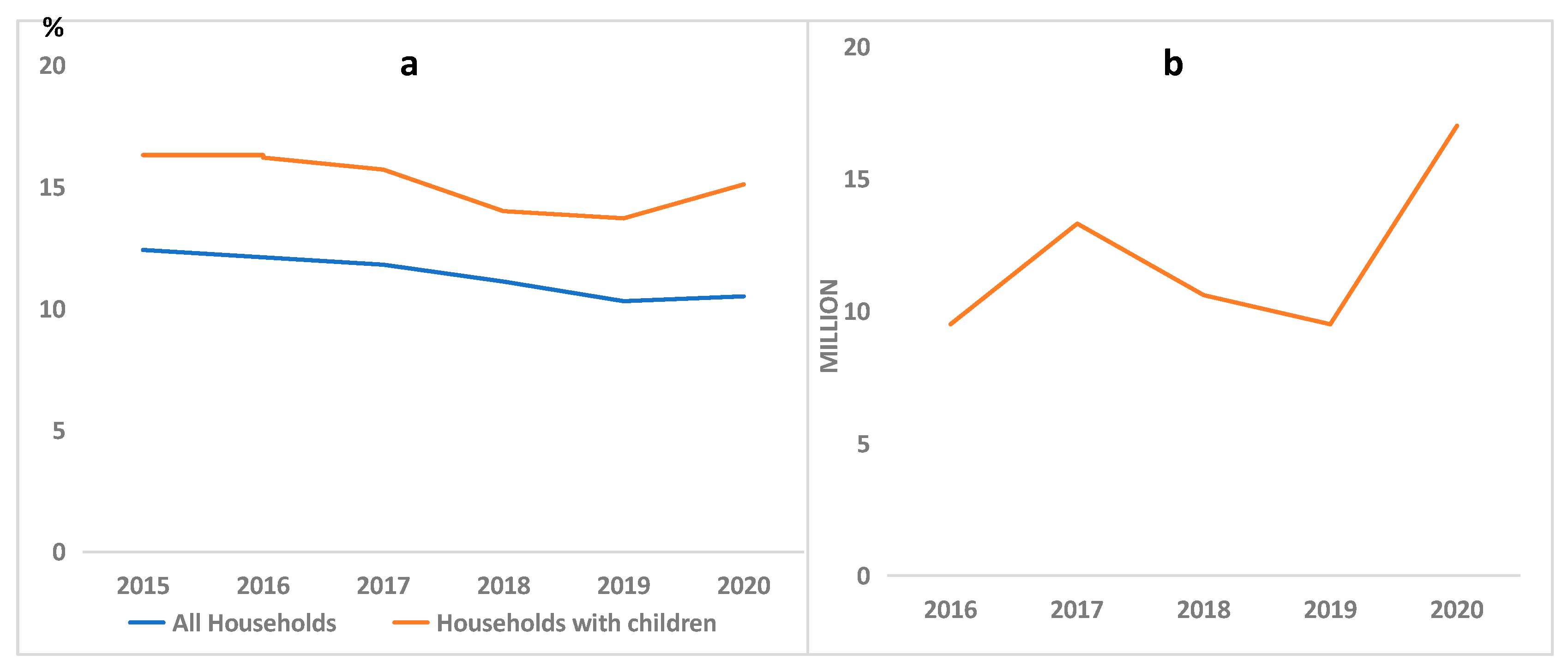
1.1. Food Insecurity: A Global Menace
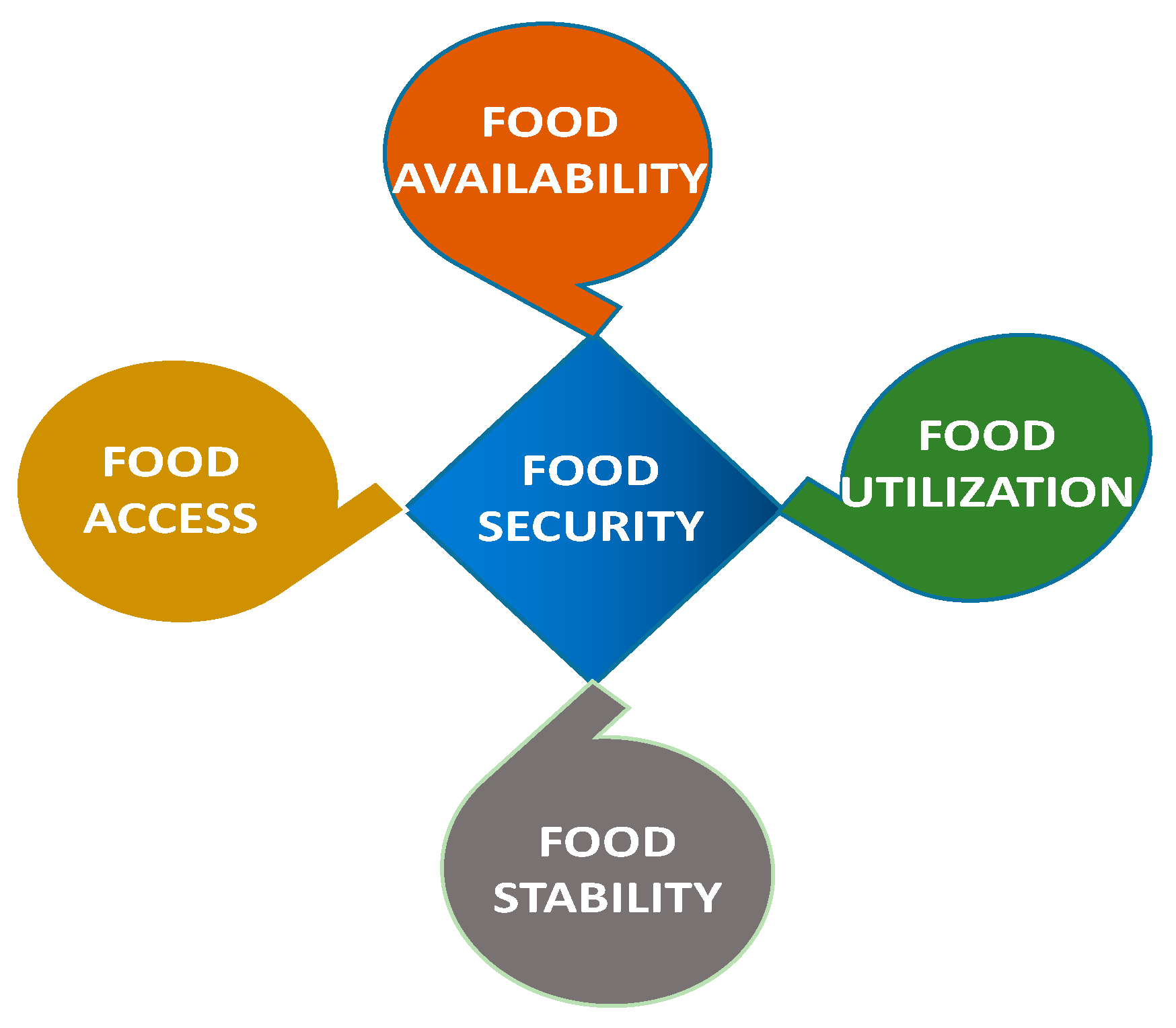
1.2. Defining a Food-Secure World
2. Biotechnology: A Viable Approach to Curbing Food Insecurity
Genetic Modification of Crops: A Silver Bullet to Combat Food Insecurity
3. Genetically Modified Crops; A Masterpiece
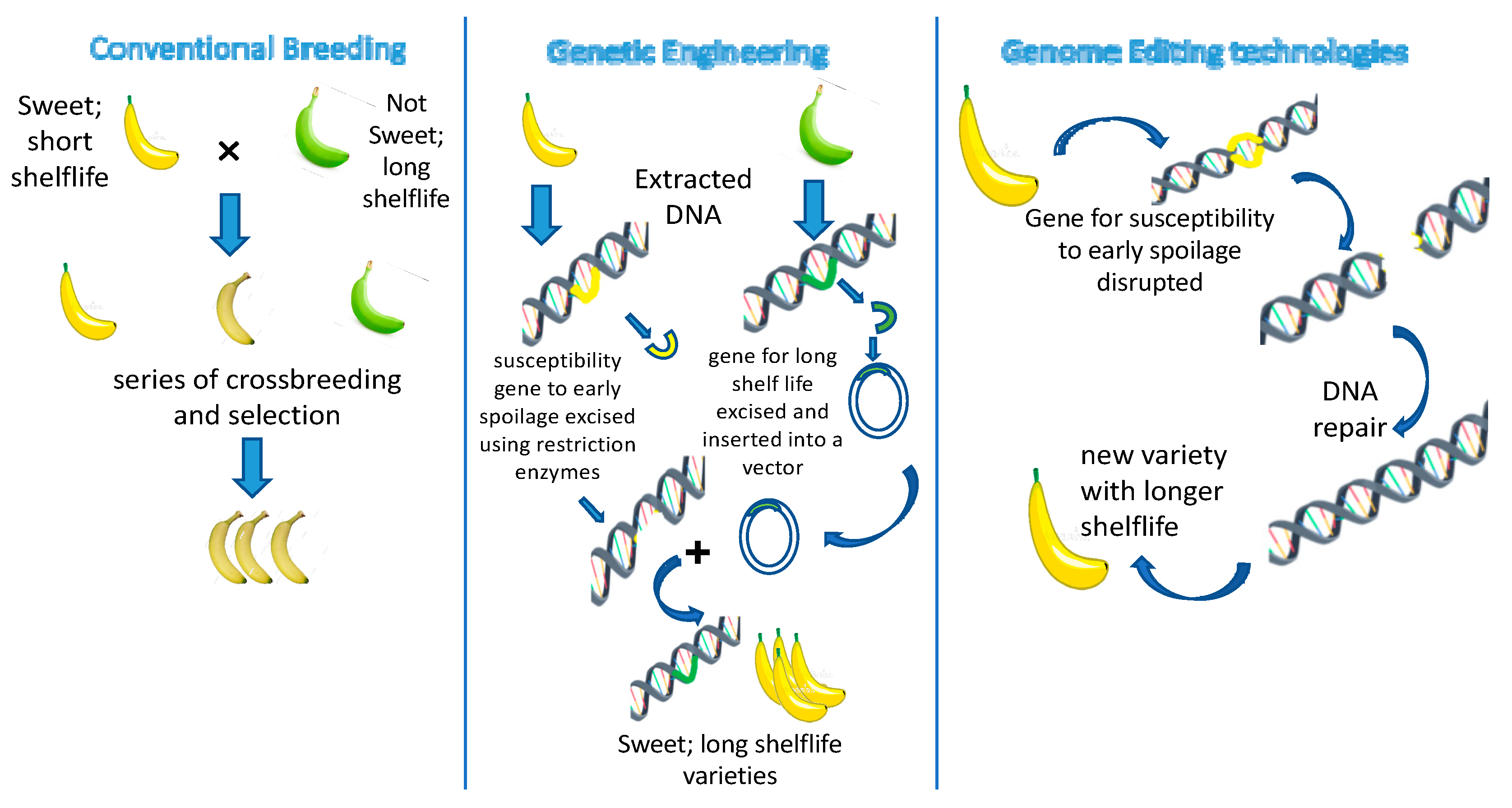
3.1. Developing Crop Varieties with Higher Yield and Extended Shelf Life
3.2. Insect Tolerance and Herbicide Resistance
3.3. Enhanced Nutritional Content
3.4. Plant-Derived Edible Vaccines
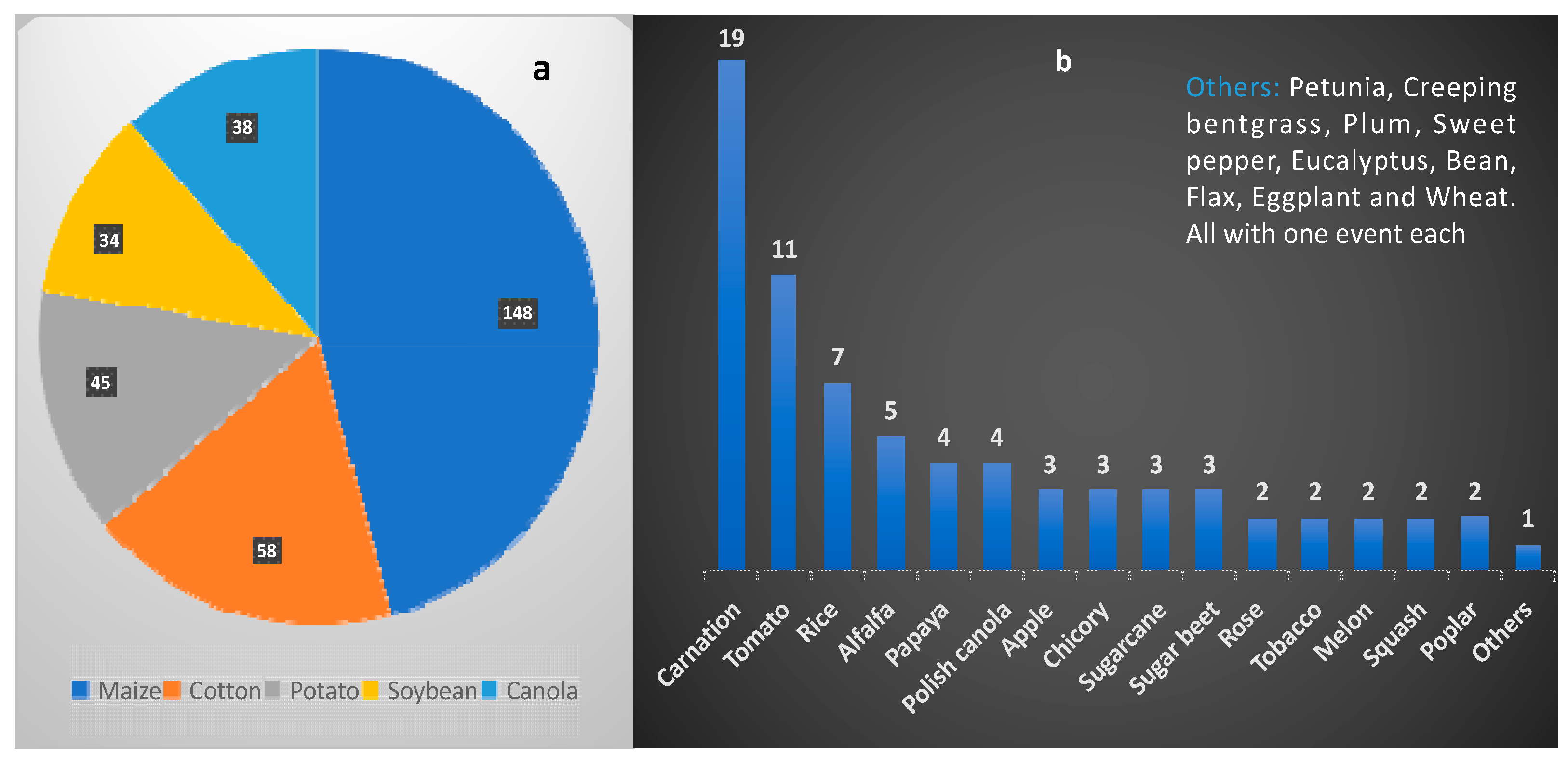
4. Global Acceptance and Perspectives on Biotechnologically Developed Crops
4.1. Europe and Africa
4.2. GMO-Free CRISPR/Cas9 Crops—Global Thoughts and Acceptance
4.3. Opposition to Global Acceptance of Biotech Crops
4.4. Contributions to Food Security and Human Health
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Basonde, R.; Andhare, P. GM crops is a solution for world food crisis? Asian J. Microbiol. Biotechnol. Environ. Sci. 2014, 17, 163–166. [Google Scholar]
- Bickel, G.; Nord, M.; Price, C.; Hamilton, W.; Cook, J. Guide to Measuring Household Food Security; USDA Food and Nutrition Service: Alexandria, VA, USA, 2000; p. 82. [Google Scholar]
- FAO; IFAD; UNICEF; WFP; WHO. The State of Food Security and Nutrition in the World 2020. Transforming Food Systems for Affordable Healthy Diets; FAO: Rome, Italy, 2020. [Google Scholar] [CrossRef]
- World Food Programme. A Global Food Crisis. 2022. Available online: www.wfp.org/global-hunger-crisis (accessed on 26 June 2023).
- Habibi-Najafi, M.B.; Lee, B.H. Biotechnology and its Impact on Food Security and Safety. Curr. Nutr. Food Sci. 2014, 10, 94–99. [Google Scholar] [CrossRef]
- Webb, P.; Stordalen, G.A.; Singh, S.; Wijesinha-Bettoni, R.; Shetty, P.; Lartey, A. Hunger and malnutrition in the 21st century. BMJ 2018, 361, k2238. [Google Scholar] [CrossRef]
- Militao, E.M.A.; Salvador, E.M.; Uthman, O.A.; Vinberg, S.; Macassa, G. Food Insecurity and Health Outcomes Other than Malnutrition in Southern Africa: A Descriptive Systematic Review. Int. J. Environ. Res. Public. Health. 2022, 21, 5082. [Google Scholar] [CrossRef]
- GNAFC; FSIN. 2020 Global Report on Food Crisis. 2020. Available online: https://www.fsinplatform.org/sites/default/files/resources/files/GRFC_2020_KM_200420.pdf (accessed on 6 July 2023).
- Clark, H.; Coll-Seck, A.M.; Banerjee, A.; Peterson, S.; Dalglish, S.L.; Ameratunga, S.; Balabanova, D.; Bhan, M.K.; Bhutta, Z.A.; Borrazzo, J.; et al. A future for the world’s children? A WHO–UNICEF–Lancet Commission. Lancet 2020, 395, 605–658. [Google Scholar] [CrossRef]
- Govender, I.; Rangiah, S.; Kaswa, R.; Nzaumvila, D. Malnutrition in children under the age of 5 years in a primary health care setting. S. Afr. Fam. Pract. 2021, 63, e1–e6, Erratum in S. Afr. Fam. Pract. 2021, 63, 5416. [Google Scholar] [CrossRef]
- IFPRI. 2015 Annual Report: International Food Policy Research Institute (IFPRI). 2016. Available online: http://ebrary.ifpri.org/cdm/ref/collection/p15738coll2/id/130442 (accessed on 1 July 2023).
- FAO; IFAD; UNICEF; WFP; WHO. The State of Food Security and Nutrition in the World 2022: Re-Purposing Food and Agricultural Policies to Make Healthy Diets More Affordable; FAO: Rome, Italy, 2022; 260p, ISBN 978-92-5-136499-4. [Google Scholar] [CrossRef]
- Coleman-Jensen, A.; Rabbitt, M.P.; Gregory, C.A.; Singh, A. Household Food Security in the United States in 2020; USDA, Economic Research Service: Washington, DC, USA, 2021; 56p. Available online: https://www.ers.usda.gov/webdocs/publications/102076/err-298.pdf (accessed on 2 July 2023).
- SWAC/OECD. Food and Nutrition Crisis 2020: Analyses & Responses; Maps & Facts; OECD: Paris, France, 2020; No. 3; 56p, Available online: https://www.oecd.org/swac/maps/Food-nutrition-crisis-2020-Sahel-West-Africa_EN.pdf (accessed on 22 June 2023).
- Swaminathan, M.S. Science in response to basic human needs. Science 2000, 284, 425. [Google Scholar]
- Tonukari, N.J.; Omotor, D.G. Biotechnology and food security in developing countries. Biotechnol. Mol. Biol. Rev. 2010, 5, 13–23. [Google Scholar]
- Habibi-Najafi, M.B. Food Biotechnology and its impact on our food supply. J. Biochem. Biotechnol. 2006, 1, 22–27. [Google Scholar]
- Peng, W.; Berry, E.M. The Concept of Food Security. In Encyclopedia of Food Security and Sustainability; Ferranti, P., Berry, E.M., Anderson, J.R., Eds.; Elsevier: Cambridge, MA, USA, 2019; Volume 2, pp. 1–7. ISBN 9780128126875. [Google Scholar]
- United Nations. Report of the World Food Conference, Rome, 5–16 November 1974. 1975. Available online: https://digitallibrary.un.org/record/701143?ln=en (accessed on 30 June 2023).
- FAO. World Food Security: A Reappraisal of the Concepts and Approaches: Director General’s Report. 1983. Available online: https://www.fao.org/3/AK626E/ak626e08.htm (accessed on 30 June 2023).
- World Bank. Poverty and Hunger: Issues and Options for Food Security in Developing Countries. 1986. Available online: https://documents1.worldbank.org/curated/en/166331467990005748/pdf/multi-page.pdf (accessed on 30 June 2023).
- FAO. Rome Declaration on Food Security and World Food Summit Plan of Action. 1996. Available online: https://www.fao.org/3/w3548e/w3548e00.htm (accessed on 22 June 2023).
- FAO. Declaration of the World Food Summit on Food Security. 2009. Available online: https://www.fao.org/fileadmin/templates/wsfs/Summit/Docs/Final_Declaration/WSFS09_Declaration.pdf (accessed on 30 June 2023).
- Chen, R.S.; Kates, R.W. World food security: Prospects and trends. Food Policy 1994, 19, 192–208. [Google Scholar] [CrossRef]
- Gil, R. Addressing Food Insecurity in SLP with a Food Security Task Force. Seed Feeds. 2021. Available online: https://seedsfeeds.org/programs-resources-news/addressing-food-insecurity (accessed on 30 June 2023).
- Khan, F.; Hasan, A. Genetically Modified Organisms: A Solution to Food Security and Environment. Int. J. Soc. Sci. 2016, 6, 1–12. [Google Scholar]
- Lamichhane, S.A. Genetically Modified Foods- Solution for food security. Int. J. Genet. Eng. Biotechnol. 2014, 5, 43–48. [Google Scholar]
- Reddy, P.B. Framing GM crops as a solution for Global Food Security. Int. J. Res. Granthaalayah 2015, 3, 1–5. [Google Scholar]
- Jamil, K. Biotechnology, a Solution to Hunger: UN Chronicle. 2012. Available online: www.un.org/en/chronicle/article/biotechnology-solution-hunger (accessed on 20 June 2023).
- Verma, A.S.; Agrahari, S.; Rastogi, S.; Singh, A. Biotechnology in the Realm of History. J. Pharm. Bioallied Sci. 2011, 3, 321–323. [Google Scholar] [CrossRef]
- Animasaun, D.A.; Azeez, M.A.; Adubi, A.O.; Durodola, F.A.; Morakinyo, J.A. Trends in genetically modified crops development in Nigeria: Issues and challenges. In Genetically Modified and Irradiated Food Controversial Issues: Facts Versus Perceptions; Andersen, V., Ed.; Elsevier: Amsterdam, The Netherlands; Academic Press: London, UK, 2020; pp. 131–150. [Google Scholar]
- Then, C.; Österreicher, A. Differences between Conventional Breeding and Genetic Engineering: An Assessment of the Statement Made by the Group of Chief Scientific Advisors’ (SAM). TESTBIOTECH. 8pp. 2019. Available online: https://www.testbiotech.org/en/content/differences-between-conventional-breeding-and-genetic-engineering-sam (accessed on 15 June 2023).
- Manshardt, R. Crop Improvement by Conventional Breeding or Genetic Engineering: How Different Are They? Biotechnology 2004, 5, 3. [Google Scholar]
- Jamil, S.; Shahzad, R.; Ahmad, S.; Fatima, R.; Zahid, R.; Anwar, M.; Iqbal, M.Z.; Wang, X. Role of Genetics, Genomics, and Breeding Approaches to Combat Stripe Rust of Wheat. Front. Nutr. 2020, 7, 580715. [Google Scholar] [CrossRef]
- Klümper, W.; Qaim, M. A meta-analysis of the impacts of genetically modified crops. PLoS ONE 2014, 9, e111629. [Google Scholar] [CrossRef]
- Animasaun, D.A.; Adedibu, P.A.; Joseph, G.G. Exploring the potentials of microalgae as an alternative source of renewable energy. Niger. J. Biotechnol. 2021, 38, 24–39. [Google Scholar] [CrossRef]
- Lemaux, P.G. Genetically Engineered Plants and Foods: A Scientist’s Analysis of the Issues (Part I). Annu. Rev. Plant Biol. 2008, 59, 771–812. [Google Scholar]
- Bruening, G.; Lyons, J.M. The case of the FLAVR SAVR tomato. Calif. Agric. 2000, 54, 6–7. [Google Scholar] [CrossRef]
- Shetty, M.J.; Chandan, K.; Krishna, H.C.; Aparna, G.S. Genetically Modified Crops: An Overview. J. Pharmacogn. Phytochem. 2018, 7, 2405–2410. [Google Scholar]
- Wada, N.; Ueta, R.; Osakabe, Y.; Osakabe, K. Precision genome editing in plants: State-of-the-art in CRISPR/Cas9-based genome engineering. BMC Plant Biol. 2020, 20, 234. [Google Scholar] [CrossRef]
- Ahmad, M. Plant breeding advancements with “CRISPR-Cas” genome editing technologies will assist future food security. Front. Plant Sci. 2023, 14, 1133036. [Google Scholar] [CrossRef]
- Liu, X.; Wu, S.; Xu, J.; Sui, C.; Wei, J. Application of CRISPR/Cas9 in plant biology. Acta Pharm. Sin. B 2017, 7, 292–302. [Google Scholar] [CrossRef]
- Yin, K.; Qiu, J. Genome editing for plant disease resistance: Applications and perspectives. Phil. Trans. R. Soc. 2019, 374, 2018032220180322. [Google Scholar] [CrossRef]
- Asmamaw, M.; Zawdie, B. Mechanism and Applications of CRISPR/Cas-9-Mediated Genome Editing. Biologics 2021, 21, 353–361. [Google Scholar] [CrossRef]
- Liu, Q.; Zhao, K.; Wang, C.; Zhang, Z.; Zheng, C.; Zhao, Y.; Zheng, Y.; Liu, C.; An, Y.; Shi, L.; et al. Multistage delivery nanoparticle facilitates efficient CRISPR/dCas9 activation and tumor growth suppression in vivo. Adv. Sci. 2019, 6, 1801423. [Google Scholar] [CrossRef]
- Li, Z.; Liu, Z.B.; Xing, A.; Moon, B.P.; Koellhoffer, J.P.; Huang, L.; Ward, R.T.; Clifton, E.; Falco, S.C.; Cigan, A.M. Cas9-Guide RNA directed genome editing in soybean. Plant Physiol. 2015, 169, 960–970. [Google Scholar] [CrossRef]
- Xu, R.; Yang, Y.; Qin, R.; Li, H.; Qiu, C.; Li, L.; Wei, P.; Yang, J. Rapid improvement of grain weight via highly efficient CRISPR/Cas9-mediated multiplex genome editing in rice. J. Genet. Genom. 2016, 43, 529. [Google Scholar]
- Dong, O.X.; Yu, S.; Jain, R.; Zhang, N.; Duong, P.Q.; Butler, C.; Li, Y.; Lipzen, A.; Martin, J.A.; Barry, K.W.; et al. Marker-free carotenoid-enriched rice generated through targeted gene insertion using CRISPR-Cas9. Nat Commun. 2020, 11, 1178. [Google Scholar] [CrossRef]
- Huang, L.; Li, Q.; Zhang, C.; Chu, R.; Gu, Z.; Tan, H.; Zhao, D.; Fan, X.; Liu, Q. Creating novel Wx alleles with fine-tuned amylose levels and improved grain quality in rice by promoter editing using CRISPR/Cas9 system. Plant Biotechnol. J. 2020, 18, 2164–2166. [Google Scholar] [CrossRef]
- Wang, W.; Pan, Q.; He, F.; Akhunova, A.; Chao, S.; Trick, H.; Akhunov, E. Transgenerational CRISPR-Cas9 activity facilitates multiplex gene editing in allopolyploid wheat. CRISPR J. 2018, 1, 65–74. [Google Scholar] [CrossRef]
- Wang, W.; Pan, Q.; Tian, B.; He, F.; Chen, Y.; Bai, G.; Akhunova, A.; Trick, H.N.; Akhunov, E. Gene editing of the wheat homologs of TONNEAU1-recruiting motif encoding gene affects grain shape and weight in wheat. Plant J. 2019, 100, 251–264. [Google Scholar] [CrossRef]
- Zhu, J.; Song, N.; Sun, S.; Yang, W.; Zhao, H.; Song, W.; Lai, J. Efficiency and inheritance of targeted mutagenesis in maize using CRISPR-Cas9. J. Genet. Genom. 2016, 43, 25–36. [Google Scholar] [CrossRef]
- D’Ambrosio, C.; Stigliani, A.L.; Giorio, G. CRISPR/Cas9 editing of carotenoid genes in tomato. Transgenic Res. 2018, 27, 367–378. [Google Scholar] [CrossRef]
- Wang, D.; Samsulrizal, N.H.; Yan, C.; Allcock, N.S.; Craigon, J.; Blanco-Ulate, B.; Ortega-Salazar, I.; Marcus, S.E.; Bagheri, H.M.; Fons, L.P. Characterization of CRISPR mutants targeting genes modulating pectin degradation in ripening tomato. Plant Physiol. 2019, 179, 544–557. [Google Scholar]
- Yuste-Lisbona, F.J.; Fernández-Lozano, A.; Pineda, B.; Bretones, S.; Ortíz-Atienza, A.; García-Sogo, B.; Müller, N.A.; Angosto, T.; Capel, J.; Moreno, V.; et al. ENO regulates tomato fruit size through the floral meristem development network. Proc. Natl. Acad. Sci. USA 2020, 117, 8187–8195. [Google Scholar] [CrossRef]
- Bull, S.E.; Seung, D.; Chanez, C.; Mehta, D.; Kuon, J.-E.; Truernit, E.; Hochmuth, A.; Zurkirchen, I.; Zeeman, S.C.; Gruissem, W. Accelerated ex situ breeding of GBSS-and PTST1-edited cassava for modified starch. Sci. Adv. 2018, 4, eaat6086. [Google Scholar] [CrossRef]
- Tuncel, A.; Corbin, K.R.; Ahn-Jarvis, J.; Harris, S.; Hawkins, E.; Smedley, M.A.; Harwood, W.; Warren, F.J.; Patron, N.J.; Smith, A.M. Cas9-mediated mutagenesis of potato starch-branching enzymes generates a range of tuber starch phenotypes. Plant Biotechnol. J. 2019, 17, 2259–2271. [Google Scholar] [CrossRef]
- Veillet, F.; Chauvin, L.; Kermarrec, M.-P.; Sevestre, F.; Merrer, M.; Terret, Z.; Szydlowski, N.; Devaux, P.; Gallois, J.-L.; Chauvin, J.-E. The solanum tuberosum GBSSI gene: A target for assessing gene and base editing in tetraploid potato. Plant Cell Rep. 2019, 38, 1065–1080. [Google Scholar] [CrossRef]
- Zsögön, A.; Čermák, T.; Naves, E.R.; Notini, M.M.; Edel, K.H.; Weinl, S.; Freschi, L.; Voytas, D.F.; Kudla, J.; Peres, L.E.P. De novo domestication of wild tomato using genome editing. Nat. Biotechnol. 2018, 36, 1211–1216. [Google Scholar]
- Rodríguez-Leal, D.; Lemmon, Z.H.; Man, J.; Bartlett, M.E.; Lippman, Z.B. Engineering quantitative trait variation for crop improvement by genome editing. Cell 2017, 171, 470–480. [Google Scholar] [PubMed]
- Lemmon, Z.H.; Reem, N.T.; Dalrymple, J.; Soyk, S.; Swartwood, K.E.; Rodriguez-Leal, D.; Van Eck, J.; Lippman, Z.B. Rapid improvement of domestication traits in an orphan crop by genome editing. Nat. Plants 2018, 4, 766–770. [Google Scholar]
- Zhou, J.; Xin, X.; He, Y.; Chen, H.; Li, Q.; Tang, X.; Zhong, Z.; Deng, K.; Zheng, X.; Akher, S.A. Multiplex QTL editing of grain-related genes improves yield in elite rice varieties. Plant Cell Rep. 2019, 38, 475–485. [Google Scholar]
- Shen, L.; Wang, C.; Fu, Y.; Wang, J.; Liu, Q.; Zhang, X.; Yan, C.; Qian, Q.; Wang, K. QTL editing confers opposing yield performance in different rice varieties. J. Integr. Plant Biol. 2018, 60, 89–93. [Google Scholar]
- Zhao, D.-S.; Li, Q.-F.; Zhang, C.-Q.; Zhang, C.; Yang, Q.-Q.; Pan, L.-X.; Ren, X.-Y.; Lu, J.; Gu, M.-H.; Liu, Q.-Q. GS9 acts as a transcriptional activator to regulate rice grain shape and appearance quality. Nat. Commun. 2018, 9, 1240. [Google Scholar]
- Vu, T.V.; Sivankalyani, V.; Kim, E.-J.; Doan, D.T.H.; Tran, M.T.; Kim, J.; Sung, Y.W.; Park, M.; Kang, Y.J.; Kim, J.-Y. Highly efficient homology-directed repair using CRISPR/Cpf1-geminiviral replicon in tomato. Plant Biotechnol. J. 2020, 18, 2133–2143. [Google Scholar]
- Ben Shlush, I.; Samach, A.; Melamed-Bessudo, C.; Ben-Tov, D.; Dahan-Meir, T.; Filler-Hayut, S.; Levy, A.A. CRISPR/Cas9 induced somatic recombination at the CRTISO locus in tomato. Genes 2021, 12, 59. [Google Scholar]
- Deng, L.; Wang, H.; Sun, C.; Li, Q.; Jiang, H.; Du, M.; Li, C.-B.; Li, C. Efficient generation of pink-fruited tomatoes using CRISPR/Cas9 system. J. Genet. Genom. 2018, 45, 51–54. [Google Scholar]
- Watanabe, K.; Oda-Yamamizo, C.; Sage-Ono, K.; Ohmiya, A.; Ono, M. Alteration of flower colour in Ipomoea nil through CRISPR/Cas9-mediated mutagenesis of carotenoid cleavage dioxygenase 4. Transgenic Res. 2018, 27, 25–38. [Google Scholar]
- Yu, J.; Tu, L.; Subburaj, S.; Bae, S.; Lee, G.-J. Simultaneous targeting of duplicated genes in Petunia protoplasts for flower color modification via CRISPR-Cas9 ribonucleoproteins. Plant Cell Rep. 2020, 40, 1037–1045. [Google Scholar] [PubMed]
- Zhong, Y.; Blennow, A.; Kofoed-Enevoldsen, O.; Jiang, D.; Hebelstrup, K.H. Protein Targeting to Starch 1 is essential for starchy endosperm development in barley. J. Exp. Bot. 2019, 70, 485–496. [Google Scholar] [PubMed]
- Sun, Y.; Jiao, G.; Liu, Z.; Zhang, X.; Li, J.; Guo, X.; Du, W.; Du, J.; Francis, F.; Zhao, Y.; et al. Generation of high-amylose rice through CRISPR/Cas9-mediated targeted mutagenesis of starch branching enzymes. Front. Plant Sci. 2017, 8, 298. [Google Scholar] [PubMed]
- Abe, K.; Araki, E.; Suzuki, Y.; Toki, S.; Saika, H. Production of high oleic/low linoleic rice by genome editing. Plant Physiol. Biochem. 2018, 131, 58–62. [Google Scholar]
- Okuzaki, A.; Ogawa, T.; Koizuka, C.; Kaneko, K.; Inaba, M.; Imamura, J.; Koizuka, N. CRISPR/Cas9-mediated genome editing of the fatty acid desaturase 2 gene in Brassica napus. Plant Physiol. Biochem. 2018, 131, 63–69. [Google Scholar]
- Jiang, W.Z.; Henry, I.M.; Lynagh, P.G.; Comai, L.; Cahoon, E.B.; Weeks, D.P. Significant enhancement of fatty acid composition in seeds of the allohexaploid, Camelina sativa, using CRISPR/Cas9 gene editing. Plant Biotechnol. J. 2017, 15, 648–657. [Google Scholar]
- Sashidhar, N.; Harloff, H.J.; Potgieter, L.; Jung, C. Gene editing of three BnITPK genes in tetraploid oilseed rape leads to significant reduction of phytic acid in seeds. Plant Biotechnol. J. 2020, 18, 2241–2250. [Google Scholar]
- Khan, M.S.S.; Basnet, R.; Islam, S.A.; Shu, Q. Mutational analysis of OsPLDα1 reveals its involvement in phytic acid biosynthesis in rice grains. J. Agric. Food Chem. 2019, 67, 11436–11443. [Google Scholar]
- Sanchez-Leon, S.; Gil-Humanes, J.; Ozuna, C.V.; Gimenez, M.J.; Sousa, C.; Voytas, D.F.; Barro, F. Low-gluten, nontransgenic wheat engineered with CRISPR/Cas9. Plant Biotechnol. J. 2018, 16, 902–910. [Google Scholar]
- Nonaka, S.; Arai, C.; Takayama, M.; Matsukura, C.; Ezura, H. Efficient increase of ɣ-aminobutyric acid (GABA) content in tomato fruits by targeted mutagenesis. Sci. Rep. 2017, 7, 7057. [Google Scholar]
- Akama, K.; Akter, N.; Endo, H.; Kanesaki, M.; Endo, M.; Toki, S. An in vivo targeted deletion of the calmodulin-binding domain from rice glutamate decarboxylase 3 (OsGAD3) increases γ-aminobutyric acid content in grains. Rice 2020, 13, 20. [Google Scholar] [PubMed]
- Wang, F.; Wang, C.; Liu, P.; Lei, C.; Hao, W.; Gao, Y.; Liu, Y.G.; Zhao, K. Enhanced rice blast resistance by CRISPR/Cas9-targeted mutagenesis of the ERF transcription factor gene OsERF922. PLoS ONE 2016, 11, e0154027. [Google Scholar] [CrossRef]
- Zhou, J.; Peng, Z.; Long, J.; Sosso, D.; Liu, B.; Eom, J.S.; Huang, S.; Liu, S.; Vera Cruz, C.; Frommer, W.B.; et al. Gene targeting by the TAL effector PthXo2 reveals cryptic resistance gene for bacterial blight of rice. Plant J. 2015, 82, 632–643. [Google Scholar] [CrossRef] [PubMed]
- Oliva, R.; Ji, C.; Atienza-Grande, G.; Huguet-Tapia, J.C.; Perez-Quintero, A.; Li, T.; Eom, J.S.; Li, C.; Nguyen, H.; Liu, B.; et al. Broad-spectrum Resistance to Bacterial Blight in Rice Using Genome Editing. Nat. Biotechnol. 2019, 37, 1344–1350. [Google Scholar] [CrossRef] [PubMed]
- Santillan Martinez, M.I.; Bracuto, V.; Koseoglou, E.; Appiano, M.; Jacobsen, E.; Visser, R.G.F.; Wolters, A.M.A.; Bai, Y. CRISPR/Cas9-targeted Mutagenesis of the Tomato Susceptibility Gene PMR4 for Resistance against Powdery Mildew. BMC Plant Biol. 2020, 20, 284. [Google Scholar] [CrossRef]
- Ortigosa, A.; Gimenez-Ibanez, S.; Solano, R. Design of a Bacterial Speck Resistant Tomato by CRISPR/Cas9-mediated Editing of SlJAZ2. Plant Biotechnol. J. 2019, 17, 665–673. [Google Scholar] [CrossRef]
- Gomez, M.A.; Lin, D.Z.; Moll, T.; Chauhan, R.D.; Hayden, L.; Renninger, K.; Beyene, G.; Taylor, N.J.; Carrington, J.C.; Staskawicz, B.J.; et al. Simultaneous CRISPR/Cas9-Mediated Editing of Cassava eIF4E Isoforms nCBP-1 and nCBP-2 Reduces Cassava Brown Streak Disease Symptom Severity and Incidence. Plant Biotechnol. J. 2019, 17, 421–434. [Google Scholar] [CrossRef]
- Baltes, N.J.; Hummel, A.W.; Konecna, E.; Cegan, R.; Bruns, A.N.; Bisaro, D.M.; Voytas, D.F. Conferring resistance to geminiviruses with the CRISPR–Cas prokaryotic immune system. Nat. Plants 2015, 1, 15145. [Google Scholar] [CrossRef]
- Ji, X.; Zhang, H.; Zhang, Y.; Wang, Y.; Gao, C. Establishing a CRISPRCas-like immune system conferring DNA virus resistance in plants. Nat. Plants 2015, 1, 15144. [Google Scholar] [CrossRef]
- Wang, Y.; Cheng, X.; Shan, Q.; Zhang, Y.; Liu, J.; Gao, C.; Qiu, J.L. Simultaneous editing of three homoeoalleles in hexaploid bread wheat confers heritable resistance to powdery mildew. Nat. Biotechnol. 2014, 32, 947–951. [Google Scholar] [CrossRef]
- Chandrasekaran, J.; Brumin, M.; Wolf, D.; Leibman, D.; Klap, C.; Pearlsman, M.; Sherman, A.; Arazi, T.; Gal-On, A. Development of broad virus resistance in non-transgenic cucumber using CRISPR/Cas9 technology. Mol. Plant Pathol. 2016, 17, 1140–1153. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhang, C.; Liu, W.; Gao, W.; Liu, C.; Song, G.; Li, W.X.; Mao, L.; Chen, B.; Xu, Y.; et al. An alternative strategy for targeted gene replacement in plants using a dual-sgRNA/Cas9 design. Sci. Rep. 2016, 6, 23890. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, J.; Wei, P.; Zhang, B.; Gou, F.; Feng, Z.; Mao, Y.; Yang, L.; Zhang, H.; Xu, N.; et al. The CRISPR/Cas9 system produces specific and homozygous targeted gene editing in rice in one generation. Plant Biotechnol. J. 2014, 12, 797–807. [Google Scholar] [CrossRef]
- Wang, L.; Chen, L.; Li, R.; Zhao, R.; Yang, M.; Sheng, J.; Shen, L. Reduced drought tolerance by CRISPR/Cas9-mediated SlMAPK3 mutagenesis in tomato plants. J. Agric. Food Chem. 2017, 65, 8674–8682. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Gao, H.; Wang, H.; Lafitte, H.R.; Archibald, R.L.; Yang, M.; Hakimi, S.M.; Mo, H.; Habben, J.E. ARGOS8 variants generated by CRISPR-Cas9 improve maize grain yield under field drought stress conditions. Plant Biotechnol. J. 2017, 15, 207–216. [Google Scholar] [CrossRef]
- Yin, Y.; Qin, K.; Song, X.; Zhang, Q.; Zhou, Y.; Xia, X.; Yu, J. BZR1 Transcription Factor Regulates Heat Stress Tolerance through FERONIA Receptor-like Kinase-Mediated Reactive Oxygen Species Signaling in Tomato. Plant Cell Physiol. 2018, 59, 2239–2254. [Google Scholar] [CrossRef]
- Oz, M.T.; Altpeter, A.; Karan, R.; Merotto, A.; Altpeter, F. CRISPR/Cas9-Mediated Multi-Allelic Gene Targeting in Sugarcane Confers Herbicide Tolerance. Front. Genome Ed. 2021, 3, 673566. [Google Scholar] [CrossRef]
- Mao, X.; Zheng, Y.; Xiao, K.; Wei, Y.; Zhu, Y.; Cai, Q.; Chen, L.; Xie, H.; Zhang, J. OsPRX2 contributes to stomatal closure and improves potassium deficiency tolerance in rice. Biochem. Biophys. Res. Commun. 2018, 495, 461–467. [Google Scholar] [CrossRef]
- Cordones, M.N.; Mohamed, S.; Tanoi, K.; Natsuko Kobayashi, N.I.; Takagi, K.; Vernet, A.; Guiderdoni, E.; Perin, C.; Sentenac, H.; Very, A.A. Production of low-Cs + rice plants by inactivation of the K + transporter OsHAK1 with the CRISPR-Cas system. Plant J. 2017, 92, 43–56. [Google Scholar] [CrossRef]
- Xu, H.; Xiao, T.; Chen, C.H.; Li, W.; Meyer, C.A.; Wu, Q.; Wu, D.; Cong, L.; Zhang, F.; Liu, J.S.; et al. Sequence determinants of improved CRISPR sgRNA design. Genome Res. 2015, 25, 1147–1157. [Google Scholar] [CrossRef]
- Brackett, N.F.; Pomés, A.; Chapman, M.D. New Frontiers: Precise Editing of Allergen Genes Using CRISPR. Front. Allergy 2022, 17, 821107. [Google Scholar] [CrossRef]
- Liu, Q.; Yang, F.; Zhang, J.; Liu, H.; Rahman, S.; Islam, S.; Ma, W.; She, M. Application of CRISPR/Cas9 in Crop Quality Improvement. Int. J. Mol. Sci. 2021, 22, 4206. [Google Scholar] [CrossRef] [PubMed]
- Camerlengo, F.; Frittelli, A.; Sparks, C.; Doherty, A.; Martignago, D.; Larré, C.; Lupi, R.; Sestili, F.; Masci, S. CRISPR-Cas9 multiplex editing of the α-amylase/trypsin inhibitor genes to reduce allergen proteins in durum wheat. Front. Sustain. Food Syst. 2020, 4, 104. [Google Scholar] [CrossRef]
- Sugano, S.; Hirose, A.; Kanazashi, Y.; Adachi, K.; Hibara, M.; Itoh, T.; Mikami, M.; Endo, M.; Hirose, S.; Maruyama, N.; et al. Simultaneous induction of mutant alleles of two allergenic genes in soybean by using site-directed mutagenesis. BMC Plant Biol. 2020, 20, 513. [Google Scholar] [CrossRef]
- Dodo, H. SBIR Phase II: Development of an Allergen-Free Peanut Using Genome Editing Technology; National Science Foundation: Alexandria, VA, USA, 2021. [Google Scholar]
- Sarah, B. Beyond Risk Considerations: Where and How Can a Debate About Non-safety Related Issues of Genome Editing in Agriculture Take Place? Front. Plant Sci. 2018, 9, 1724. [Google Scholar] [CrossRef]
- Ayanoğlu, F.B.; Elçin, A.E.; Elçin, Y.M. Bioethical issues in genome editing by CRISPR-Cas9 technology. Turk. J. Biol. 2020, 44, 110–120. [Google Scholar] [CrossRef]
- Pellegrino, E.; Bedini, S.; Nuti, M.; Ercoli, L. Impact of genetically engineered maize on agronomic, environmental and toxicological traits: A meta-analysis of 21 years of field data. Sci. Rep. 2018, 8, 3113. [Google Scholar] [CrossRef]
- Brookes, G.; Barfoot, P. Economic impact of GM crops: The global income and production effects 1996-2012. GM Crops Food 2014, 5, 65–75. [Google Scholar] [CrossRef]
- Brookes, G. Genetically Modified (GM) Crop Use 1996-2020: Environmental Impacts Associated with Pesticide Use change. GM Crops Food 2022, 13, 262–289. [Google Scholar] [CrossRef]
- Klee, H.J.; Giovannoni, J.J. Genetics and control of tomato fruit ripening and quality attributes. Annu. Rev. Genet. 2011, 45, 41–59. [Google Scholar] [CrossRef]
- Elitzur, T.; Yakir, E.; Quansah, L.; Zhangjun, F.; Vrebalov, J.; Khayat, E.; Giovannoni, J.J.; Friedman, H. Banana MaMADS Transcription Factors Are Necessary for Fruit Ripening and Molecular Tools to Promote Shelf-Life and Food Security. Plant Physiol. 2016, 171, 380–391. [Google Scholar] [PubMed]
- Haun, W.; Coffman, A.; Clasen, B.M.; Demorest, Z.L.; Lowy, A.; Ray, E.; Retterath, A.; Stoddard, T.; Juillerat, A.; Cedrone, F.; et al. Improved soybean oil quality by targeted mutagenesis of the fatty acid desaturase 2 gene family. Plant Biotechnol. J. 2014, 12, 934–940. [Google Scholar] [CrossRef] [PubMed]
- Gianotto, A.C.; Rocha, M.S.; Cutri, L.; Lopes, F.C.; Dal’Acqua, W.; Hjelle, J.J.; Lirette, R.P.; Oliveira, W.S.; Sereno, M.L. The insect-protected CTC91087-6 sugarcane event expresses Cry1Ac protein preferentially in leaves and presents compositional equivalence to conventional sugarcane. GM Crops Food 2019, 10, 208–219. [Google Scholar] [CrossRef] [PubMed]
- Ligon, J.; Lessard, P.; Raab, R. Petition for the Determination of Nonregulated Status of Maize Event PY203; Agrivida Inc.: Woburn, MA, USA, 2019; 114p. [Google Scholar]
- Yu, X.; Sun, Y.; Lin, C.; Wang, P.; Shen, Z.; Zhao, Y. Development of Transgenic Maize Tolerant to Both Glyphosate and Glufosinate. Agronomy 2023, 13, 226. [Google Scholar] [CrossRef]
- Anderson, J.A.; Hong, B.; Moellring, E.; TeRonde, S.; Walker, C.; Wang, Y.; Maxwell, C. Composition of forage and grain from genetically modified DP202216 maize is equivalent to non-modified conventional maize (Zea mays L.). GM Crops Food 2019, 10, 77–89. [Google Scholar] [CrossRef] [PubMed]
- MacIntosh, S.C.; Shaw, M.; Connelly, M.; Yao, Z. Food and Feed Safety of NS-B5ØØ27-4 Omega-3 Canola (Brassica napus): A New Source of Long-Chain Omega-3 Fatty Acids. Front. Nutr. 2021, 8, 716659. [Google Scholar] [CrossRef]
- Ayala, F.; Fedrigo, G.V.; Burachik, M.; Miranda, P.V. Compositional equivalence of event IND-ØØ412-7 to non-transgenic wheat. Transgenic Res. 2019, 28, 165–176. [Google Scholar] [CrossRef]
- Mullins, E.; Bresson, J.; Dalmay, T.; Dewhurst, I.; Epstein, M.; Firbank, L.; Guerche, P.; Hejatko, J.; Moreno, F.J.; Naegeli, H.; et al. Assessment of genetically modified Maize MON 87429 for food and feed uses, under Regulation (EC) No 1829/2003 (application EFSA-GMO-NL-2019-161). EFSA J. 2022, 20, e07589. [Google Scholar] [CrossRef]
- Smith, B.; Zimmermann, C.; Carlson, A.; Mathesius, C.; Mukerji, P.; McNaughton, J.; Walker, C.; Roper, J. Evaluation of the safety and nutritional equivalency of maize grain with genetically modified event DP-Ø23211-2. GM Crops Food 2021, 12, 396–408. [Google Scholar] [CrossRef]
- Horikoshi, R.; Ferrari, G.; Dourado, P.; Climaco, J.; Vertuan, H.; Evans, A.; Pleau, M.; Morrell, K.; José, M.; Anderson, H.; et al. MON 95379 Bt maize as a new tool to manage sugarcane borer (Diatraea saccharalis) in South America. Pest Manag. Sci. 2022, 78, 3456–3466. [Google Scholar] [CrossRef]
- Webber, G.D. Insect-Resistant Crops through Genetic Engineering. North Central Regional Publication, University of Missouri Extension. 1995. Available online: https://mospace.umsystem.edu/xmlui/bitstream/handle/10355/7598/InsectResistantCropsGeneticEngineering.pdf?sequence=1&isAllowed=y (accessed on 12 June 2023).
- Koch, M.S.; Ward, J.M.; Levine, S.L.; Baum, G.A.; Vicini, J.L.; Hammond, B.G. The food and environmental safety of Bt crops. Front. Plant Sci. 2015, 6, 283–336. [Google Scholar] [PubMed]
- Abbas, M.S.T. Genetically engineered (modified) crops (Bacillus thuringiensis crops) and the world controversy on their safety. Egypt. J. Biol. Pest Control 2018, 28, 5. [Google Scholar]
- Brookes, G.; Barfoot, P. Global income and production impacts of using GM crop technology 1996–2014. GM Crops Food 2016, 7, 38–77. [Google Scholar] [CrossRef] [PubMed]
- ISAAA. Global Status of Commercialized Biotech/GM Crops: 2019. ISAAA Brief 55. 2019. Available online: https://www.isaaa.org/resources/publications/briefs/55/default.asp (accessed on 2 July 2023).
- Baum, J.; Bogaert, T.; Clinton, W.; Heck, G.R.; Feldmann, P.; Ilagan, O.; Johnson, S.; Plaetinck, G.; Munyikwa, T.; Pleau, M.; et al. Control of coleopteran insect pests through RNA interference. Nat. Biotechnol. 2007, 25, 1322–1326. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.B.; Cai, W.J.; Wang, J.W.; Hong, G.J.; Tao, X.Y.; Wang, L.J.; Huang, Y.P.; Chen, X.Y. Silencing a cotton bollworm P450 monooxygenase gene by plant-mediated RNAi impairs larval tolerance of gossypol. Nat. Biotechnol. 2007, 25, 1307–1313. [Google Scholar] [CrossRef]
- Jin, S.; Singh, N.D.; Li, L.; Zhang, X.; Daniell, H. Engineered chloroplast dsRNA silences cytochrome p450 monooxygenase, V-ATPase and chitin synthase genes in the insect gut and disrupts Helicoverpa armigera larval development and pupation. Plant Biotechnol. J. 2015, 13, 435–446. [Google Scholar] [CrossRef]
- Mamta, B.; Rajam, M.V. RNAi technology: A new platform for crop pest control. Physiol. Mol. Biol. Plants. 2017, 23, 487–501. [Google Scholar] [CrossRef]
- Malik, H.J.; Raza, A.; Amin, I.; Scheffler, J.A.; Scheffler, B.E.; Brown, J.K.; Mansoor, S. RNAi-mediated mortality of the whitefly through transgenic expression of double-stranded RNA homologous to acetylcholinesterase and ecdysone receptor in tobacco plants. Sci. Rep. 2016, 6, 38469. [Google Scholar] [CrossRef]
- Thakur, N.; Upadhyay, S.K.; Verma, P.C.; Chandrashekar, K.; Tuli, R.; Singh, P.K. Enhanced whitefly resistance in transgenic tobacco plants expressing double stranded RNA of v-ATPase A gene. PLoS ONE 2014, 9, e87235. [Google Scholar] [CrossRef]
- Mao, J.; Zeng, F. Plant-mediated RNAi of a gap gene-enhanced tobacco tolerance against the Myzus persicae. Transgenic Res. 2014, 23, 145–152. [Google Scholar] [CrossRef]
- Yu, R.; Xu, X.; Liang, Y.; Tian, H.; Pan, Z.; Jin, S.; Wang, N.; Zhang, W. The insect ecdysone receptor is a good potential target for RNAi-based pest control. Int. J. Biol. Sci. 2014, 10, 1171–1180. [Google Scholar] [CrossRef] [PubMed]
- Carrière, Y.; Ellers-Kirk, C.; Sisterson, M.; Antilla, L.; Whitlow, M.; Dennehy, T.J.; Tabashnik, B.E. Long-term regional suppression of pink bollworm by Bacillus thuringiensis cotton. Proc. Natl. Acad. Sci. USA 2003, 100, 1519–1523. [Google Scholar] [CrossRef] [PubMed]
- Hutchison, W.D.; Burkness, E.C.; Mitchell, P.D.; Moon, R.D.; Leslie, T.W.; Fleischer, S.J.; Abrahamson, M.; Hamilton, K.L.; Steffey, K.L.; Gray, M.E.; et al. Area wide Suppression of European Corn Borer with Bt Maize Reaps Savings to Non-Bt Maize Growers. Science 2010, 330, 222–225. [Google Scholar] [CrossRef] [PubMed]
- Dively, G.P.; Venugopal, P.D.; Bean, D.; Whalen, J.; Holmstrom, K.; Kuhar, T.P. Regional pest suppression associated with widespread Bt maize adoption benefits vegetable growers. Proc. Natl. Acad. Sci. USA 2018, 115, 3320–3325. [Google Scholar] [CrossRef]
- Wu, K.M.; Lu, Y.H.; Feng, H.Q.; Jiang, Y.Y.; Zhao, J.Z. Suppression of cotton bollworm in multiple crops in China in areas with Bt toxin-containing cotton. Science 2008, 321, 1676–1678. [Google Scholar] [CrossRef]
- National Academies of Sciences, Engineering, and Medicine. Genetically Engineered Crops: Experiences and Prospects—Stats. 2016. Available online: https://www.nap.edu/catalog/23395/genetically-engineered-crops-experiences-and-prospects (accessed on 7 July 2023).
- Achary, V.; Sheri, V.; Manna, M.; Panditi, V.; Borphukan, B.; Ram, B.; Agarwal, A.; Fartyal, D.; Teotia, D.; Masakapalli, S.K.; et al. Overexpression of improved EPSPS gene results in field level glyphosate tolerance and higher grain yield in rice. Plant Biotechnol. J. 2020, 18, 2504–2519. [Google Scholar] [CrossRef]
- Anderson, J.A.; Ellsworth, P.C.; Faria, J.C.; Head, G.P.; Owen, M.D.K.; Pilcher, C.D.; Shelton, A.M.; Meissle, M. Genetically Engineered Crops: Importance of Diversified Integrated Pest Management for Agricultural Sustainability. Front. Bioeng. Biotechnol. 2019, 7, 14. [Google Scholar]
- Dong, H.; Huang, Y.; Wang, K. The Development of Herbicide Resistance Crop Plants Using CRISPR/Cas9-Mediated Gene Editing. Genes 2021, 12, 912. [Google Scholar] [CrossRef]
- Butt, H.; Rao, G.S.; Sedeek, K.; Aman, R.; Kamel, R.; Mahfouz, M. Engineering herbicide resistance via prime editing in rice. Plant Biotechnol. J. 2020, 18, 2370–2372. [Google Scholar] [CrossRef]
- Shimatani, Z.; Kashojiya, S.; Takayama, M.; Terada, R.; Arazoe, T.; Ishii, H.; Teramura, H.; Yamamoto, T.; Komatsu, H.; Miura, K.; et al. Targeted base editing in rice and tomato using a CRISPR-Cas9 cytidine deaminase fusion. Nat. Biotechnol. 2017, 35, 441–443. [Google Scholar]
- Liu, X.; Qin, R.; Li, J.; Liao, S.; Shan, T.; Xu, R.; Wu, D.; Wei, P. A CRISPR-Cas9-mediated domain-specific base-editing screen enables functional assessment of ACCase variants in rice. Plant Biotechnol. J. 2020, 18, 1845–1847. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhu, J.; Wu, H.; Liu, C.; Huang, C.; Lan, J.; Zhao, Y.; Xie, C. Precise base editing of non-allelic acetolactate synthase genes confers sulfonylurea herbicide resistance in maize. Crop J. 2020, 8, 449–456. [Google Scholar] [CrossRef]
- Tian, S.; Jiang, L.; Cui, X.; Zhang, J.; Guo, S.; Li, M.; Zhang, H.; Ren, Y.; Gong, G.; Zong, M.; et al. Engineering herbicide-resistant watermelon variety through CRISPR/Cas9-mediated base-editing. Plant Cell. Rep. 2018, 37, 1353–1356. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Chen, C.; Xian, G.; Liu, D.; Lin, L.; Yin, S.; Sun, Q.; Fang, Y.; Zhang, H.; Wang, Y.; et al. Engineering herbicide-resistant oilseed rape by CRISPR/Cas9-mediated cytosine base-editing. Plant Biotechnol. J. 2020, 18, 1857–1859. [Google Scholar] [CrossRef]
- Wang, Z.; Wan, L.; Xin, Q.; Zhang, X.; Song, Y.; Wang, P.; Hong, D.; Fan, Z.; Yang, G. Optimising glyphosate tolerance in rapeseed (Brassica napus L.) by CRISPR/Cas9-based geminiviral donor DNA replicon system with Csy4-based single-guide RNA processing. J. Exp. Bot. Bot. 2021, 72, 4796–4808. [Google Scholar] [CrossRef]
- Li, C.; Zong, Y.; Wang, Y.; Jin, S.; Zhang, D.; Song, Q.; Zhang, R.; Gao, C. Expanded base editing in rice and wheat using a Cas9-adenosine deaminase fusion. Genome Biol. 2018, 19, 59. [Google Scholar] [CrossRef]
- Bouis, H.E.; Hotz, C.; McClafferty, B.; Meenakshi, J.V.; Pfeiffer, W.H. Biofortification: A new tool to reduce micronutrient malnutrition. Food Nutr. Bull. 2011, 32, S31–S40. [Google Scholar]
- Al-Babili, S.; Beyer, P. Golden Rice Five years on the road Five years to go? Trends Plant Sci. 2005, 10, 565–573. [Google Scholar] [CrossRef]
- Hefferon, K.L. Nutritionally enhanced food crops; progress and perspectives. Int. J. Mol. Sci. 2015, 16, 3895–3914. [Google Scholar] [CrossRef]
- Pérez-Massot, E.; Banakar, R.; Gómez-Galera, S.; Zorrilla-López, U.; Sanahuja, G.; Arjó, G.; Miralpeix, B.; Vamvaka, E.; Farré, G.; Rivera, S.M. The contribution of transgenic plants to better health through improved nutrition: Opportunities and constraints. Genes. Nutr. 2013, 8, 29–41. [Google Scholar] [CrossRef]
- Rajaeieh, G.; Takian, A.; Kalantari, N.; Mohammadi-Nasrabadi, F. Analysis for policy to overcome barriers to reducing the prevalence of vitamin a deficiency among children (15–23 months) in Iran. BMC Public Health 2021, 21, 1234. [Google Scholar] [CrossRef] [PubMed]
- Tang, G.; Qin, J.; Dolnikowski, G.G.; Russell, R.M.; Grusak, M.A. Golden Rice is an effective source of vitamin A. Am. J. Clin. Nutr. 2009, 89, 1776–1783. [Google Scholar] [CrossRef] [PubMed]
- Howe, J.A.; Tanumihardjo, S.A. Carotenoid-biofortified maize maintains adequate vitamin a status in Mongolian gerbils. J. Nutr. 2006, 136, 2562–2567. [Google Scholar] [CrossRef]
- Banakar, R.; Alvarez Fernandez, A.; Díaz-Benito, P.; Abadia, J.; Capell, T.; Christou, P. Phytosiderophores determine thresholds for iron and zinc accumulation in biofortified rice endosperm while inhibiting the accumulation of cadmium. J. Exp. Bot. 2017, 68, 4983–4995. [Google Scholar] [CrossRef]
- Matres, J.M.; Arcillas, E.; Cueto-Reaño, M.F.; Sallan-Gonzales, R.; Trijatmiko, K.R.; Slamet-Loedin, I. Biofortification of Rice Grains for Increased Iron Content. In Rice Improvement; Ali, J., Wani, S.H., Eds.; Springer: Cham, Switzerland, 2016. [Google Scholar] [CrossRef]
- Senguttuvel, P.; Padmavathi, G.; Jasmine, C.; Sanjeeva Rao, D.; Neeraja, C.N.; Jaldhani, V.; Beulah, P.; Gobinath, R.; Aravind Kumar, J.; Sai Prasad, S.V.; et al. Rice biofortification: Breeding and genomic approaches for genetic enhancement of grain zinc and iron contents. Front. Plant Sci. 2023, 14, 1138408. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.Q.; Yu, W.H.; Wu, H.Y.; Zhang, C.Q.; Sun, S.S.; Liu, Q.Q. Lysine biofortification in rice by modulating feedback inhibition of aspartate kinase and dihydrodipicolinate synthase. Plant Biotechnol. J. 2021, 19, 490–501. [Google Scholar] [CrossRef]
- Mugode, L.; Há, B.; Kaunda, A.; Sikombe, T.; Phiri, S.; Mutale, R.; Davis, C.; Tanumihardjo, S.; de Moura, F.F. Carotenoid retention of biofortified provitamin a maize (Zea mays L.) after Zambian traditional methods of milling, cooking and storage. J. Agric. Food Chem. 2014, 62, 6317–6325. [Google Scholar] [CrossRef]
- Jeong, J.; Guerinot, M.L. Biofortified and bioavailable: The gold standard for plant-based diets. Proc. Natl. Acad. Sci. USA 2008, 105, 1777–1778. [Google Scholar] [CrossRef]
- Patel, P.; Patel, R.; Patel, S.; Patel, Y.; Patel, M.; Trivedi, R. Edible Vaccines: A Nutritional Substitute for Traditional Immunization. Pharmacogn. Rev. 2022, 16, 62–69. [Google Scholar] [CrossRef]
- Stander, J.; Mbewana, S.; Meyers, A.E. Plant-derived human vaccines: Recent developments. Biol. Drugs 2022, 36, 573–589. [Google Scholar] [CrossRef]
- Qureshi, M.B. Arif, S.; Rathore, S.S. Edible Plant Vaccines: A Step Towards Revolution in the Field of Immunology. Int. J. Agric. Biol. 2023, 29, 361–369. [Google Scholar]
- Phoolcharoen, W.; Dye, J.M.; Kilbourne, J.; Piensook, K.; Pratt, W.D.; Arntzen, C.J.; Chen, Q.; Mason, H.S.; Herbst-Kralovetz, M.M. A nonreplicating subunit vaccine protects mice against lethal Ebola virus challenge. Proc. Natl. Acad. Sci. USA 2011, 108, 20695–20700. [Google Scholar] [CrossRef] [PubMed]
- Thanavala, Y.; Mahoney, M.; Pal, S.; Scott, A.; Richter, L.; Natarajan, N.; Goodwin, P.; Arntzen, C.J.; Mason, H.S. Immunogenicity in humans of an edible vaccine for hepatitis B. Proc. Natl. Acad. Sci. USA 2005, 102, 3378–3382. [Google Scholar] [CrossRef] [PubMed]
- Mason, H.S.; Haq, T.A.; Clements, J.D.; Arntzen, C.J. Edible vaccine protects mice against Escherichia coli heat-labile enterotoxin (LT): Potatoes expressing a synthetic LT-B gene. Vaccine 1998, 16, 1336–1343. [Google Scholar] [CrossRef]
- Mason, H.S.; Ball, J.M.; Shi, J.J.; Jiang, X.; Estes, M.K.; Arntzen, C.J. Expression of Norwalk virus capsid protein in transgenic tobacco and potato and its oral immunogenicity in mice. Proc. Natl. Acad. Sci. USA 1996, 93, 5335–5340. [Google Scholar] [CrossRef]
- Chowdhury, K.; Bagasra, O. An edible vaccine for malaria using transgenic tomatoes of varying sizes, shapes and colors to carry different antigens. Med. Hypotheses 2007, 68, 22–30. [Google Scholar] [CrossRef]
- Abdurakhmonov, I.; Buriev, Z.; Shermatov, S.; Usmanov, D.; Mirzakhmedov, M.; Ubaydullaeva, K.; Kamburova, V.; Rakhmanov, B.; Ayubov, M.; Abdullaev, A.; et al. The edible tomato COVID-19 vaccine, TOMAVAC, induces neutralising IgGs. Researchsquare 2023. [Google Scholar] [CrossRef]
- Oszvald, M.; Kang, T.J.; Tomoskozi, S.; Tamas, C.; Tamas, L.; Kim, T.G.; Yang, M.S. Expression of a synthetic neutralizing epitope of porcine epidemic diarrhea virus fused with synthetic B subunit of Escherichia coli heat labile enterotoxin in rice endosperm. Mol. Biotechnol. 2007, 35, 215–223. [Google Scholar] [CrossRef]
- Huy, N.X.; Kim, S.H.; Yang, M.S.; Kim, T.G. Immunogenicity of a neutralizing epitope from porcine epidemic diarrhea virus: M cell targeting ligand fusion protein expressed in transgenic rice calli. Plant Cell Rep. 2012, 31, 1933–1942. [Google Scholar] [CrossRef]
- Kumar, G.B.; Ganapathi, T.R.; Revathi, C.J.; Srinivas, L.; Bapat, V.A. Expression of hepatitis B surface antigen in transgenic banana plants. Planta 2005, 222, 484–493. [Google Scholar] [CrossRef]
- Kim, T.G.; Kim, M.Y.; Kim, B.G.; Kang, T.J.; Kim, Y.S.; Jang, Y.S.; Arntzen, C.J.; Yang, M.S. Synthesis and assembly of Escherichia coli heat-labile enterotoxin B subunit in transgenic lettuce (Lactuca sativa). Protein Expr. Purif. 2007, 51, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Huy, N.X.; Yang, M.S.; Kim, T.G. Expression of a cholera toxin B subunit-neutralizing epitope of the porcine epidemic diarrhea virus fusion gene in transgenic lettuce (Lactuca sativa L.). Mol. Biotechnol. 2011, 48, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Karasev, A.V.; Foulke, S.; Wellens, C.; Rich, A.; Shon, K.J.; Zwierzynski, I.; Hone, D.; Koprowski, H.; Reitz, M. Plant based HIV-1 vaccine candidate: Tat protein produced in spinach. Vaccine 2005, 23, 1875–1880. [Google Scholar] [CrossRef] [PubMed]
- Hahn, B.S.; Jeon, I.S.; Jung, Y.J.; Kim, J.B.; Park, J.S.; Ha, S.H.; Kim, K.H.; Kim, H.M.; Yang, J.S.; Kim, Y.H. Expression of hemagglutinin-neuraminidase protein of Newcastle disease virus in transgenic tobacco. Plant Biotechnol. Rep. 2007, 1, 85–92. [Google Scholar] [CrossRef]
- Tacket, C.O. Plant-based vaccines against diarrheal diseases. Trans. Am. Clin. Climatol. Assoc. 2007, 118, 79–87. [Google Scholar]
- Issaro, N.; Wang, D.; Liu, M.; Tassaneetrithep, B.; Phawong, C.; Rattanarojpong, T.; Jiang, C. Transgenic carrot plant-made edible vaccines against human infectious diseases. J. Innov. Pharm. Biol. Sci. 2018, 5, 43–48. [Google Scholar]
- Mishra, N.; Gupta, P.N.; Khatri, K.; Goyal, A.K.; Vyas, S.P. Edible vaccines: A new approach to oral immunization. Ind. J. Biotechnol. 2008, 7, 283–294. [Google Scholar]
- Kurup, V.M.; Thomas, J. Edible Vaccines: Promises and Challenges. Mol. Biotechnol. 2020, 62, 79–90. [Google Scholar] [CrossRef]
- Kapusta, J.; Modelska, A.; Figlerowicz, M.; Pniewski, T.; Letellier, M.; Lisowa, O.; Yusibov, V.; Koprowski, H.; Plucienniczak, A.; Legocki, A.B. A plant-derived edible vaccine against hepatitis B virus. FASEB J. 1999, 13, 1796–1799, Erratum in FASEB J. 1999, 13, 2339. [Google Scholar] [CrossRef]
- Tacket, C.O.; Mason, H.S.; Losonsky, G.; Estes, M.K.; Levine, M.M.; Arntzen, C.J. Human immune responses to a novel norwalk virus vaccine delivered in transgenic potatoes. J. Infect. Dis. 2000, 182, 302–305. [Google Scholar] [CrossRef]
- Kurokawa, S.; Nakamura, R.; Mejima, M.; Kozuka-Hata, H.; Kuroda, M.; Takeyama, N.; Oyama, M.; Satoh, S.; Kiyono, H.; Masumura, T.; et al. MucoRice-cholera toxin B-subunit, a rice-based oral cholera vaccine, down-regulates the expression of a-amylase/trypsin inhibitor-like protein family as major rice allergens. J. Proteome Res. 2013, 12, 3372–3382. [Google Scholar] [CrossRef] [PubMed]
- ISAAA. ISAAA’s GM Approval Database. 2022. Available online: www.isaaa.org/gmapprovaldatabase (accessed on 28 December 2022).
- James, C. 20th Anniversary (1996 to 2015) of the Global Commercialization of Biotech Crops and Biotech Crop Highlights in 2015; ISAAA Brief. No. 51; ISAAA: Ithaca, NY, USA, 2015; ISBN 978-1-892456-65-6. [Google Scholar]
- Ichim, M.C. The more favorable attitude of the citizens toward GMOs supports a new regulatory framework in the European Union. GM Crops Food 2021, 12, 18–24. [Google Scholar] [CrossRef]
- Rabbanee, F.K.; Afroz, T.; Naser, M.M. Are consumers loyal to genetically modified food? Evidence from Australia. Brit. Food J. 2020, 123, 803–819. [Google Scholar] [CrossRef]
- Kuzma, J.; Kokotovich, A. Renegotiating GM crop regulation: Targeted gene-modification technology raises new issues for the oversight of genetically modified crops. EMBO Rep. 2011, 12, 883–888. [Google Scholar] [CrossRef] [PubMed]
- Sprink, T.; Eriksson, D.; Schiemann, J.; Hartung, F. Regulatory hurdles for genome editing: Process- vs. product-based approaches in different regulatory contexts. Plant Cell Rep. 2016, 35, 1493–1506. [Google Scholar] [CrossRef] [PubMed]
- Nerkar, G.; Devarumath, S.; Purankar, M.; Kumar, A.; Valarmathi, R.; Devarumath, R.; Appunu, C. Advances in Crop Breeding Through Precision Genome Editing. Front. Genet. 2022, 13, 880195. [Google Scholar] [CrossRef]
- Lassoued, R.; Phillips, P.W.P.; Macall, D.M.; Hesseln, H.; Smyth, S.J. Expert opinions on the regulation of plant genome editing. Plant Biotechnol. J. 2021, 19, 1104–1109. [Google Scholar] [CrossRef]
- Voigt, B. EU regulation of gene-edited plants—A reform proposal. Front. Genome Ed. 2023, 14, 1119442. [Google Scholar] [CrossRef]
- Escajedo San-Epifanio, L.; Filibi, I.; Lasa López, A.; Puigdomènech, P.; Uncetabarrenechea Larrabe, J. Possible EU futures for CRISPR-edited plants: Little margin for optimism? Front. Plant Sci. 2023, 14, 1141455. [Google Scholar] [CrossRef]
- Zimny, T. Regulation of GMO field trials in the EU and new genomic techniques: Will the planned reform facilitate experimenting with gene-edited plants? BioTechnologia 2023, 104, 75–83. [Google Scholar] [CrossRef]
- Davison, J.; Ammann, K. New GMO regulations for old: Determining a new future for EU crop biotechnology. GM Crops Food 2017, 8, 13–34. [Google Scholar] [CrossRef] [PubMed]
- Sven, W.; Jacobsen, E. Feeding the world. Genetically Modified Crops Versus Agricultural Biodiversity. Agron. Sustain. Dev. 2013, 33, 651–662. [Google Scholar]
- ISAAA. Executive Summary Global Status of Commercialized Biotech/GM Crops; Brief 46; ISAAA: Ithaca, NY, USA, 2013. [Google Scholar]
- Young, S. GMO and the Nutritional Content of Food. Discovery Eye Foundation. 2015. Available online: https://discoveryeye.org/gmo-and-nutritional-content-of-food/ (accessed on 28 June 2023).
- Qaim, M.; Kouser, S. Genetically modified crops and food security. PLoS ONE 2013, 8, e64879. [Google Scholar] [CrossRef] [PubMed]
- Brookes, G. Crop Biotechnology Continues to Provide Higher Farmer Income and Significant Environmental Benefits. PG Economics. 2020. Available online: https://pgeconomics.co.uk/press+releases/25/ (accessed on 11 June 2023).
- Gruère, G.; Sengupta, D. Bt Cotton and farmer suicides in India: An evidence-based assessment. J. Dev. Stud. 2011, 47, 316–337. [Google Scholar] [CrossRef]
- Smyth, S.J. The human health benefits from GM crops. Plant Biotechnol. J. 2020, 18, 887–888. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait Modified | Crop | Target Gene(s) | Gene(s) Role | References | |
|---|---|---|---|---|---|
| Yield parameters | Fruit size | Tomato | CLV3 | Shoot and floral meristem development | [59] |
| ENO | Encodes enolase, which catalyzes the conversion of 2-phosphoglycerate to phosphoenolpyruvate | [55] | |||
| fas, lc | Determines locule number | [60] | |||
| Fas, OVATE | Fruit shape and size | [59] | |||
| Groundcherry | ClV1 | Controls shoot and floral meristem size | [61] | ||
| Grain size | Wheat | TaGW2 | Determines grain weight | [50] | |
| Rice | GW5 | Grain width and weight | [42] | ||
| OsGS3, OsGn1a, and OsGW2 | Grain size | [62] | |||
| GS3, Gn1a | Controls grain number, size, and density of erect panicles | [63] | |||
| TGW6, GW2, GW5 | Grain length | ||||
| Grain shape | Wheat | TaGW7 | Cell division and organ growth | [51] | |
| Rice | GS9 | Cell division and grain development | [64] | ||
| Grain color | Rice | ANT1 | Controls anthocyanin pigmentation in different vegetative tissues | [65] | |
| CRTISO | Catalyzes prolycopene to lycopene | [66] | |||
| Psy1, CrtR-b2 | Influences carotenoid accumulation | [53] | |||
| SlMYB12 | Flavonol biosynthesis | [67] | |||
| Maize | Psy1 | Phytoene synthesis | [52] | ||
| Floral color | Ipomoea nil | CCD | Synthesis of apocarotenoid flavor and aroma volatiles | [68] | |
| Petunia | F3H | Flavonoid metabolism | [69] | ||
| Nutrient Quality | Increased carotenoid content | Rice | ZmPsy, SSU-crtI | Carotenoid biosynthetic genes | [48] |
| Reduced amylose content | Sweet potato | IbGBSSI | Starch metabolism | [54] | |
| Cassava | PTST1, GBSS | Amylose biosynthesis | [56] | ||
| Rice | OsGBSSI | Amylopectin and amylose synthesis | [49] | ||
| Potato | StGBSS | Amylose biosynthesis | [58] | ||
| Barley | HvGBSSIa | Starch metabolism | [70] | ||
| Increased amylose content | Rice | OsBEI and OsBEIIb | Amylose biosynthesis | [71] | |
| Potato | StSBE1, StSBE2 | Amylose biosynthesis | [57] | ||
| Sweet potato | IbGBSSI, IbSBEII | Amylopectin and amylose synthesis | [54] | ||
| Increased oleic acid content | Rice | OsFAD2-1 | Controls oleic acid content | [72] | |
| Tomato | BnFAD2 | Oleic acid regulation | [73] | ||
| Camelina | CsFAD2 | Synthesis of linoleic acid from oleic acid | [74] | ||
| Reduced Phytic acid | Rapeseed | BnITPK | Critical in the phytic acid pathway | [75] | |
| Rice | OsPLDα1 | Phytic acid biosynthesis | [76] | ||
| Reduced gluten content | Wheat | α-gliadin genes | Gluten production | [77] | |
| Increased Gamma-Aminobutyric Acid (GABA) content | Rice | SlGAD2, SlGAD3 | Regulates γ-aminobutyric acid levels | [78] | |
| OsGAD3 | Gamma-aminobutyric acid synthesis | [79] | |||
| Biotic stress | Blast Resistance | Rice | OsERF922 | Negatively regulates disease resistance in rice | [80] |
| Bacterial blight | OsSWEET13 | Sugar transport system | [81] | ||
| OsSWEET11, OsSWEET13, and OsSWEET14 | Resistance against Xanthomonas | [82] | |||
| Powdery mildew | Tomato | Pmr4 | Callose synthase | [83] | |
| Bacterial speck disease | SlJAZ2 | Stomatal reopening by cor | [84] | ||
| Brown streak disease | Cassava | nCBP-1, nCBP-2 | Protein synthesis | [85] | |
| Bean yellow dwarf virus (BeYDV) resistance | N. benthamiana | BeYDV | Protein synthesis | [86] | |
| Virus resistance | dsDNA of virus (A7, B7, and C3 regions) | Protein synthesis | [87] | ||
| Powdery mildew resistance | Bread wheat | TaMLO-A1, TaMLO-B1, and TaMLOD1 | Protein synthesis | [88] | |
| Ipomovirus | Cucumber | eIF4E | mRNA translation initiation | [89] | |
| Abiotic stress | Drought tolerance | A. thaliana | MIR169a | Regulates translation of target genes | [90] |
| UGT79B2, UGT79B3 | Catalyzes the transfer of a glycosyl moiety | ||||
| Rice | OsDERF1, OsPMS3, OsEPSPS, OsMSH1, OsMYB5 | Critical in plant development | [91] | ||
| Tomato | SlMAPK3 | Regulates plants’ response to salt stress | [92] | ||
| Maize | ARGOS8 | Negative regulator of ethylene responses | [93] | ||
| Heat stress | Tomato | BZR1 | Regulates brassinosteroid response | [94] | |
| Herbicide tolerance | Soybean | ALS1 | Branched-chain amino acid synthesis | [46] | |
| Sugarcane | ALS | Leucine, isoleucine, and valine biosynthetic pathways | [95] | ||
| Potassium deficiency tolerance | Rice | OsPRX2 | Regulates plants’ response to k+ deficiency | [96] | |
| Low cesium accumulation | OsHAK-1 | Mediator of cs uptake, critical in potassium-mediated sugar metabolism | [97] | ||
| Various abiotic stress tolerance | OsAOX1a, OsAOX1b, OsAOX1c, OsBEL | Regulates plants’ response to stress | [98] |
| S/N | Plant | Identifier | Introduced Gene(s) | Gene Product(s) | Source(s) | Acquired Trait(s) | Method of Trait Introduction | Developer | Reference |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Soybean | FAD2KO | Mutated FAD2-1A and FAD2-1B | Changed oil composition | Glycine max | Increased oleic acid content of oil | Agrobacterium-mediated plant transformation; TALENs | Calyxt, Inc., Minnesota, USA | [111] |
| 2 | Fuji apple | OKA-NBØØ3-1 | PGAS PPO suppression gene | double-stranded RNA (dsRNA) | Malus domestica | Reduced polyphenol oxidase activity; non-browning | A. tumefaciens-mediated plant transformation | Okanagan Specialty Fruits, Inc., Summerland, British Columbia | |
| 3 | Cotton | TAM-66274-5 | dCS | dsRNA that suppresses the expression of endogenous d-cadinene synthase gene | Gossypium hirsutum L. | Low gossypol | A. tumefaciens-mediated plant transformation | Texas A&M AgriLife Research, Texas, USA | |
| 4 | Sugar from sugarcane | CTC91Ø87-6 | Cry1Ac | Cry1Ac delta-endotoxin | Bacillus thuringiensis subsp. Kurstaki strain HD73 | Lepidopteran insect resistance | NA | Centro de Tecnologia Canavieira, São Paulo, Brazil | [112] |
| 5 | Maize | AGV-PY203-4 | phy02 | Express the phytase enzyme (Phy02) | Escherichia coli | Phytase production | A. tumefaciens-mediated plant transformation | Agrivida, Inc., Massachusetts, United States | [113] |
| 6 | Maize | DBN-Ø9936-2 | cry1Ab; epsps (Ag) | Cry1Ab delta-endotoxin; 5-enolpyruvylshikimate-3-phosphate-synthase enzyme | Bacillus thuringiensis subsp. Kurstaki; Arthrobacter globiformis | Glyphosate herbicide tolerance, lepidopteran insect resistance | A. tumefaciens-mediated plant transformation | Beijing DaBeiNong Biotechnology Co. Ltd. (DBNBC), Beijing, China | [114] |
| 7 | Maize | DP-2Ø2216-6 | zmm28; mo-pat | transcription factor (ZMM28); phosphinothricin acetyltransferase (PAT) | Zea mays; Streptomyces viridochromogenes | Glufosinate herbicide tolerance, enhanced photosynthesis/yield | A. tumefaciens-mediated plant transformation | Pioneer Hi-Bred International, Inc., Iowa, USA; Dow AgroSciences LLC, Indiana, USA. | [115] |
| 8 | Potato | SPS-ØØØZ6-5 | asn1; ppo5; PhL; Vlnv; Rpi-vnt1 | double-stranded RNA; late blight resistance protein | Solanum tuberosum; Solanum venturii | Lowered free asparagine, reduced black spot, lowered reducing sugars, foliar late blight resistance | A. tumefaciens-mediated plant transformation | J.R. Simplot Company, Idaho, USA | |
| 9 | Maize | DBN-Ø9858-5 | epsps (Ag); pat | 5-enolpyruvylshikimate-3-phosphate-synthase enzyme; phosphinothricin N-acetyltransferase (PAT) enzyme | Arthrobacter globiformis; Streptomyces viridochromogenes | Glyphosate herbicide tolerance, glufosinate herbicide tolerance | Beijing DaBeiNong Biotechnology Co. Ltd. (DBNBC), Beijing, China. | ||
| 10 | Canola | NS-B5ØØ27-4 | Lackl-delta12D; Picpa-omega-3D; Micpu-delta-6D; Pyrco-delta-6E; Pavsa-delta-5D; Pyrco-delta-5E; Pavsa-delta-4D and pat | Fatty acid desaturases (delta-12, omega-3/delta-15, delta-6, delta-5, and delta-4) and phosphinothricin N-acetyltransferase (PAT) enzyme | Lachancea kluyveri; Pichia pastoris; Micromonas pusilla; Pyramimonas cordata; Pavlova salina and Streptomyces viridochromogenes | Modified oil/fatty acid; glufosinate herbicide tolerance | A. tumefaciens-mediated plant transformation | Nuseed Americas Inc., California, USA | [116] |
| 11 | Canola | BPS-BFLFK-2 | PsD12D; OtD6D; TcD5D; TcD4D, PlD4D; PiO3D, PirO3D; PpD6E; TpD6E; OtD5E; AtAHAS | Fatty acid desaturases (delta-12; delta-6; delta-5; delta-4, and omega-3); Fatty acid elongases (delta-6 and delta-5); Large catalytic subunit of acetoxhydroxyacid synthase (At-AHAS-L) | Phytophthora sojae; Ostreococcus tauri; Thraustochytrium sp.; Pavlova lutheri; Phytophthora infestans; Pythium irregulare; Physcomitrella patens; Thalassiosira pseudonana; Ostreococcus tauri; Arabidopsis thaliana | Modified oil/fatty acid, imazamox herbicide tolerance | A. tumefaciens-mediated plant transformation | BASF Plant Science, NC, USA | |
| 12 | Soybean | BCS-GM151-6 | cry14Ab-1.b; hppdPf4Pa | Cry14Ab1 protein; modified 4-hydroxyphenylpyruvate dioxygenase (HPPD-4) enzyme | Bacillus thuringiensis; Pseudomonas fluorescens strain A32 | Nematode resistance, tolerance to hppd-inhibiting herbicides | A. tumefaciens-mediated plant transformation | BASF Corporation, TX, USA | |
| 13 | Wheat | IND-ØØ412-7 | Hahb-4 | Isolated nucleic acid molecule encoding the transcription factor Hahb-4 | Helianthus annuus | Drought stress tolerance | Microparticle bombardment of plant cells or tissue | Bioceres Inc., Sante Fe, Argentina | [117] |
| 14 | Maize | MON-87429-9 | pat; dmo; cp4 epsps (aroA:CP4); ft_t | phosphinothricin N-acetyltransferase (PAT) enzyme; dicamba mono-oxygenase enzyme; herbicide tolerant form of 5-enolpyruvulshikimate-3-phosphate synthase (EPSPS) enzyme; 2,4-D and FOPs dioxygenase protein (FT_T) | Streptomyces viridochromogenes; Stenotrophomonas maltophilia strain DI-6; Agrobacterium tumefaciens strain CP4; Sphingobium herbicidovorans | Glufosinate herbicide tolerance; dicamba herbicide tolerance; glyphosate herbicide tolerance; 2,4-d herbicide tolerance | Agrobacterium tumefaciens-mediated plant transformation | Bayer CropScience LP, Australia | [118] |
| 15 | Maize | DP-Ø23211-2 | IPD072Aa; pat; pmi | IPD072Aa protein; Phosphinothricin N-acetyltransferase (PAT protein); Phosphomannose isomerase (PMI protein) | Pseudomonas chlororaphis; Streptomyces viridochromogenes; Escherichia coli | Insect resistance; Glufosinateherbicide tolerance | NA | Pioneer Hi-Bred International, Inc., Iowa, USA | [119] |
| 16 | Canola | MON-941ØØ-2 | dmo | dicamba mono-oxygenase enzyme | Stenotrophomonas maltophilia strain DI-6 | Dicamba herbicide tolerance | NA | Bayer CropScience LP, Australia | |
| 17 | Maize | MON-95379-3 | Cry1B; Cry1Da | Cry1B.868; Cry1Da_7 | Bacillus thuringiensis | Insect resistance | NA | Bayer CropScience LP, Australia | [120] |
| Plant | Transgenic Product | Targeted Disease | Administration Method | References |
|---|---|---|---|---|
| Solanum tuberosum | HBsAg | Hepatitis B | Oral | [167] |
| E. coli LT-B | Enteritis | Oral | [168] | |
| Norwalk virus cp | Diarrhea | Oral | [169] | |
| Solanum lycopersicum | CSP, MSP1 | Malaria | Oral | [170] |
| SI protein of SARS-CoV-2 | COVID-19 | Oral | [171] | |
| Oryza sativa | Coli LTB and synthetic COE of PEDV | Enteritis | Oral | [172] |
| sCOE-CO1 | Enteritis | Oral | [173] | |
| Musa sp. | HBsAg | Hepatitis B | Oral | [174] |
| Lettuce sativa | Synthetic LTB | Enteritis | Oral | [175] |
| sCTB-sCOE | Diarrhea | Oral | [176] | |
| Spinacea oleraceae | HIV-Tat | HIV-1 | Oral | [177] |
| Nicotiana tabacum | Newcastle disease virus | Newcastle disease | Subcutaneous | [178] |
| Zea mays | ETEC | Diarrhea | Oral | [179] |
| Daucus carota | Taliglucerasealfa | Gaucherdisease | Intramuscular | [180] |
| Country | Cultivation Area (Million Hec.) | % of GM Crops in Total Crops Cultivated | GM Crops |
|---|---|---|---|
| USA | 70.9 | 46.13 | maize, soybean, cotton, canola, sugarbeet, alfalfa, papaya, squash, potato |
| Brazil | 44.2 | 61.49 | soybean, maize, cotton |
| Argentina | 24.5 | 64.41 | soybean, maize, cotton |
| India | 11.6 | 7.39 | cotton |
| Canada | 11 | 25.74 | canola, maize, soybean, sugarbeet |
| China | 3.7 | 3.51 | cotton, papaya, poplar |
| Paraguay | 3.6 | 83.9 | soybean, maize, cotton |
| Pakistan | 2.9 | 13.63 | cotton |
| South Africa | 2.3 | 19.13 | maize, soybean, cotton |
| Uruguay | 1.4 | 79.2 | soybean, maize |
| Bolivia | 1.1 | 28.21 | soybean |
| Philippines | 0.7 | 1.47 | maize |
| Australia | 0.7 | 12.9 | cotton, canola |
| Burkina Faso | 0.4 | 7.2 | cotton |
| Myanmar | 0.3 | 2.78 | cotton |
| Mexico | 0.1 | 0.61 | cotton, soybean |
| Spain | 0.1 | 0.41 | maize |
| Columbia | 0.1 | 0.86 | cotton, maize |
| Sudan | 0.1 | 6.12 | cotton |
| Honduras | <0.1 | 2.65 | maize |
| Chile | <0.1 | 0.79 | maize, soybean, canola |
| Portugal | <0.1 | 11.61 | maize |
| Vietnam | <0.1 | 0.05 | maize |
| Czech Republic | <0.1 | 0.03 | maize |
| Slovakia | <0.1 | 0.01 | maize |
| Costa Rica | <0.1 | 0.04 | cotton, soybean |
| Bangladesh | <0.1 | 0 | eggplant |
| Romania | <0.1 | 0 | maize |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Animasaun, D.A.; Adedibu, P.A.; Shkryl, Y.; Emmanuel, F.O.; Tekutyeva, L.; Balabanova, L. Modern Plant Biotechnology: An Antidote against Global Food Insecurity. Agronomy 2023, 13, 2038. https://doi.org/10.3390/agronomy13082038
Animasaun DA, Adedibu PA, Shkryl Y, Emmanuel FO, Tekutyeva L, Balabanova L. Modern Plant Biotechnology: An Antidote against Global Food Insecurity. Agronomy. 2023; 13(8):2038. https://doi.org/10.3390/agronomy13082038
Chicago/Turabian StyleAnimasaun, David Adedayo, Peter Adeolu Adedibu, Yury Shkryl, Faith Olatayo Emmanuel, Liudmila Tekutyeva, and Larissa Balabanova. 2023. "Modern Plant Biotechnology: An Antidote against Global Food Insecurity" Agronomy 13, no. 8: 2038. https://doi.org/10.3390/agronomy13082038
APA StyleAnimasaun, D. A., Adedibu, P. A., Shkryl, Y., Emmanuel, F. O., Tekutyeva, L., & Balabanova, L. (2023). Modern Plant Biotechnology: An Antidote against Global Food Insecurity. Agronomy, 13(8), 2038. https://doi.org/10.3390/agronomy13082038