Growing Degree Days during the Late Reproductive Phase Determine Spike Density and Cognate Yield Traits

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Conditions
2.2. Agronomic Practices and Data Collection
2.3. Statistical Analysis
3. Results and Discussion
3.1. Estimation of Mean Square Values
3.2. Study of Mean Values among Parents
3.3. Estimation of GCA and SCA Effects
3.4. Proportional Contribution of Line and Testers
3.5. Genetic Components and Degree of Dominance
3.6. Heritability, Genetic Gain and Correlation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Slafer, G.A.; Abeledo, L.G.; Miralles, D.J.; Gonzalez, F.G.; Whitechurch, E.M. Photoperiod sensitivity during stem elongation as an avenue to raise potential yield in wheat. Euphytica 2001, 119, 191–197. [Google Scholar] [CrossRef]
- Slafer, G.A. Differences in phasic development rate amongst wheat cultivars independent of responses to photoperiod and vernalization. A viewpoint of the intrinsic earliness hypothesis. J. Agric. Sci. 1996, 126, 403–419. [Google Scholar] [CrossRef]
- Liang, Y.; Farooq, M.U.; Zeng, R.; Tang, Z.; Zhang, Y.; Zheng, T.; Ei, H.H.; Ye, X.; Jia, X.; Zhu, J. Breeding of selenium rich red glutinous rice, protein extraction and analysis of the distribution of selenium in grain. Int. J. Agric. Biol. 2018, 20, 1005–1011. [Google Scholar]
- Liang, Y.; Farooq, M.U.; Hu, Y.; Tang, Z.; Zhang, Y.; Zeng, R.; Zheng, T.; Ei, H.H.; Ye, X.; Jia, X.; et al. Study on stability and antioxidant activity of red anthocyanidin glucoside rich hybrid rice, its nutritional and physicochemical characteristics. Food Sci. Technol. Res. 2018, 24, 687–696. [Google Scholar]
- Farooq, M.U.; Tang, Z.; Zeng, R.; Liang, Y.; Zhang, Y.; Zheng, T.; Ei, H.H.; Ye, X.; Jia, X.; Zhu, J. Accumulation, mobilization and transformation of selenium in rice grain provided with foliar sodium selenite. J. Sci. Food Agric. 2018. In press. [Google Scholar]
- Reynolds, M.P.; Rajaram, S.; Mcnab, A. Increasing Yield Potential in Wheat: Breaking the Barriers; CIMMYT: Ciudad Obregon, Sonora, Mexico, 1996. [Google Scholar]
- Ali, M.A.; Nawab, N.N.; Rasool, G.; Saleem, M. Estimates of variability and correlations for quantitative traits in Cicer arietinum. J. Agric. Soc. Sci. 2008, 4, 177–179. [Google Scholar]
- Khaliq, I.; Parveen, N.; Chowdhry, M.A. Correlation and path coefficient analyses in bread wheat. Int. J. Agric. Biol. 2004, 6, 633–635. [Google Scholar]
- Jain, S.; Sastry, E. Heterosis and combining ability for grain yield and its contributing traits in bread wheat (Triticum aestivum L.). J. Agric. Allied Sci. 2012, 1, 17–22. [Google Scholar]
- Iqbal, M.; Navabi, A.; Salmon, D.F.; Yang, R.-C.; Murdoch, B.M.; Moore, S.S.; Spane, D. Genetic analysis of flowering and maturity time in high latitude spring wheat. Euphytica 2007, 154, 207–218. [Google Scholar] [CrossRef]
- Chowdhary, M.; Sajad, M.; Ashraf, M. Analysis on combining ability of metric traits in bread wheat, Triticum aestivum. J. Agric. Res. 2007, 45, 11–17. [Google Scholar]
- Mangi, S.A.; Sial, M.A.; Ansari, B.A.; Arain, M.A. Study of genetic parameters in segregating populations of spring wheat. Pak. J. Bot. 2008, 39, 2407–2413. [Google Scholar]
- Widyaratne, G.; Zijlstra, R. Nutritional value of wheat and corn distiller’s dried grain with solubles: Digestibility and digestible contents of energy, amino acids and phosphorus, nutrient excretion and growth performance of grower-finisher pigs. Can. J. Anim. Sci. 2007, 87, 103–114. [Google Scholar] [CrossRef] [Green Version]
- Farooq, M.U.; Cheema, A.A.; Ishaaq, I.; Zhu, J. Correlation and genetic component studies for peduncle length affecting grain yield in wheat. Int. J. Adv. Appl. Sci. 2018, 5, 67–75. [Google Scholar] [CrossRef]
- Sayre, K.D.; Rajaram, S.; Fischer, R. Yield potential progress in short bread wheats in northwest Mexico. Crop Sci. 1997, 37, 36–42. [Google Scholar] [CrossRef]
- Nielsen, R. What Exactly do You Mean by ‘Growing Degree Day’. Available online: https://www.agry.purdue.edu/ext/corn/news/articles.01/Corn_GDD_Calc-0423.pdf (accessed on 1 May 2015).
- Fellahi, Z.E.A.; Hannachi, A.; Bouzerzour, H.; Boutekrabt, A. Line× tester mating design analysis for grain yield and yield related traits in bread wheat (Triticum aestivum L.). Int. J. Agron. 2013. [Google Scholar] [CrossRef]
- Panse, V.G. Statistical Methods for Agricultural Workers; Indian Council of Agricultural Research: New Delhi, Indian, 1954; pp. 154–196. [Google Scholar]
- Steel, R.G.D.; Torrie, J.C. Principles and Procedures of Statistics: A Biometrical Approach; McGraw-Hill: New York, NY, USA, 1980. [Google Scholar]
- Kempthorne, O. An Introduction to Genetic Statistics; John Wiley and Sons, Inc.: New York, NY, USA, 1957. [Google Scholar]
- Team, R.C. R: A Language and Environment for Statistical Computing; The R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Clewer, A.G.; Scarisbrick, D.H. Practical Statistics and Experimental Design for Plant and Crop Science; John Wiley and Sons, Inc.: Chichester, UK; New York, NY, USA, 2001. [Google Scholar]
- Singh, N.; Singh, Y.; Singh, V. Variation in physiological traits in promising wheat varieties under late sown condition. Indian J. Plant Physiol. 2005, 10, 171. [Google Scholar]
- Verma, O.; Srivastava, H. Genetic component and combining ability analyses in relation to heterosis for yield and associated traits using three diverse rice-growing ecosystems. Field Crop. Res. 2004, 88, 91–102. [Google Scholar] [CrossRef]
- Tiwari, D.K.; Pandey, P.; Giri, S.P.; Dwivedi, J.L. Prediction of gene action, heterosis and combining ability to identify superior rice hybrids. Int. J. Bot. 2011, 7, 126–144. [Google Scholar] [CrossRef]
- Sarker, U.; Biswas, P.S.; Prasad, B.; Mian, M.K. Heterosis and genetic analysis in rice hybrids. Pak. J. Biol. Sci. 2002, 5, 1–5. [Google Scholar]
- Rashid, M.; Cheema, A.A.; Ashraf, M. Line × tester analysis in basmati rice. Pak. J. Bot. 2007, 39, 2035–2042. [Google Scholar]
- Borghi, B.; Perenzin, M.; Nash, R. Combining ability estimates in bread wheat and performance of 100 F1 hybrids produced using a chemical hybridising agent. J. Genet. Breed. 1989, 43, 11–16. [Google Scholar]
- Javaid, A.; Masood, S.; Minhas, N. Analysis of combining ability in wheat (Triticum aestivum L.) using F2 generation. Pak. J. Biol. Sci. 2001, 4, 1303–1305. [Google Scholar]
- Nazeer, W.; Hussain, T.; Khan, M.A.; Naeem, M.; Amjid, M.W.; Hussain, K. Mechanism of inheritance for quantitative traits in interaspecific crosses of Triticum aestivum L. World Appl. Sci. J. 2013, 22, 1440–1448. [Google Scholar]
- Hussain, Q.; Aziz, T.; Khalil, I.H.; Ahmad, N.; Asim, M.; Adnan, M.; Farooq, M.U. Estimation of heritability and selection response for some yield traits in F3 populations of wheat. Int. J. Agric. Appl. Sci. 2017, 9, 6–13. [Google Scholar]
- Yadav, A.K.; Maan, R.K.; Kumar, S.; Kumar, P. Research note variability, heritability and genetic advance for quantitative characters in hexaploid wheat (Triticum aestivum L.). Electron. J. Plant Breed. 2011, 2, 405–408. [Google Scholar]
- Miralles, D.J.; Katz, S.D.; Colloca, A.; Slafer, G.A. Floret development in near isogenic wheat lines differing in plant height. Field Crop. Res. 1998, 59, 21–30. [Google Scholar] [CrossRef]
- Rajper, A.A.; Baloch, S.K.; Baloch, K.; Ahmed, S.; Kaleri, A.A.; Leghari, A.L.; Kaleri, S.H.; Soothar, J.K.; Soomro, S.R.; Kaleri, R.R. 14. Analysis path coefficient of yield earliness traits in wheat (Triticum aestivum L.). Pure Appl. Biol. 2018, 7, 112–120. [Google Scholar]
- Slafer, G.A.; Rawson, H. Sensitivity of wheat phasic development to major environmental factors: A re-examination of some assumptions made by physiologists and modellers. Funct. Plant Biol. 1994, 21, 393–426. [Google Scholar] [CrossRef]




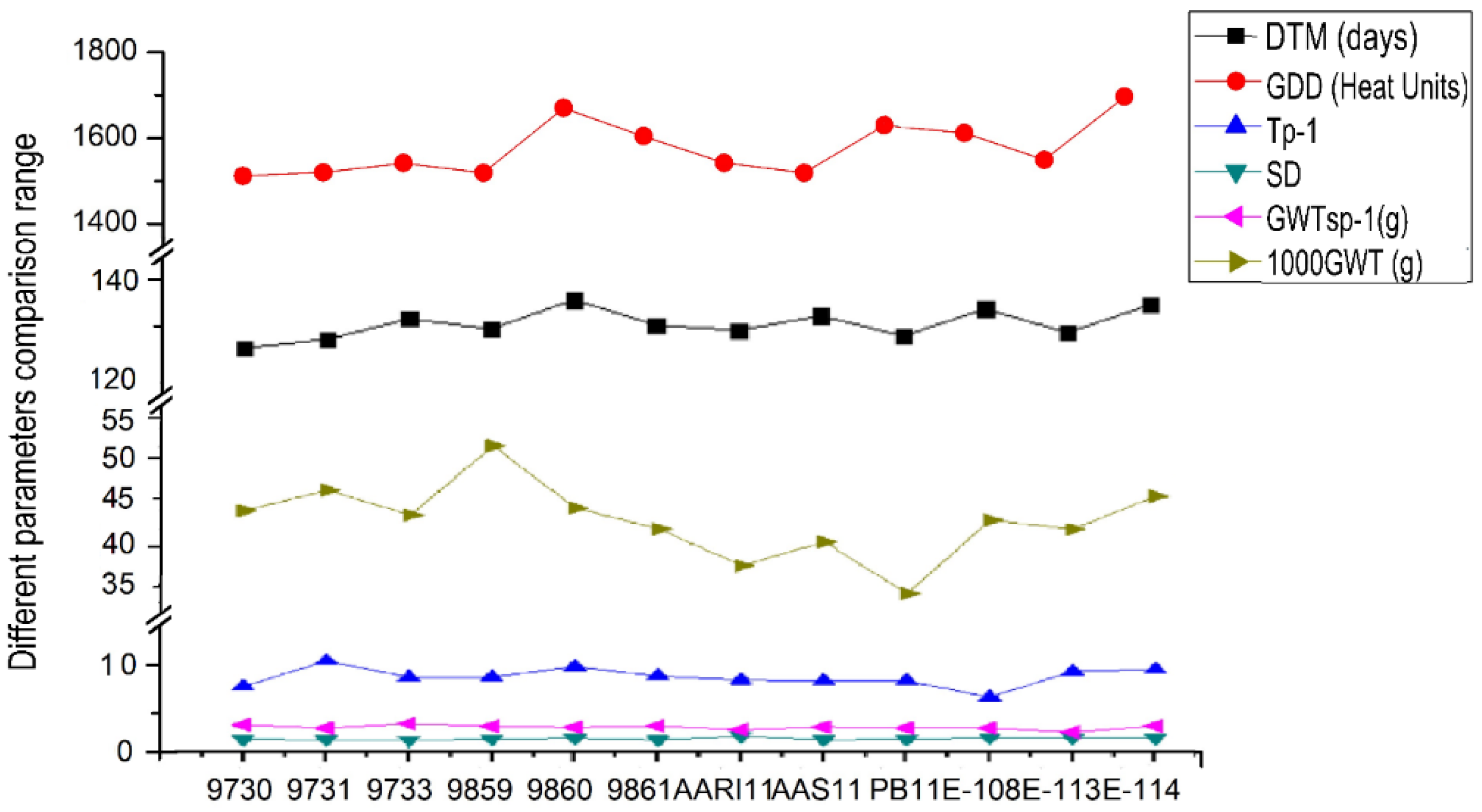{kind=link}
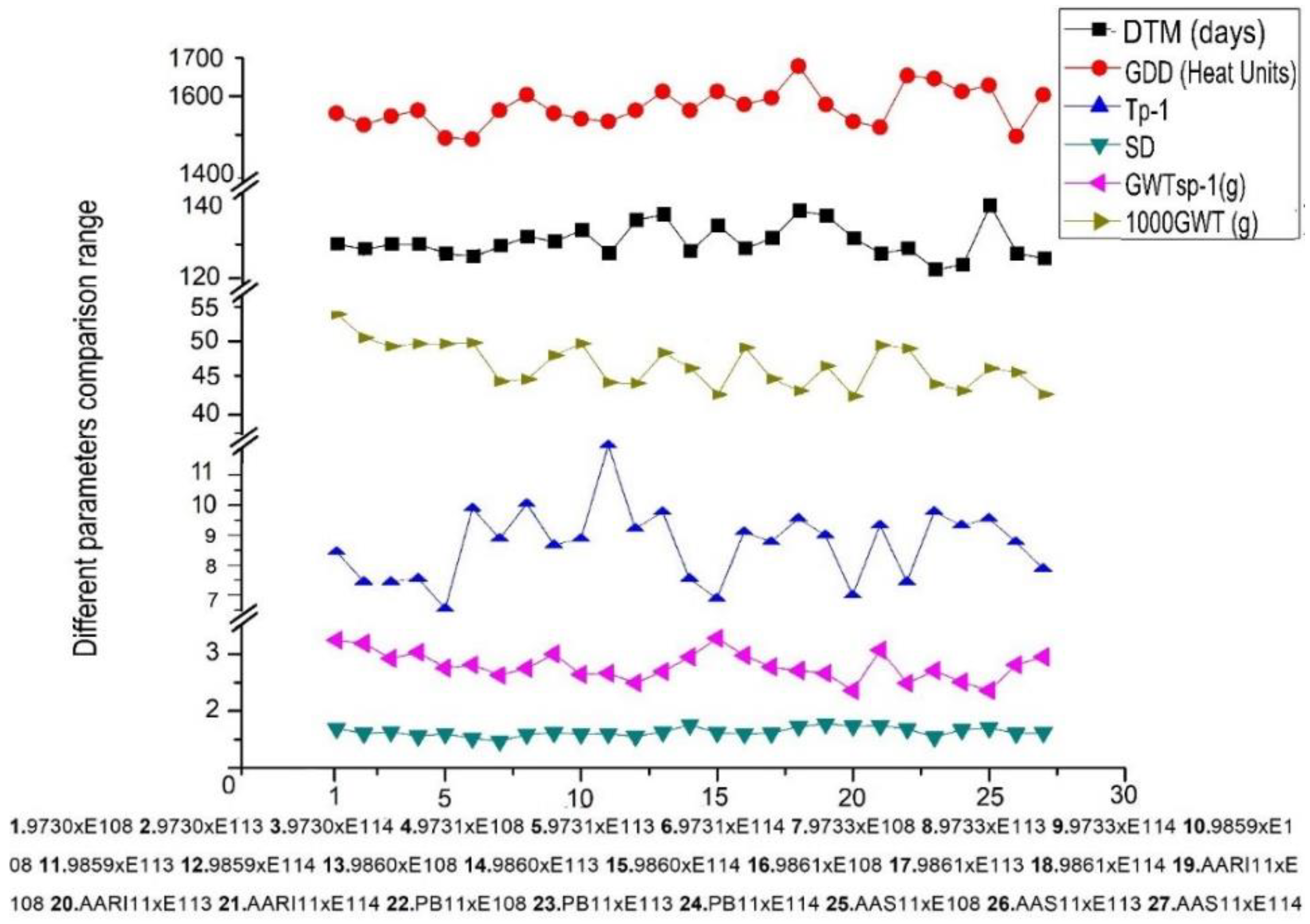{kind=link}
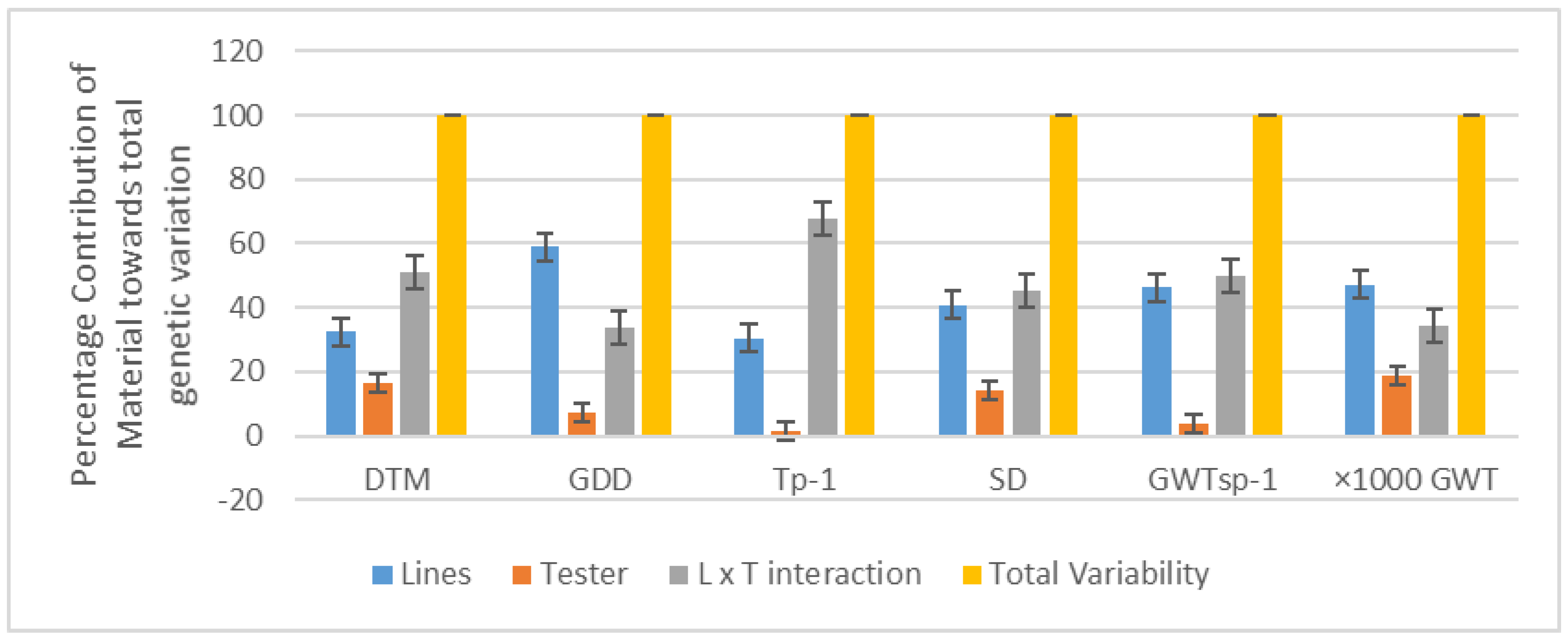{kind=link}
| Source of Variation | d.f | DTM | GDD | Tp−1 | SD | GWTsp−1 | ×1000 GWT |
|---|---|---|---|---|---|---|---|
| Replications | 2 | 0.577 NS | 1810.200 * | 0.855 NS | 0.0097776 NS | 0.106 NS | 2.943 NS |
| Treatments | 38 | 32.710 ** | 8531.742 ** | 4.086 ** | 0.027416 ** | 0.196 ** | 46.924 ** |
| Parents | 11 | 23.174 ** | 12281.764 ** | 3.684 ** | 0.00302 ** | 0.202 ** | 58.984 ** |
| Parents vs. crosses | 1 | 0.145 NS | 372.657 NS | 0.201 NS | 0.313389 * | 0.398 * | 337.790 ** |
| Crosses | 26 | 37.997 ** | 7259.005 ** | 4.405 ** | 0.017475 ** | 0.185 ** | 30.634 ** |
| Lines | 8 | 40.255 ** | 13944.306 ** | 4.566 ** | 0.007112 ** | 0.278 ** | 46.898 ** |
| Testers | 2 | 80.658 ** | 6959.387 ** | 0.044 NS | 0.02436 ** | 0.093 NS | 73.961 ** |
| Lines × Testers | 16 | 31.535 ** | 3953.807 ** | 4.870 ** | 0.036279 * | 0.150 * | 17.086 ** |
| Error | 76 | 0.349 | 406.072 | 0.712 | 0.006678 | 0.080 | 1.477 |
| Lines | DTM | GDD | Tp−1 | SD | GWTsp−1 | ×1000 GWT |
|---|---|---|---|---|---|---|
| 9730 | −0.977 NS | −28.812 NS | −0.918 NS | 1.245 * | 0.324 NS | 4.685 NS |
| 9731 | −2.177 NS | −57.585 NS | −0.696 NS | −0.283 NS | 0.071 * | 3.130 NS |
| 9733 | 0.004 * | 2.132 * | 0.504 * | −0.070 NS | 0.000 NS | −1.037 NS |
| 9859 | 1.463 NS | −25.590 NS | 1.340 NS | 0.306 * | −0.194 NS | −0.704 NS |
| 9860 | 2.352 NS | 23.504 NS | −0.624 NS | 0.447 * | 0.180 * | −1.004 NS |
| 9861 | 1.887 NS | 45.438 NS | 0.449 * | 2.191 ** | 0.026 * | −1.093 NS |
| AARI-11 | 1.142 NS | −27.835 NS | −0.252 NS | 0.508 * | −0.096 NS | −0.626 NS |
| AAS-11 | −4.163 NS | 64.660 NS | 0.155 * | −1.640 * | −0.225 NS | −1.393 NS |
| PB-11 | 0.468 ** | 4.088 * | 0.042 * | 2.063 ** | −0.087 NS | −1.959 NS |
| Testers | ||||||
| E-108 | 1.767 NS | 13.956 ** | 0.043 * | 1.597 * | −0.046 NS | −0.963 NS |
| E-113 | −1.687 NS | −17.546 NS | −0.037 NS | 1.448 * | −0.020 NS | −0.948 NS |
| E-114 | −0.079 NS | 3.590 * | −0.006 NS | 2.222 * | 0.066 * | 1.911 NS |
| S.E for lines | 0.197 | 6.717 | 0.281 | 0.794 | 0.094 | 0.405 |
| S.E for testers | 0.114 | 3.878 | 0.162 | 0.797 | 0.054 | 0.234 |
| Hybrids | DTM | GDD | Tp−1 | SD | GWTsp−1 | ×1000 GWT |
|---|---|---|---|---|---|---|
| 9730 × E-108 | −1.433 NS | −1.390 NS | 0.625 * | −18.077 NS | 0.171 * | 0.789 * |
| 9730 × E-113 | 1.021 NS | 0.229 * | −0.295 NS | 2.811 NS | 0.091 * | 0.215 * |
| 9730 × E-114 | 0.413 * | 1.160 * | −0.329 NS | 1.153 * | −0.262 NS | −1.004 NS |
| 9731 × E-108 | −0.233 * | 34.466 NS | −0.488 NS | 1.522 ** | 0.211 * | −1.956 NS |
| 9731 × E-113 | 1.221 NS | −5.265 NS | −1.408 NS | 0.527 * | −0.089 NS | 0.904 * |
| 9731 × E-114 | −0.987 NS | −29.201 NS | 1.895 NS | −13.108 NS | −0.122 NS | 1.052 * |
| 9733 × E-108 | −2.791 NS | −25.251 NS | −0.358 NS | −8.589 NS | −0.118 NS | −3.256 NS |
| 9733 × E-113 | 2.700 NS | 47.118 NS | 0.882 ** | 0.297 * | −0.024 NS | −0.130 NS |
| 9733 × E-114 | 0.091 * | −21.867 NS | −0.525 NS | 4.840 NS | 0.142 * | 3.385 NS |
| 9859 × E-108 | −0.840 NS | −18.678 NS | −1.194 NS | 1.425 ** | 0.088 * | 1.911 NS |
| 9859 × E-113 | −2.313 NS | 5.473 * | 2.002 NS | 1.939 NS | 0.082 * | −0.896 NS |
| 9859 × E-114 | 3.153 NS | 13.205 * | −0.808 NS | 3.440 NS | −0.171 NS | −1.015 NS |
| 9860 × E-108 | 1.604 NS | 2.194 * | 1.660 NS | 0.767 * | −0.232 NS | 0.878 * |
| 9860 × E-113 | −2.721 NS | −14.754 NS | −0.480 NS | −3.878 NS | −0.004 NS | 1.470 NS |
| 9860 × E-114 | 1.117 NS | 12.560 * | −1.180 NS | 3.625 NS | 0.236 * | −2.348 NS |
| 9861 × E-108 | −5.180 NS | −52.673 NS | −0.080 NS | 84.000 NS | 0.202 * | 1.700 NS |
| 9861 × E-113 | 0.487 * | −4.354 NS | −0.333 NS | −9.800 NS | −0.024 NS | 0.026 * |
| 9861 × E-114 | 4.693 NS | 57.027 NS | 0.413 * | −29.697 | −0.178 NS | −1.726 NS |
| AARI-11 × E-108 | 2.558 NS | 20.599 ** | 0.515 * | 0.735 * | 0.011 * | −1.467 NS |
| AARI-11 × E-113 | 1.155 NS | 7.718 * | −1.409 NS | 2.047 NS | −0.315 NS | −2.974 NS |
| AARI-11 × E-114 | −3.713 NS | −28.317 ** | 0.894 ** | 1.454 ** | 0.305 ** | 4.441 * |
| PB-11 × E-108 | 0.860 NS | 2.522 * | −1.452 NS | −13.591 NS | −0.034 NS | 1.867 NS |
| PB-11 × E-113 | −0.126 NS | 26.073 NS | 0.965 ** | −0.149 NS | 0.160 * | −0.441 NS |
| PB-11 × E-114 | −0.734 NS | −28.595 NS | 0.487 * | 0.850 * | −0.126 NS | −1.426 NS |
| AAS-11 × E-108 | 5.456 NS | 38.210 NS | 0.771 * | −1.545 NS | −0.298 NS | −0.467 NS |
| AAS-11 × E-113 | −1.424 NS | −62.238 NS | 0.075 * | −2.097 NS | 0.122 * | 1.826 NS |
| AAS-11 × E-114 | −4.032 NS | 24.027 NS | −0.846 NS | −1.108 NS | 0.176 * | −1.359 NS |
| S.E for crosses | 0.341 | 11.634 | 0.487 | 0.796 | 0.163 | 0.702 |
| Genetic Variation | Days to 50% Maturity | Growing Degree Days | Productive Tillers per Plant | Spike Density | Grain Weight per Spike | ×1000-Grain Weight |
|---|---|---|---|---|---|---|
| Variance of GCA | 3.336 | 933.060 | 0.213 | 1.002 | 0.017 | 3.880 |
| Variance of SCA | 10.395 | 1182.578 | 1.386 | 7.122 | 0.024 | 5.203 |
| Additive variance | 6.671 | 1866.121 | 0.427 | 1.002 | 0.034 | 7.761 |
| Dominance variance | 10.395 | 1182.578 | 1.386 | 7.122 | 0.024 | 5.203 |
| Variance ratio of GCA to SCA | 0.321 | 0.789 | 0.154 | 0.141 | 0.714 | 0.746 |
| Degree of dominance | 1.248 | 0.796 | 1.802 | 2.666 | 0.837 | 0.819 |
| DTM | GDD | Tp−1 | SD | GWTsp−1 | ×1000 GWT | |
|---|---|---|---|---|---|---|
| Ve | 0.3 | 406.1 | 0.7 | 0.0067 | 0.1 | 1.5 |
| Vg | 10.787 | 2708.557 | 1.124667 | 0.006907 | 0.038667 | 15.149 |
| Vp | 11.136 | 3114.629 | 1.836667 | 0.013585 | 0.118667 | 16.626 |
| H2 | 0.96866 | 0.869624 | 0.612341 | 0.508431 | 0.325843 | 0.911163 |
| GA | 5.689168 | 85.41762 | 1.460566 | 0.104299 | 0.197554 | 6.538871 |
| CV% | 0.5 | 1.3 | 9.7 | 5.0 | 10 | 2.7 |
| Phenotypic Correlation | Genotypic Correlation | |||||
|---|---|---|---|---|---|---|
| DTM | GDD | Tp−1 | SD | GWTsp−1 | ×1000 GWT | |
| DTM | 1 ** | 0.45125 ** | 0.08029 NS | 0.2902 ** | −0.19064 * | −0.0225 NS |
| GDD | 0.40744 ** | 1 ** | 0.13473 NS | 0.25388 ** | −0.18587 * | −0.28906 ** |
| Tp−1 | 0.05706 NS | 0.13118 NS | 1 ** | −0.1084 NS | −0.31496 ** | −0.02665 NS |
| SD | 0.20942 * | 0.19091 * | −0.17791 * | 1 ** | −0.46559 ** | −0.01083 NS |
| GWTsp−1 | −0.10889 NS | −0.08238 NS | −0.16573 NS | −0.24462 ** | 1 ** | 0.2847 ** |
| ×1000 GWT | −0.01657 NS | −0.27514 ** | −0.01835 NS | −0.01083 NS | 0.20208 ** | 1 ** |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farooq, M.U.; Khan, A.S.; Ishaaq, I.; Cheema, A.A.; Afzal, M.S.; Ali, A.; Zhu, J. Growing Degree Days during the Late Reproductive Phase Determine Spike Density and Cognate Yield Traits. Agronomy 2018, 8, 217. https://doi.org/10.3390/agronomy8100217
Farooq MU, Khan AS, Ishaaq I, Cheema AA, Afzal MS, Ali A, Zhu J. Growing Degree Days during the Late Reproductive Phase Determine Spike Density and Cognate Yield Traits. Agronomy. 2018; 8(10):217. https://doi.org/10.3390/agronomy8100217
Chicago/Turabian StyleFarooq, Muhammad Umer, Abdus Salam Khan, Iqra Ishaaq, Asim Ali Cheema, Muhammad Shahzad Afzal, Asif Ali, and Jianqing Zhu. 2018. "Growing Degree Days during the Late Reproductive Phase Determine Spike Density and Cognate Yield Traits" Agronomy 8, no. 10: 217. https://doi.org/10.3390/agronomy8100217
APA StyleFarooq, M. U., Khan, A. S., Ishaaq, I., Cheema, A. A., Afzal, M. S., Ali, A., & Zhu, J. (2018). Growing Degree Days during the Late Reproductive Phase Determine Spike Density and Cognate Yield Traits. Agronomy, 8(10), 217. https://doi.org/10.3390/agronomy8100217