1. Introduction
Phosphorus (P) is a limiting factor for plant production on steep slopes in the humid tropics, where many weathered volcanic ash soils (Andosols) are located [
1]. Low P availability can often be crop yield-limiting; and it is difficult to mitigate, because of P fixation by iron- and aluminum-oxides [
2]. In addition, many parts of the world have inadequate access to fertilizers, because sources of high-grade P for fertilizer processing are limited and/or fertilizers are not economically feasible for agricultural producers [
3].
Management strategies, such as the development of P-efficient genotypes, may help improve crop production. For example, the common bean (
Phaseolus vulgaris L.) is the most important dietary legume in Latin America, because it contains up to 25% protein, in addition to its large fiber and complex carbohydrate content [
4]. However, bean yields continue to be low in Central America, due to poor soil P availability [
5]. Bean genotypes that can efficiently use soil P reserves have played a significant role in improving food security in this region [
6]. Lynch [
7] found that efficient bean genotypes have greater root mass, root length and root/shoot ratio, and their roots demonstrated an extraordinary ability to sense and respond to localized changes in P availability. To date, the bean breeding program has focused on improving this crop’s root architecture for enhancing P uptake [
8]. Henry
et al. [
6] suggested that traits that increase P acquisition may also result in an increase in bean shoot biomass and yield, in addition to a greater concentration of P in shoot tissue.
The addition of manure, compost or plant biomass (mulch) in a cut-and-carry system is an agroecosystem management strategy commonly used in tropical regions to help maintain soil fertility and improve P availability for crop growth [
9]. In a cut-and-carry system, defined as the transfer of biomass from external sources to the area of crop production, mulch is strategically located on the soil surface. The mulch biomass acts as a soil protector by minimizing erosion and as a soil amendment by enhancing soil fertility and crop yield [
10]. Biomass for cut-and-carry systems is typically obtained from perennial trees or legumes that are capable of acquiring a large fraction of P from relatively less available forms of soil P and are capable of accumulating a greater P concentration in leaves [
11]. Palm
et al. [
12] suggested that some biomass materials with a greater P (>2.5 mg P g
−1) concentration have the potential to increase P availability in soils. The Mexican sunflower [
Tithonia diversifolia (Hemsl.) Gray] may be a suitable shrub species for cut-and-carry systems, because of its greater foliar nutrient content [
13]. In Ghana, Partey
et al. [
14] observed that
T. diversifolia leaves had the greatest rate of decomposition and nutrient release rates when compared to four other leguminous species [
Senna spectabilis (DC) H. Irwin & Barnaby,
Gliricidia sepium (Jacq.) Walp.,
Laucaena leucacephala (Lam.) De Wit and
Acacia auriculiformis Benth.] used as mulch biomass in agroforestry systems.
For the long-term sustainability and assurance of food and soil security, the addition of mulch biomass in combination with mineral P fertilizer or manure may be the most optimal fertilization strategy [
12]. To date, most studies have evaluated the response of
T. diversifolia mulch biomass addition on soil fertility and crop production [
15,
16,
17]. Some studies have also evaluated the influence of
T. diversifolia mulch biomass on bean genotype production on a P-limited soil [
18]. However, no study has evaluated the response of bean genotypes to the application of leguminous and non-leguminous mulch biomass in a P-limited soil. The objectives of this study were to quantify biomass production, nutrient concentrations and yield of four different bean genotypes in response to the addition of mulch biomass from
T. diversifolia and
Cajanus cajan (L.) Millsp. in a cut-and-carry system to a P-deficient Andosol.
3. Results
Bean genotypes did not influence soil characteristics significantly. As such, values presented in
Table 1 are mean values of soil chemical characteristics from all bean subplots within each treatment. Only K was significantly different between mulch treatments, showing a greater concentration in the
T. diversifolia mulch biomass treatment.
Table 1.
Soil chemical characteristics (0–12 cm) of an Andosol at the bean flowering phase in treatments with no mulch or mulch biomass from T. diversifolia and C. cajan in a Costa Rican Andosol. Standard errors are given in parentheses.
Table 1.
Soil chemical characteristics (0–12 cm) of an Andosol at the bean flowering phase in treatments with no mulch or mulch biomass from T. diversifolia and C. cajan in a Costa Rican Andosol. Standard errors are given in parentheses.
| | Control | T. diversifolia | C. cajan |
|---|
| pH | 4.9 (0.1)A | 4.7 (0.1)A | 4.9 (0.1)A |
| Exchangeable acidity (cmolc+ L−1) | 1.3 (0.2)A | 1.2 (0.1)A | 1.1 (0.2)A |
| Available P (mg kg−1) | 6.6 (1.0)A | 7.0 (0.6)A | 6.6 (0.5)A |
| Exchangeable K (cmolc+ L−1) | 0.2 (0.1)B | 0.4 (0.1)A | 0.2 (0.1)B |
| Exchangeable Ca (cmolc+ L−1) | 1.9 (0.5)A | 1.7 (0.4)A | 1.8 (0.4)A |
| Exchangeable Mg (cmolc+ L−1) | 0.3 (0.1)A | 0.3 (0.1)A | 0.3 (0.1)A |
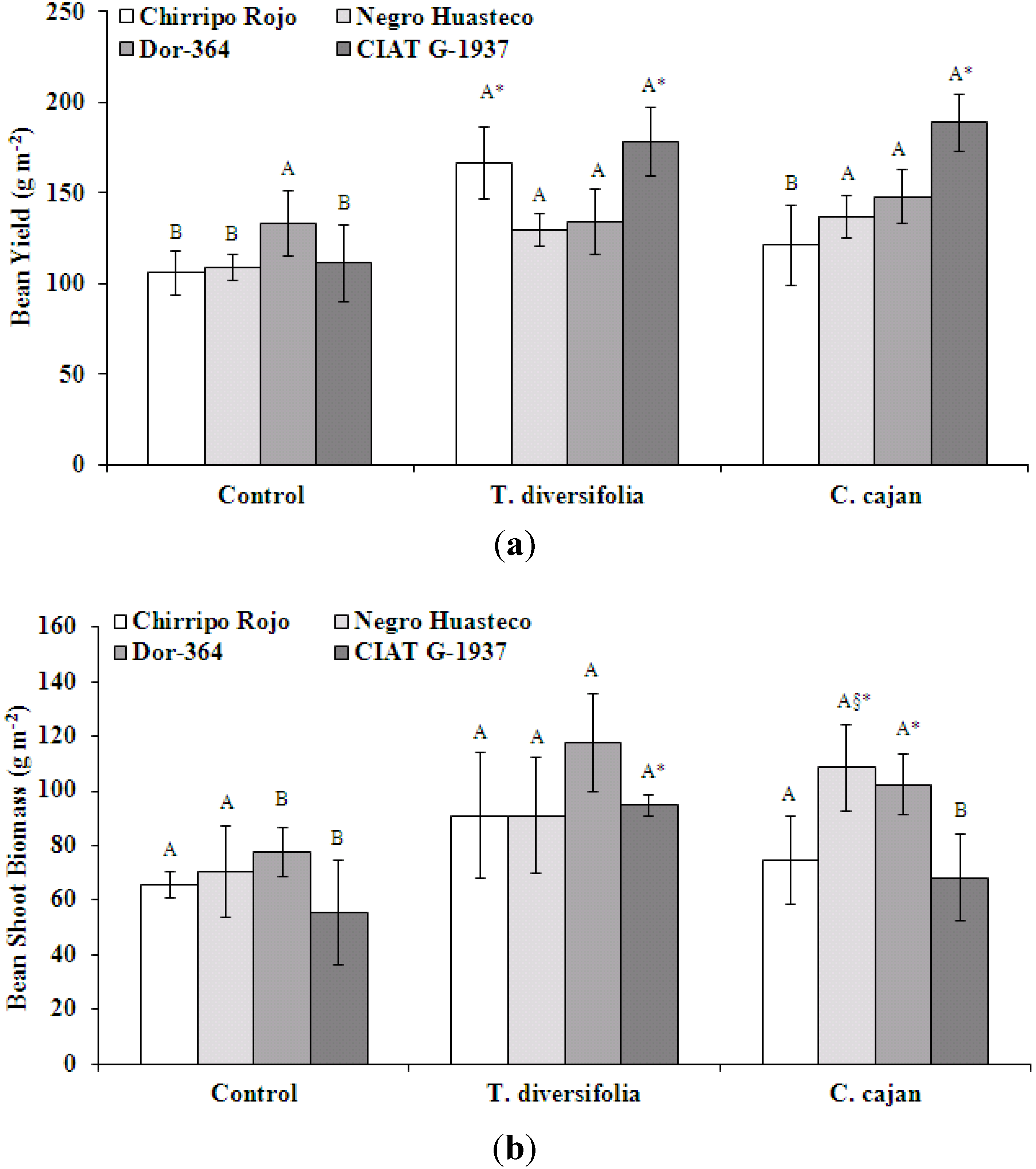
Bean yield (g m
−2) was significantly different between mulch biomass treatments; with a significantly greater yield in both mulch biomass treatments compared to the control (
Figure 1a). When yield is expressed as a mean value of bean genotypes, yield for
T. diversifolia was 152 g m
−2, 149 g m
−2 for
C. cajan and 115 g m
−2 for the control treatment. Chirripo Rojo and CIAT G-1937 had a significantly greater yield in the
T. diversifolia mulch biomass treatment. In the
C. cajan mulch biomass treatment, only CIAT G-1937 had a significantly greater yield. Expressing yield as a mean value of mulch biomass treatments, CIAT G-137 (159 g m
−2) had the greatest yield followed by Dor-364 (138 g m
−2), Chirripo Rojo (131 g m
−2) and Negro Huasteco (125 g m
−2). The interaction effects of mulch treatment-by-bean genotype were not significant for bean yield [
F(6, 25) = 0.619,
p = 0.713].
Bean shoot biomass (g m
−2) was significantly different between mulch biomass treatments (
Figure 1b), with the greatest biomass in the
T. diversifolia mulch treatment for the Dor-364 and CIAT G-1937 genotypes. When shoot biomass is expressed as a mean value of bean genotypes, shoot biomass production for
T. diversifolia was 99 g m
−2, 88 g m
−2 for
C. cajan and 67 g m
−2 for the control treatment. Only Negro Huasteco and Dor-364 were significantly greater compared to the other genotypes in the
C. cajan mulch biomass treatment. Expressing shoot biomass as a mean value of mulch treatments, Dor-364 (99 g m
−2) had the greatest yield followed by Negro Huasteco (90 g m
−2), Chirripo Rojo (77 g m
−2) and CIAT G-1937 (73 g m
−2). The interaction effects of mulch treatment-by-bean genotype were not significant bean shoot biomass [
F(6, 25) = 0.514,
p = 0.792].
Figure 1.
(a) Bean yield (g m−2) at harvest of four different bean genotypes in treatments with no mulch or mulch from T. diversifolia or C. cajan of four different genotypes in treatments with no mulch or mulch from T. diversifolia or C. cajan on a Costa Rican Andosol. Different upper case letters above standard error bars indicate significant differences at p < 0.05 in the comparison of mulch treatments within the same genotype. Values followed by *, comparing differences between bean genotypes and within mulch treatments, are significantly different at p < 0.05; (b) Bean shoot biomass (g m−2) at the flowering phase of four different genotypes in treatments with no mulch or mulch from T. diversifolia or C. cajan on a Costa Rican Andosol. Different upper case letters above standard error bars indicate significant differences at p < 0.05 in the comparison of mulch treatments within the same genotype (§ C. cajan was significantly different from the control only). Values followed by *, comparing differences between bean genotypes and within mulch treatments, are significantly different at p < 0.05.
Figure 1.
(a) Bean yield (g m−2) at harvest of four different bean genotypes in treatments with no mulch or mulch from T. diversifolia or C. cajan of four different genotypes in treatments with no mulch or mulch from T. diversifolia or C. cajan on a Costa Rican Andosol. Different upper case letters above standard error bars indicate significant differences at p < 0.05 in the comparison of mulch treatments within the same genotype. Values followed by *, comparing differences between bean genotypes and within mulch treatments, are significantly different at p < 0.05; (b) Bean shoot biomass (g m−2) at the flowering phase of four different genotypes in treatments with no mulch or mulch from T. diversifolia or C. cajan on a Costa Rican Andosol. Different upper case letters above standard error bars indicate significant differences at p < 0.05 in the comparison of mulch treatments within the same genotype (§ C. cajan was significantly different from the control only). Values followed by *, comparing differences between bean genotypes and within mulch treatments, are significantly different at p < 0.05.
![Agronomy 03 00232 g001]()
Bean shoot nutrient concentrations (mg g
−1) were significantly different between mulch biomass treatments (
Table 2). Bean shoot P concentration was significantly greater in the
T. diversifolia mulch biomass treatment compared to
C. Cajun (CIAT G-1937 genotype only) and the control (Chirripo Rojo and CIAT G-1937 genotypes only). Potassium concentrations followed a similar pattern, showing a significantly greater concentration of this nutrient in the
T. diversifolia mulch biomass treatment compared to that with
C. cajan (Negro Huasteco genotype only) and the control (Negro Huasteco, Dor-364 and CIAT G-1937 bean genotypes only). The control treatment had a significantly greater Ca concentration compared to the treatment with
T. diversifolia (Chirripo Rojo, Negro Huasteco and CIAT G-1937 genotypes only) and
C. Cajun (Negro Huasteco and CIAT G-1937 genotypes only). The control treatment also had a significantly greater Mg concentration compared to the treatment with
T. diversifolia (Chirripo Rojo, Negro Huasteco and CIAT G-1937 genotypes only) and
C. Cajun (Negro Huasteco and CIAT G-1937 genotypes only). When comparing differences between bean genotypes, only Chirripo Rojo had a significantly lower P concentration in the control treatment, whereas K concentration was significantly greater for CIAT G-1937 in the
T. diversifolia mulch biomass treatment. For Mg, Negro Huasteco had a significantly lower concentration of this nutrient in the control treatment. The interaction effects of mulch treatment-by-bean genotype were not significant shoot P [
F(6, 24) = 0.413,
p = 0.863], K [
F(6, 24) = 0.377,
p = 0.886], Ca [
F(6, 24) = 0.477,
p = 0.819] and Mg [
F(6, 24) = 0.675,
p = 0.671] concentrations.
Table 2.
Shoot nutrient concentrations (mg g−1) of four bean genotypes at the flowering phase in treatments with biomass addition from T. diversifolia or C. cajan or without biomass addition (control) on a Costa Rican Andosol. Standard errors are given in parentheses.
Table 2.
Shoot nutrient concentrations (mg g−1) of four bean genotypes at the flowering phase in treatments with biomass addition from T. diversifolia or C. cajan or without biomass addition (control) on a Costa Rican Andosol. Standard errors are given in parentheses.
| | | P (mg g−1) | K (mg g−1) | Ca (mg g−1) | Mg (mg g−1) |
|---|
| Control | Chirripo Rojo | 2.23 (0.18)B * | 28.3 (5.8)A | 19.9 (2.1)A | 3.97 (0.80)A |
| Negro Huasteco | 3.13 (0.33)A | 31.3 (5.5)B | 22.3 (1.2)A | 2.80 (0.21)A * |
| Dor-364 | 3.03 (0.26)A | 28.3 (7.2)B | 21.1 (1.4)A | 3.33 (0.18)A |
| CIAT G-1937 | 2.87 (0.23)B | 29.6 (8.5)B | 20.5 (0.9)A | 4.43 (0.88)A |
| Tithonia diversifolia | Chirripo Rojo | 3.37 (0.07)A | 35.6 (3.4)A | 16.0 (1.1)B | 3.47 (0.48)A |
| Negro Huasteco | 3.20 (0.18)A | 40.5 (3.2)A | 18.3 (2.1)B | 2.28 (0.17)B |
| Dor-364 | 3.30 (0.10)A | 41.5 (1.6)A | 17.8 (2.5)A | 2.63 (0.34)B |
| CIAT G-1937 | 3.35 (0.07)A | 46.9 (2.1)A * | 17.5 (0.4)B | 3.05 (0.61)B |
| Cajanus cajan | Chirripo Rojo | 3.27 (0.33)A | 39.1 (3.8)A | 19.0 (0.8)A | 2.87 (0.26)B |
| Negro Huasteco | 2.93 (0.17)A | 34.6 (1.2)B | 16.7 (2.2)B | 2.47 (0.15)A |
| Dor-364 | 2.97 (0.23)A | 36.9 (5.0)A | 17.9 (1.5)A | 2.77 (0.18)B |
| CIAT G-1937 | 3.03 (0.18)B | 41.8 (3.6)A | 18.7 (2.0)B | 2.63 (0.15)B |
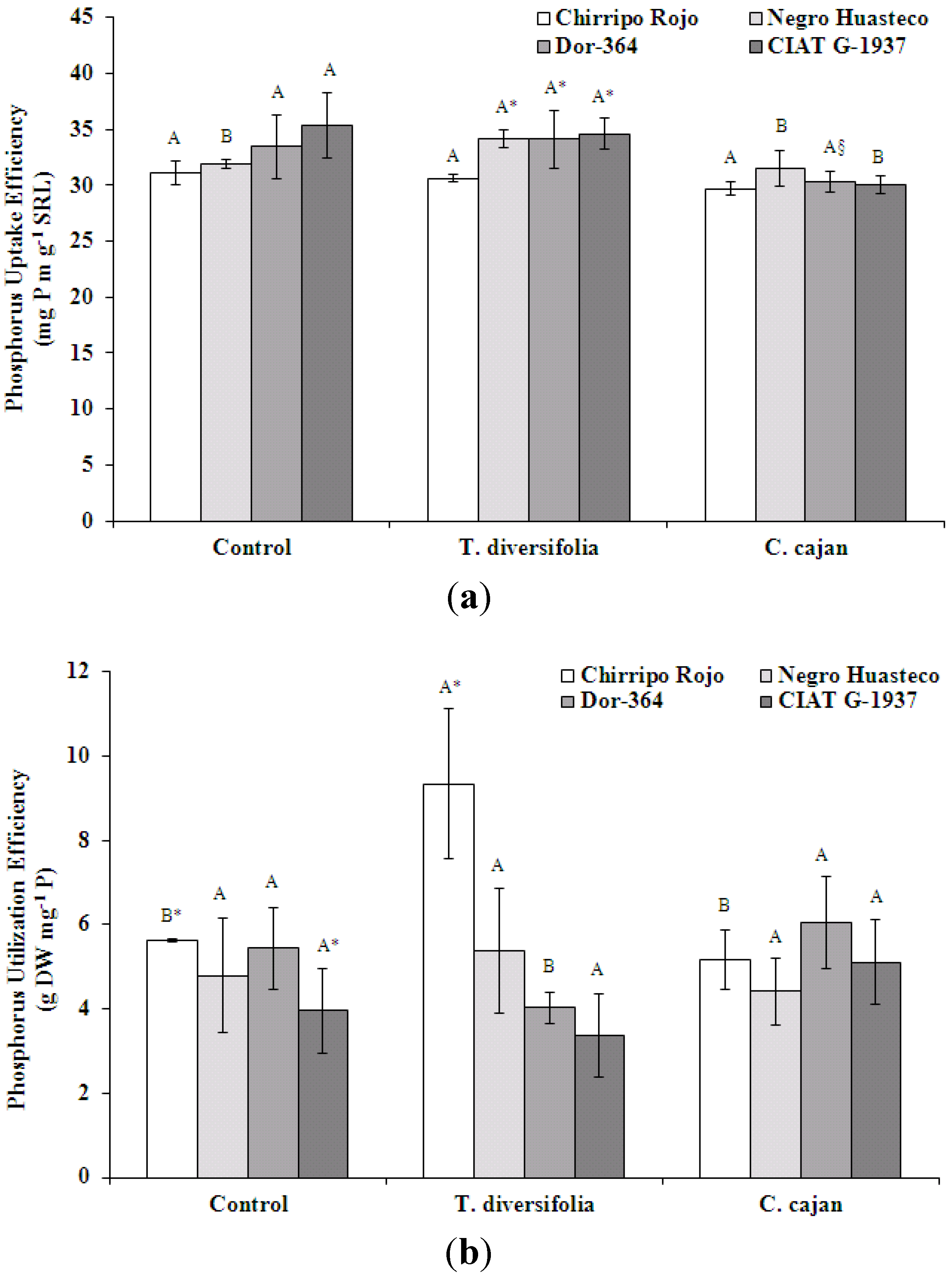
Phosphorus uptake efficiency (mg P m g
−1 SRL) was significantly different between mulch biomass treatments (
Figure 2a). Negro Huasteco had a significantly greater P uptake efficiency in the
T. diversifolia mulch biomass treatment compared to the control. Phosphorus uptake efficiency was also significantly greater for Negro Huasteco, Dor-364 and CIAT G-1937 in the
T. diversifolia mulch biomass treatment compared to
C. cajan. When P uptake efficiency is expressed as a mean value of bean genotypes, P uptake efficiency was 33 mg P m g
−1 SRL for the
T. diversifolia and control treatments and 30 mg P m g
−1 SRL for the
C. cajan mulch biomass treatment. Negro Huasteco, Dor-364 and CIAT G-1937 were significantly greater compared to Chirripo Rojo only in the
T. diversifolia mulch biomass treatment. Expressing P uptake efficiency as a mean value of mulch treatments, P uptake efficiency was 33 mg P m g
−1 SRL (
T. diversifolia) and 30 mg P m g
−1 SRL (
C. cajan) for Chirripo Rojo. The interaction effects of mulch treatment-by-bean genotype were not significant for P uptake efficiency [
F(6, 25) = 0.498,
p = 0.804].
Figure 2.
(a) Bean genotype phosphorus uptake efficiency (mg P m g−1 SRL), where SRL is specific root length at the flowering phase in treatments with no mulch or mulch from T. diversifolia or C. cajan on a Costa Rican Andosol. Different upper case letters above standard error bars indicate significant differences at p < 0.05 in the comparison of mulch treatments within the same genotype (C. cajan were significantly different from T. diversifolia only). Values followed by *, comparing differences between bean genotypes and within mulch treatments, are significantly different at p < 0.05; (b) Bean genotype phosphorus utilization efficiency (g DW mg−1 P) at the flowering phase in treatments with no mulch or mulch from T. diversifolia or C. cajan on a Costa Rican Andosol. Different upper case letters above standard error bars indicate significant differences at p < 0.05 in the comparison of mulch treatments within the same genotype. Values followed by *, comparing differences between bean genotypes and within mulch treatments, are significantly different at p < 0.05.
Figure 2.
(a) Bean genotype phosphorus uptake efficiency (mg P m g−1 SRL), where SRL is specific root length at the flowering phase in treatments with no mulch or mulch from T. diversifolia or C. cajan on a Costa Rican Andosol. Different upper case letters above standard error bars indicate significant differences at p < 0.05 in the comparison of mulch treatments within the same genotype (C. cajan were significantly different from T. diversifolia only). Values followed by *, comparing differences between bean genotypes and within mulch treatments, are significantly different at p < 0.05; (b) Bean genotype phosphorus utilization efficiency (g DW mg−1 P) at the flowering phase in treatments with no mulch or mulch from T. diversifolia or C. cajan on a Costa Rican Andosol. Different upper case letters above standard error bars indicate significant differences at p < 0.05 in the comparison of mulch treatments within the same genotype. Values followed by *, comparing differences between bean genotypes and within mulch treatments, are significantly different at p < 0.05.
![Agronomy 03 00232 g002]()
Phosphorus utilization efficiency (g DW mg
−1 P) was significantly different between mulch biomass treatments, with the greatest utilization efficiency occurring in Chirripo Rojo in the
T. diversifolia treatment (
Figure 2b). When P utilization efficiency is expressed as a mean value of bean genotypes; P utilization efficiency was 5 g DW mg
−1 P for the
C. cajan and control treatments and 6 g DW mg
−1 P for the
T. diversifolia mulch biomass treatment. Chirripo Rojo had a significantly greater P utilization efficiency compared to CIAT G-1937 in the control treatment only. Expressing P uptake efficiency as a mean value of mulch treatments, P uptake efficiency was 7 g DW mg
−1 P for Chirripo Rojo, 5 g DW mg
−1 P for Negro Huasteco and Dor-364 and 4 g DW mg
−1 P for CIAT G-1937. The interaction effects of mulch treatment-by-bean genotype were not a significant P utilization efficiency [
F(6; 25) = 0.888;
p = 0.519].
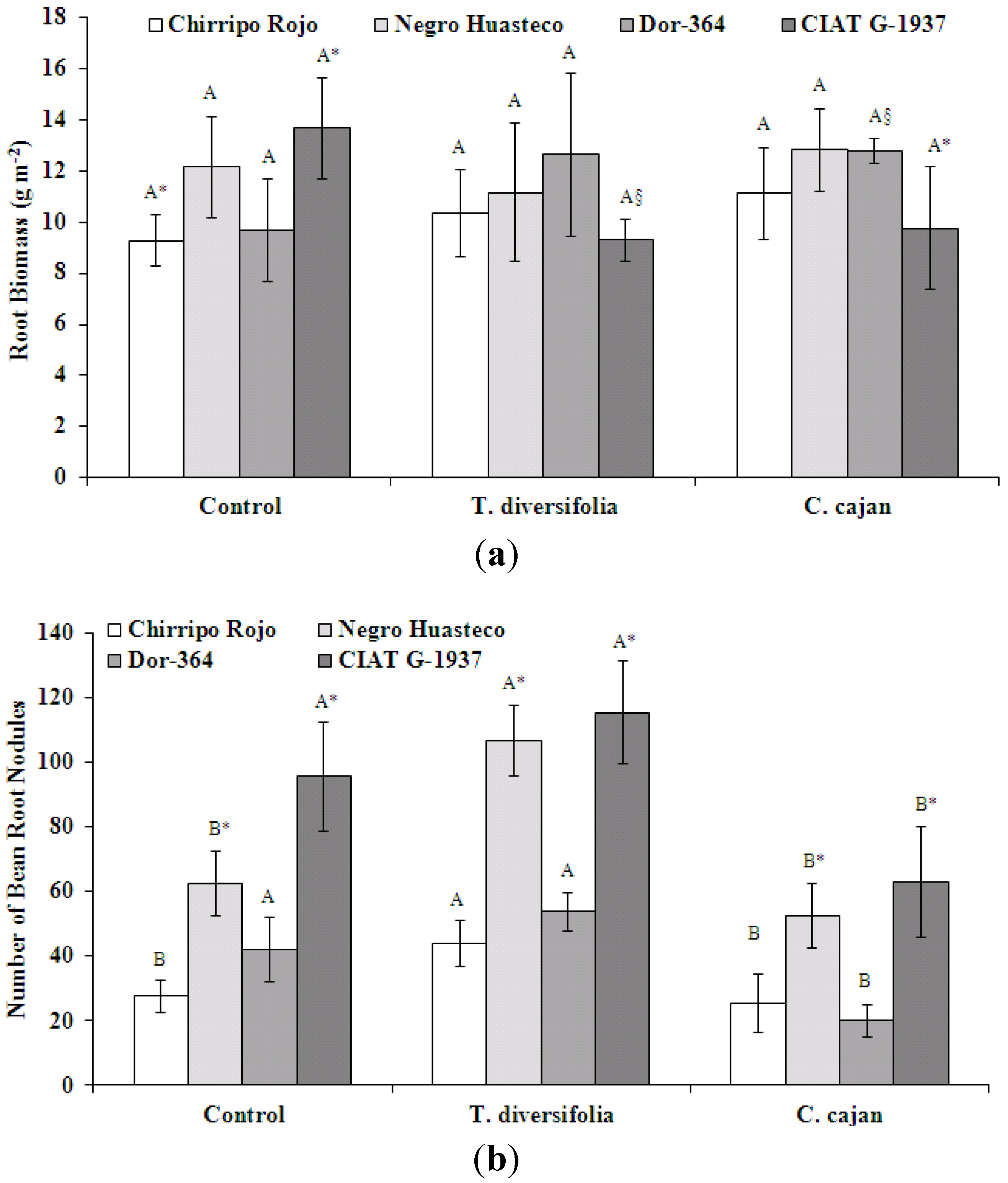
Bean root biomass (g m
−2) was not significantly different between mulch treatments (
Figure 3a). When root biomass is expressed as a mean value of bean genotypes, root biomass was ~11 g m
−2 for all treatments. Differences between bean genotypes showed that Chirripo Rojo had a significantly lower root biomass compared to CIAT G-1937 in the control treatment only. Expressing root biomass as a mean value of mulch treatments, root biomass was 12 g m
−2 for Negro Huasteco and Dor-364, 11 g m
−2 CIAT G-1937 and 10 g m
−2 for Chirripo Rojo. The interaction effects of mulch treatment-by-bean genotype were not significant for root biomass [
F(6, 25) = 0.833,
p = 0.556].
The number of root nodules was significantly different between mulch biomass treatments (
Figure 3b). The number of root nodules was significantly greater for Chirripo Rojo and Negro Huasteco in the
T. diversifolia mulch biomass treatment compared to the control. All genotypes had a significantly greater number of root nodules in the
T. diversifolia mulch biomass treatment compared to the
C. cajan treatment. When the number of root nodules is expressed as a mean value of bean genotypes, the number of nodules was 80 in the
T. diversifolia mulch treatment, 57 in the control and 40 in the
C. cajan mulch biomass treatment. In all three treatments, CIAT G-1937 had a significantly greater number of root nodules. In the
T. diversifolia and
C. cajan treatment, Negro Huasteco also had a significantly greater number of root nodules. Expressing the number of root nodules as a mean value of mulch treatments, the root nodule number was 91 for CIAT G-1937, 71 for Negro Huasteco, 39 for Dor-364 and 32 for Chirripo Rojo. The interaction effects of mulch treatment-by-bean genotype were not significant for the number of root nodules [
F(6, 25) = 0.637,
p = 0.699].
Figure 3.
(a) Root biomass (g m−2) of four different bean genotypes at the flowering phase grown in treatments with no mulch or mulch from T. diversifolia or C. cajan on a Costa Rican Andosol. Different upper case letters above standard error bars indicate significant differences at p < 0.05 in the comparison of mulch treatments within the same genotype [C. cajan (within Dor-364) and T. diversifolia (within CIAT G-1937) were significantly different from the control only]. Values followed by *, comparing differences between bean genotypes and within mulch treatments, are significantly different at p < 0.05; (b) Number of root nodules of four different bean genotypes at the flowering phase grown in treatments with no mulch or mulch from T. diversifolia or C. cajan on a Costa Rican Andosol. Different upper case letters above standard error bars indicate significant differences at p < 0.05 in the comparison of mulch treatments within the same genotype. Values followed by *, comparing differences between bean genotypes and within mulch treatments, are significantly different at p < 0.05.
Figure 3.
(a) Root biomass (g m−2) of four different bean genotypes at the flowering phase grown in treatments with no mulch or mulch from T. diversifolia or C. cajan on a Costa Rican Andosol. Different upper case letters above standard error bars indicate significant differences at p < 0.05 in the comparison of mulch treatments within the same genotype [C. cajan (within Dor-364) and T. diversifolia (within CIAT G-1937) were significantly different from the control only]. Values followed by *, comparing differences between bean genotypes and within mulch treatments, are significantly different at p < 0.05; (b) Number of root nodules of four different bean genotypes at the flowering phase grown in treatments with no mulch or mulch from T. diversifolia or C. cajan on a Costa Rican Andosol. Different upper case letters above standard error bars indicate significant differences at p < 0.05 in the comparison of mulch treatments within the same genotype. Values followed by *, comparing differences between bean genotypes and within mulch treatments, are significantly different at p < 0.05.
![Agronomy 03 00232 g003]()
4. Discussion
Results from this study showed that the addition of mulch from
T. diversifolia or
C. cajan did not substantially affect soil chemistry, including soil pH. Similarly, Amusan
et al. [
26] and Das
et al. [
27] did not observe changes in soil pH when either manure or biomass or a combination of manure and biomass were added to an acidic soil in southwestern Nigeria and India. The lack of a significant increase in soil pH may also be due to the smaller amount of mulch biomass added from
T. diversifolia or
C. cajan compared to other studies [
28]. However, the significantly greater soil K concentration was likely due to a larger quantity of this nutrient in
T. diversifolia biomass compared to
C. cajan. Mustonen
et al. [
17] observed that the concentration of K in aboveground
T. diversifolia biomass was 20.1 mg g
−1 compared to 15.5 mg g
−1 in
C. cajan.
A lack of change in most soil chemical properties evaluated in this study, despite the addition of various amendments, may also be due to the short-term nature of this study. Miller and Miller [
29] suggested that the application of organic materials to crop land could affect soil properties, but this may not be apparent over the short-term (<5 years). For example, The
et al. [
30] found that the long-term application of organic amendments resulted in increased soil P, Ca, Mg and organic matter; and the slow release of these nutrients increased soil fertility and crop yield in subsequent years [
31].
Bean yield was within range of that reported by Mustonen
et al. [
17], Lopez [
32] and Gosh
et al. [
33] in the
T. diversifolia and
C. cajan mulch biomass treatments. Results from our study were similar to that of Mukuralinda
et al. [
34], who found that maize grain yield in Rwanda increased three-fold when biomass from
T. diversifolia was added to the growing maize crop. The increased bean yield observed in our study likely occurred due to the additional nutrients contributed from the input of
T. diversifolia or
C. cajan mulch biomass [
32,
33]. Additionally, previous application of poultry manure on this site in combination with the mulch biomass input from
T. diversifolia or
C. cajan may have resulted in a synergistic effect that positively influenced bean yield, especially in P efficient genotypes, such as CIAT G-1937 [
26]. This is because organic resources from mulch biomass treatments may have the capacity to enhance the availability of P through a variety of mechanisms, including the blocking of P-sorption sites, preventing P-fixation and stimulating microbial P-uptake [
1].
Bean shoot biomass fell within range of that reported from other studies in Central America [
35]. For example, in Costa Rica, Henry
et al. [
36] quantified a shoot biomass of 98 g m
−2 for Chirripo Rojo, which was similar to that in the
T. diversifolia mulch biomass treatment in our study. Aboveground biomass production of all bean genotypes in our study was greater compared to that reported by Roy
et al. [
37] in control and mulch treatments. This may be due to the prior input of poultry manure in our study in addition to a better quality of the mulch biomass from
T. diversifolia or
C. cajan compared to that of rice straw (
Oryza sativa L.) and tropical whiteweed (
Ageratum conyzoides L.) in the study by Roy
et al. [
37].
Miller
et al. [
8] and Fageria
et al. [
38] also observed a greater variability among bean genotypes with respect to bean yield and biomass production. Although shoot biomass production is closely related to yield, this relationship may be difficult to establish in legumes [
38]. This was evident in the P-efficient CIAT G-1937 genotype, which had a greater yield compared to shoot biomass in the mulch treatments. This suggested that the efficient genotypes may allocate a larger amount of resources to the production of pods rather than shoot biomass. In a field experiment in Brazil, evaluating the effect of soil chemistry on bean shoot nutrient concentration, Fageria
et al. [
5] reported results similar to that of our study and found that K had the greatest concentration and P the lowest concentration in bean shoot biomass. This suggested that bean genotypes responded differently to the addition of mulch biomass depending on the nutrient under evaluation [
5]. Ho
et al. [
39] pointed out that such differences may be due to a variation in root architecture among bean genotypes and, therefore, their ability to take-up and accumulate nutrients in aboveground biomass.
Mukuralinda
et al. [
34] and Niang
et al. [
40] reported a 3.5-fold increase in P uptake in treatments with
T. diversifolia mulch biomass compared to other commonly used agroforestry species in West Africa. This may be due to the larger nutrient quality of
T. diversifolia biomass compared to that of
C. cajan [
17] and its greater mineralization potential [
14]. Nielsen
et al. [
22] also found differences among bean genotypes, where the P-efficient CIAT G-1937 had a greater P uptake and utilization efficiency when grown under low, medium and high P availability compared to Dor-364. Although the common bean is a diverse species that includes genotypes with contrasting P uptake and utilization efficiencies, results from our study implied that differences in P utilization efficiency were minimal. Nielsen
et al. [
22] suggested that this is because, at the cellular level, efficient utilization of a commonly limiting nutrient may have been subject to natural selection, and utilization efficiency at the whole plant level may likely be related to fundamental differences in form, size and phenology.
Results from our study showed that the addition of mulch biomass from
T. diversifolia or
C. Cajun did not affect bean root biomass production. Similarly, Roy
et al. [
37] did not observe differences in bean root biomass in treatments with mulch, compost or vermicompost compared to a control. This suggested that the additional nutrients added from
T. diversifolia and
C. cajan mulch biomass were likely allocated to aboveground components, which was reflective of a larger bean yield. Miller
et al. [
8] found that variation among bean genotypes with respect to root biomass production is common even when levels of soil P were low or adequate. Results from our study support that of Nielsen
et al. [
22], who found that P efficient bean genotypes have a larger root biomass under low P availability. Our study showed that in the control treatment, the CIAT G-1937 genotype was more efficient in P uptake (35 mg P m g
−1 SRL) and P utilization (4 g DW mg
−1 P) and had a larger root biomass (14 g m
−2) compared to Chirripo Rojo, which had a P uptake and utilization of 31 mg P m g
−1 SRL and 6 g DW mg
−1 P and a root biomass of 9 g m
−2.
Vargas and Graham [
41] found a large variation in the number of nodules on bean roots and noted that this was mostly dependent on the genotype. They observed that the number of nodules ranged from 0 to 190 per plant [
41]. Increased number of root nodules, especially in the
T. diversifolia mulch treatment, suggested that when soil amendments are used, the establishment of N
2-fixing bacteria is facilitated, resulting in the formation of a larger number of nodules [
42]. Teixeira
et al. [
43] indicated that nodulation depends on the P supply and differs between bean cultivars; some cultivars show an intense decline in the number of nodules after flowering. Time of sampling, therefore, may strongly affect the results in studies evaluating the effect of mulch biomass addition to P-limited soils. Additionally, the input of external amendments, such as manure, inorganic fertilizers or mulch biomass increased the number of nodules [
33]. Gosh
et al. [
33] attributed this to the repeated application of organic matter from manure and/or mulch biomass, which led to an improved soil physical and chemical environment for nitrogenase activity [
33].


{kind=link}
{kind=link}
{kind=link}


