Bacillus Pumilus Strain TUAT-1 and Nitrogen Application in Nursery Phase Promote Growth of Rice Plants under Field Conditions



Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Preparation and Chemical Analysis
2.2. Nursery Preparation
2.3. Transplanting of Rice Plants into the Paddy Field
2.4. Data Analysis
3. Results
3.1. Seedling Growth at Nursery Stage
3.2. Growth at Maximum Tillering Stage
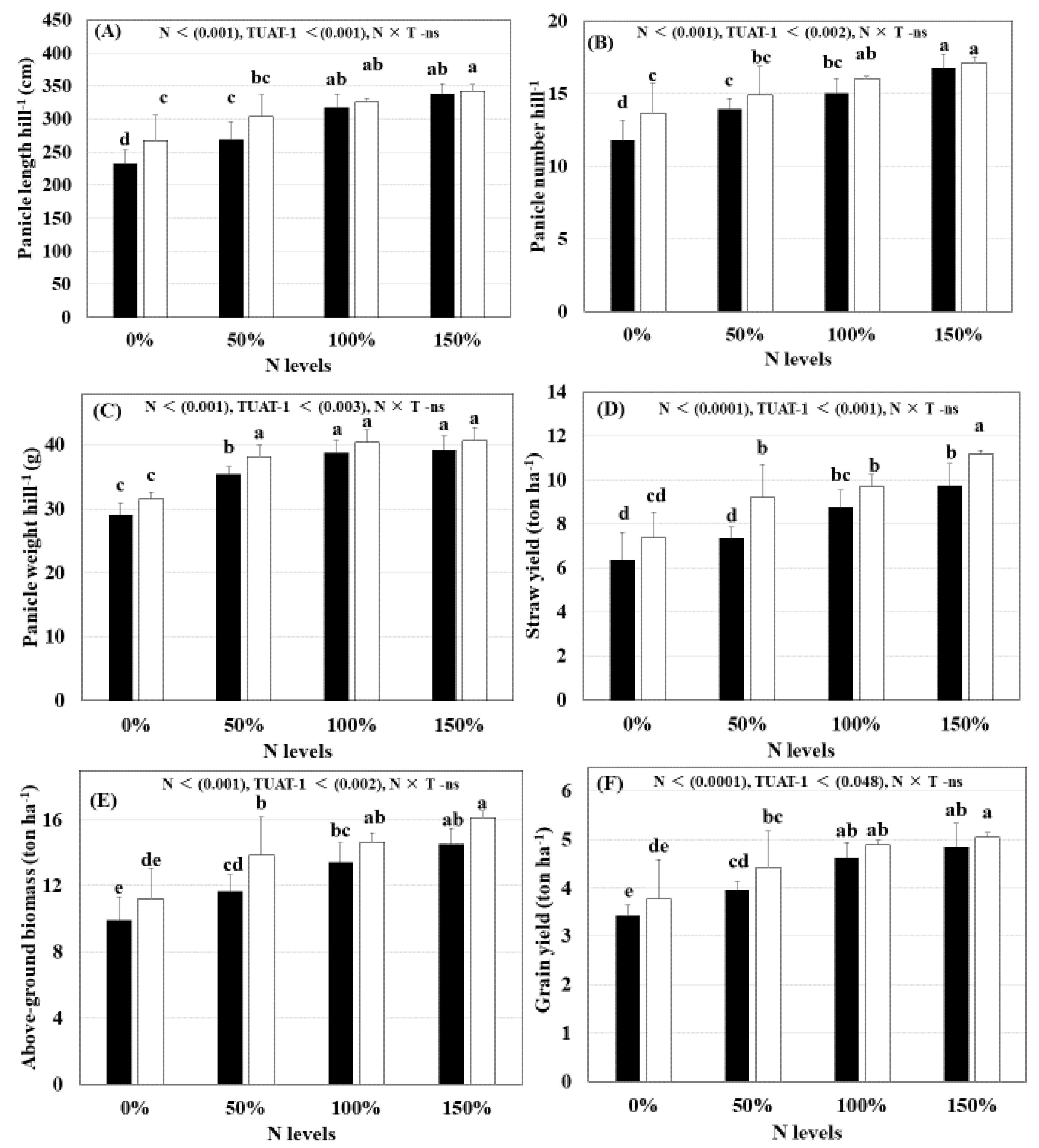
3.3. Yield and Yield Component Parameters
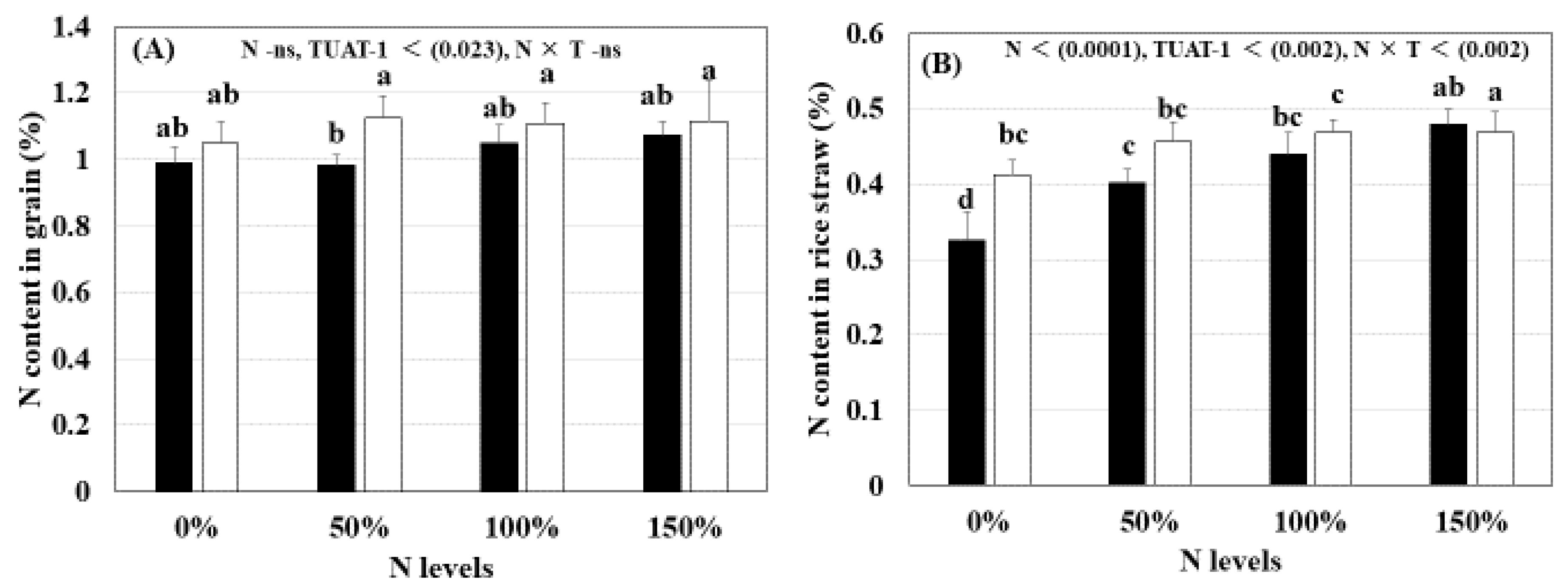
3.4. Nitrogen Content (%) in Grain and Straw
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ali, J.; Aslam, U.M.; Tariq, R.; Murugaiyan, V.; Schnable, P.S.; Li, D.; Marfori-Nazarea, M.C.; Hernandez, J.E.; Arif, M.; Xu, J.; et al. Exploiting the Genomic Diversity of Rice (Oryza sativa L.): SNP-Typing in 11 Early-Backcross Introgression-Breeding Populations. Front. Plant Sci. 2018, 9, 849. [Google Scholar] [CrossRef] [PubMed]
- World Rice Acreage from 2008/2009 to 2015/2016 (In Million Hectares): Statista; 2017. Available online: https://www.statista.com/statistics/271969/world-rice-acreage-since-2008/ (accessed on 21 August 2017).
- Habibah, J.; Le, P.T.; Khairiah, J.; Ahmad, M.R.; Fouzi, B.A.; Ismail, B.S. Speciation of heavy metals in paddy soils from selected areas in Kedah and Penang, Malaysia. Afri. J. Biotechnol. 2011, 4, 13505–13513. [Google Scholar] [CrossRef]
- Souza, R.; Beneduzi, A.; Ambrosini, A.; Costa, P.B.; Meyer, J.; Vargas, L.K.; Schoenfeld, R.; Passaglia, L.M.P. The effect of plant growth-promoting rhizobacteria on the growth of rice (Oryza sativa L.) cropped in southern Brazilian fields. Plant Soil. 2013, 366, 585–603. [Google Scholar] [CrossRef]
- Shariatmadari, Z.; Riahi, H.; Seyed-Hashtroudi, M.; Ghassempour, A.; Aghashariatmadary, Z. Plant growth promoting cyanobacteria and their distribution in terrestrial habitats of Iran. Soil Sci. Plant Nutri. 2013, 59, 535–547. [Google Scholar] [CrossRef] [Green Version]
- Idris, E.E.S.; Iglesias, D.J.; Talon, M.; Borriss, R. Tryptophan-dependent production of Indole 3-Acetic Acid (IAA) affects level of plant growth promotion by Bacillus amyloliquefaciens FZB42. Mol. Plant Microbe Interact. 2007, 20, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Bottini, R.; Cassan, F.; Picolli, P. Gibberellin production by bacteria and its involvement in plant growth promotion and yield increase. Appl. Microbiol. Biotechnol. 2004, 65, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Bloemberg, G.V.; Lugtenberg, B.F.J. Molecular Basis of plant growth promotion and biocontrol by rhizobacteria. Curr. Opin. Plant Biol. 2001, 4, 343–350. [Google Scholar] [CrossRef]
- Aung, H.P.; Salem, D.; Oo, A.Z.; Aye, Y.S.; Yokoyama, T.; Suzuki, S.; Sekimoto, H.; Bellingrath-Kimura, S.D. Growth and 137Cs uptake of four Brassica species influenced by inoculation with a plant growth-promoting rhizobacterium Bacillus pumilus in three contaminated farmlands in Fukushima prefecture, Japan. Sci. Total Environ. 2015, 521, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Torii, A. Analysis of field factors resulting frutuations of yield and nutritional uptakes of forage rice Leaf Star with inoculation of an endophytic nitrogen fixing bacteria TUAT-1. Master Thesis, Tokyo University of Agriculture and Technology, Tokyo, Japan, March 2012. [Google Scholar]
- Rao, V.R.; Jena, P.K.; Adhya, T.K. Inoculation of rice with nitrogen-fixing bacteria problems and perspectives. Biol. Fertil. Soils. 1987, 4, 21–26. [Google Scholar]
- Ikeda, S.; Okubo, T.; Kaneko, T.; Inaba, S.; Maekawa, T.; Eda, S.; Sato, S.; Tabata, S.; Mitsui, H.; Minamisawa, K. Community shifts of soybean stem-associated bacteria responding to different nodulation phenotypes and N levels. ISME J. 2010, 4, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, K.; Ikeda, S.; Eda, S.; Mitsui, H.; Hanzawa, E.; Kisara, C.; Kazama, Y.; Kushida, A.; Shinano, T.; Minamisawa, K.; et al. Impact of plant genotype and nitrogen level on rice growth response to inoculation with Azospirillum sp. Strain B510 under paddy field conditions. Soil Sci. Plant Nutri. 2010, 56, 636–644. [Google Scholar] [CrossRef]
- Blakemore, L.C.; Searle, P.L.; Daly, B.K. Science Report: Methods for Chemical Analysis of Soils; New Zealand Soil Bureau: Australia, 1977.
- Klute, A. Methods of soil analysis, Part 1. In Physical and Mineralogical Methods, 2nd ed.; Soil Science Society of America, Inc.: Madison, WI, USA, 1990. [Google Scholar]
- Yoshinaga, S.; Heinai, H.; Ohsumi, A.; Furuhata, M.; Ishimaru, T. Characteristics of growth and quality, and factors contributing to high yield in newly developed rice variety ‘Akidawara’. Plant Prod. Sci. 2018, 21, 186–192. [Google Scholar] [CrossRef] [Green Version]
- Saito, K.; Fukuta, Y.; Yanagihara, S.; Ahouanton, K.; Sokei, Y. Beyond NERICA: Identifying high-yielding rice varieties adapted to rainfed upland conditions in Benin and their plant Characteristics. Jpn. J. Trop. Agric. 2014, 58, 51–57. [Google Scholar]
- Mizuno, N.; Minami, M. Rapid decomposition method for determination of N, K, Mg, Ca, Fe and Mn in agricultural plants with sulfuric acid and hydrogen peroxide. Jpn. Soil Sci. Plant Nutr. 1980, 51, 418–420. (In Japanese) [Google Scholar]
- Sharma, A.; Shankhdhar, D.; Sharma, A.; Shankhdhar, S.C. Growth promotion of the rice genotypes by PGPRs isolated from rice rhizosphere. J. Soil Sci. Plant Nutri. 2014, 14, 505–517. [Google Scholar] [CrossRef]
- Lynch, J.P.; Lynch, A.F.; Jonathan, P. Roots of the second green revolution. Aust. J. Bot. 2007, 55, 493–512. [Google Scholar] [CrossRef]
- Shaharoona, B.; Naveed, M.; Arshad, M.; Zahir, Z.A. Fertilizer-dependent efficiency of Pseudomonads for improving growth, yield, and nutrient use efficiency of wheat (Triticum aestivum L.). Appl. Microbiol. Biotechnol. 2008, 79, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Spaepen, S.; Vanderleyden, J.; Remans, R. Indole-3-acetic acid in microbial and microorganism-plant signaling. FEMS Microbiol Rev. 2007, 31, 425–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Podile, A.R.; Kishore, G.K. Plant growth promoting rhizobacteria. In Plant Associated Bacteria; Gnanamanickam, S.S., Ed.; Springer: Amsterdam, The Netherlands, 2006; pp. 195–230. [Google Scholar] [CrossRef]
- Badri, D.V.; Vivanco, J.M. Regulation and function of root exudates. Plant Cell Environ. 2009, 32, 666–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Persello-Cartieaux, F.; Naussaume, L.; Robaglia, C. Tales from the underground: Molecular plant-rhizobacteria interactions. Plant Cell Environ. 2003, 26, 189–199. [Google Scholar] [CrossRef]
- Niu, D.D.; Liu, H.X.; Jiang, C.H.; Wang, Y.P.; Wang, Q.Y.; Jin, H.L.; Guo, J.H. The plant growth-promoting rhizobacterium Bacillus cereus AR156 induces systemic resistance in Arabidopsis thaliana by simultaneously activating salicylate-and jasmonate/ethylene-dependent signaling pathways. Mol. Plant Microbe. 2011, 24, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Wahyudi, A.T.; Astuti, R.P.; Widyawati, A.; Meryandini, A.; Nawangsih, A.A. Characterization of Bacillus spp. strains isolated from rhizosphere of soybean plants for their use as potential plant growth for promoting Rhizobacteria. J. Microbiol. Antimicrob. 2011, 2, 406–417. [Google Scholar]
- Ortíz-Castro, R.; Valencia-Cantero, E.; López-Bucio, J. Plant growth promotion by Bacillus megaterium involves cytokinin signaling. Plant Signal Behav. 2008, 3, 263–265. [Google Scholar] [CrossRef] [PubMed]
- Ros, C.; White, P.F.; Bell, R.W. Nursery Fertilizer Application Increases Rice Growth and Yield in Rainfed Lowlands with or without Post-Transplanting Crop Stress. Am. J. Plant Sci. 2015, 6, 2878–2892. [Google Scholar] [CrossRef]
- Panda, M.M.; Reddy, M.D.; Sharma, A.R. Yield performance of rainfed lowland rice as affected by nursery fertilizer under conditions of intermediate deepwater (15–50 cm) and flash floods. Plant Soil. 1991, 132, 65–71. [Google Scholar] [CrossRef]
- Kumar, K.V.K.; Yellareddygari, S.K.; Reddy, M.S.; Kloepper, J.W.; Lawrence, K.S.; Zhou, X.G.; Groth, D.E.; Krishnam Raju, S.K.; Miller, M.E. Efficacy of Bacillus subtilis MBI 600 against sheath blight caused by Rhizoctonia solani and on growth and yield of rice. Rice Sci. 2012, 9, 55–63. [Google Scholar] [CrossRef]
- Wang, Y.; Thorup-Kristensen, K.; Jensen, L.S.; Magid, J. Vigorous Root Growth Is a Better Indicator of Early Nutrient Uptake than Root Hair Traits in Spring Wheat Grown under Low Fertility. Front. Plant Sci. 2016, 7, 865. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, S.; Ban, T.; Kanda, S.; Win, A.T.; Toyota, K. Biomass production and nutrient cycling. In Research Approaches to Sustainable Biomass Systems; Tojo, S., Hirasawa, T., Eds.; Elsevier: Oxford, UK, 2013; pp. 290–294. [Google Scholar]
- Khan, A.; Sirajuddin, M.; Zhao, X.Q.; Javed, T.; Khan, K.S.; Bano, A.; Shen, R.F.; Masood, S. Bacillus pumilus enhances tolerance in rice (Oryza sativa L.) to combined stresses of NaCl and high boron due to limited uptake of Na+. Environ. Experi. Bot. 2016, 124, 120–129. [Google Scholar] [CrossRef]
- Armada, E.; Azcón, R.; López-Castillo, O.M.; Calvo-Polanco, M.; Ruiz-Lozano, J.M. Autochthonous arbuscular mycorrhizal fungi and Bacillus thuringiensis from a degraded Mediterranean area can be used to improve physiological traits and performance of a plant of agronomic interest under drought conditions. Plant Physiol. Biochem. 2015, 90, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Yadav, J.; Verma, J.P.; Jaiswal, D.K.; Kumar, A. Evaluation of PGPR and different concentration of phosphorus level on plant growth, yield and nutrient content of rice (Oryza sativa). Ecol. Eng. 2014, 62, 123–128. [Google Scholar]
- Mishra, R.P.N.; Singh, R.K.; Jaiswal, H.K.; Kumar, V.; Maurya, S. Rhizobium-mediated induction of phenolics and plant growth promotion in rice (Oryza sativa L.). Curr. Microbiol. 2006, 52, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Okon, Y.; Labandera-Gonzalez, C.A. Agronomic applications of Azospirillum—An evaluation of 20 years worldwide field inoculation. Soil Biol. Biochem. 1994, 26, 1591–1601. [Google Scholar] [CrossRef]
- Dobbelaere, S.; Croonenborghs, A.; Thys, A.; Ptacek, D.; Vanderleyden, J.; Dutto, P.; Labandera-Gonzalez, C.; Caballero-Mellado, J.; Aguirre, J.F.; Kapulnik, Y.; et al. Responses of agronomically important crops to inoculation with Azospirillum. Aust. J. Plant Physiol. 2001, 28, 871–879. [Google Scholar] [CrossRef]
- Adesemoye, A.O.; Torbert, H.A.; Kloepper, J.W. Increased plant uptake of nitrogen from 15N-depleted fertilizer using plant growth promoting rhizobacteria. Appl. Soil Ecol. 2010, 46, 54–58. [Google Scholar] [CrossRef]
- Duarah, I.; Deka, M.; Saikia, N.; Deka, B.H.P. Phosphate solubilizers enhance NPK fertilizer use efficiency in rice and legume cultivation. 3 Biotech 2011, 1, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Adesemoye, A.O.; Egamberdieva, D. Beneficial effects of plant growth-promoting rhizobacteria on improved crop production: Prospects for developing economies. In Bacteria in Agrobiology: Crop Productivity; Maheshwari, D.K., Saraf, M., Aeron, A., Eds.; Springer: Berlin, Germany, 2013; pp. 45–63. [Google Scholar] [CrossRef]
- Liu, X.; Wang, H.; Zhou, J.; Hu, F.; Zhu, D.; Chen, Z.; Liu, Y. Effect of N fertilization pattern on rice yield, N use efficiency and fertilizer-N fate in the Yangtze river Basin, China. PLoS ONE 2016, 11, e0166002. [Google Scholar] [CrossRef] [PubMed]
- Çakmakçi, R.; Dönmez, F.; Aydın, A.; Şahin, F. Growth promotion of plants by plant growth-promoting rhizobacteria under greenhouse and two different field soil conditions. Soil Biol. Biochem. 2006, 38, 1482–1487. [Google Scholar] [CrossRef]
- Biswas, J.C.; Ladha, J.K.; Dazzo, F.B.; Yanni, Y.G.; Rolfe, B.G. Rhizobial inoculation influences seedling vigor and yield of rice. Agron. J. 2000, 92, 880–886. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Soil Physiochemical Property | Value |
|---|---|
| pH (H2O) | 6.25 |
| Total carbon content (%) | 5.25 |
| Total nitrogen content (%) | 0.33 |
| Cation-exchange capacity (cmolc/kg) | 20.55 |
| Sand (%) | 40.12 |
| Silt (%) | 32.27 |
| Clay (%) | 27.61 |
| Treatments | SPAD | SL (cm) | SB (g) | RB (g) | TRL (cm) | RSA (cm2) |
|---|---|---|---|---|---|---|
| 0% N | 18.6 c | 12.1 cd | 16.0 b | 15.3 d | 12.7 f | 240.3 d |
| 0% N + TUAT-1 | 19.6 c | 13.6 c | 21.6 b | 20.6 c | 19.7 de | 335.7 c |
| 50% N | 18.1 bc | 15.2 ab | 27.0 a | 18.3 c | 17.1 ef | 268.7 cd |
| 50% N + TUAT-1 | 24.8 cd | 15.0 bc | 29.1 a | 25.0 b | 29.2 bc | 466.9 b |
| 100% N | 21.4 bc | 15.0 ab | 25.3 a | 19.6 c | 18.7 de | 315.7 c |
| 100% N + TUAT-1 | 26.8 bc | 15.2 ab | 25.9 a | 25.9 b | 31.9 b | 520.9 ab |
| 150% N | 25.4 ab | 16.3 a | 26.9 a | 23.8 b | 26.6 cd | 405.3 b |
| 150% N + TUAT-1 | 32.1 a | 16.2 a | 28.1 a | 29.2 a | 39.2 a | 682.0 a |
| Analysis of variance | p value | |||||
| Nitrogen (N) | 0.001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 |
| TUAT-1 (T) | 0.006 | 0.957 | 0.127 | 0.0001 | 0.0001 | 0.0001 |
| N × T | 0.439 | 0.512 | 0.387 | 0.76 | 0.074 | 0.127 |
| Treatments | SPAD | Plant Height (cm) | TN | TB (g) |
|---|---|---|---|---|
| 0% N | 39.6 d | 82.2 c | 8.7 c | 9.5 d |
| 0% N + TUAT-1 | 40.2 d | 86.7 ab | 13.6 ab | 17.0 c |
| 50% N | 40.0 d | 82.8 c | 11.2 bc | 10.7 d |
| 50% N + TUAT-1 | 41.4 c | 86.8 ab | 13.4 ab | 18.4 bc |
| 100% N | 40.5 c | 84.4 bc | 12.5 ab | 16.2 bc |
| 100% N + TUAT-1 | 41.7 b | 88.4 a | 15.7 a | 20.9 a |
| 150% N | 42.1 a | 87.6 a | 13.7 ab | 18.6 ab |
| 150% N + TUAT-1 | 42.3 ab | 89.1 a | 16.4 a | 21.8 a |
| Analysis of variance | p value | |||
| Nitrogen (N) | 0.002 | 0.0001 | 0.004 | 0.0001 |
| TUAT-1 (T) | 0.001 | 0.001 | 0.002 | 0.0001 |
| N × T | 0.232 | 0.023 | 0.536 | 0.029 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Win, K.T.; Oo, A.Z.; Ohkama-Ohtsu, N.; Yokoyama, T. Bacillus Pumilus Strain TUAT-1 and Nitrogen Application in Nursery Phase Promote Growth of Rice Plants under Field Conditions. Agronomy 2018, 8, 216. https://doi.org/10.3390/agronomy8100216
Win KT, Oo AZ, Ohkama-Ohtsu N, Yokoyama T. Bacillus Pumilus Strain TUAT-1 and Nitrogen Application in Nursery Phase Promote Growth of Rice Plants under Field Conditions. Agronomy. 2018; 8(10):216. https://doi.org/10.3390/agronomy8100216
Chicago/Turabian StyleWin, Khin Thuzar, Aung Zaw Oo, Naoko Ohkama-Ohtsu, and Tadashi Yokoyama. 2018. "Bacillus Pumilus Strain TUAT-1 and Nitrogen Application in Nursery Phase Promote Growth of Rice Plants under Field Conditions" Agronomy 8, no. 10: 216. https://doi.org/10.3390/agronomy8100216
APA StyleWin, K. T., Oo, A. Z., Ohkama-Ohtsu, N., & Yokoyama, T. (2018). Bacillus Pumilus Strain TUAT-1 and Nitrogen Application in Nursery Phase Promote Growth of Rice Plants under Field Conditions. Agronomy, 8(10), 216. https://doi.org/10.3390/agronomy8100216