Genetic Diversity of Diurnal Carbohydrate Accumulation in White Clover (Trifolium repens L.)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Quantitation of Glucose, Fructose, Sucrose, Starch and Biomass
2.3. Statistical Analysis of Phenotypic Data
2.4. Iodine Staining of Starch in Plants
3. Results
3.1. Phenotypic Variation of Glucose, Fructose, Sucrose, and Starch Content in White Clover Leaves
3.2. Influence of Glasshouse Environment on Leaf Glucose, Fructose, Sucrose, and Starch Contents
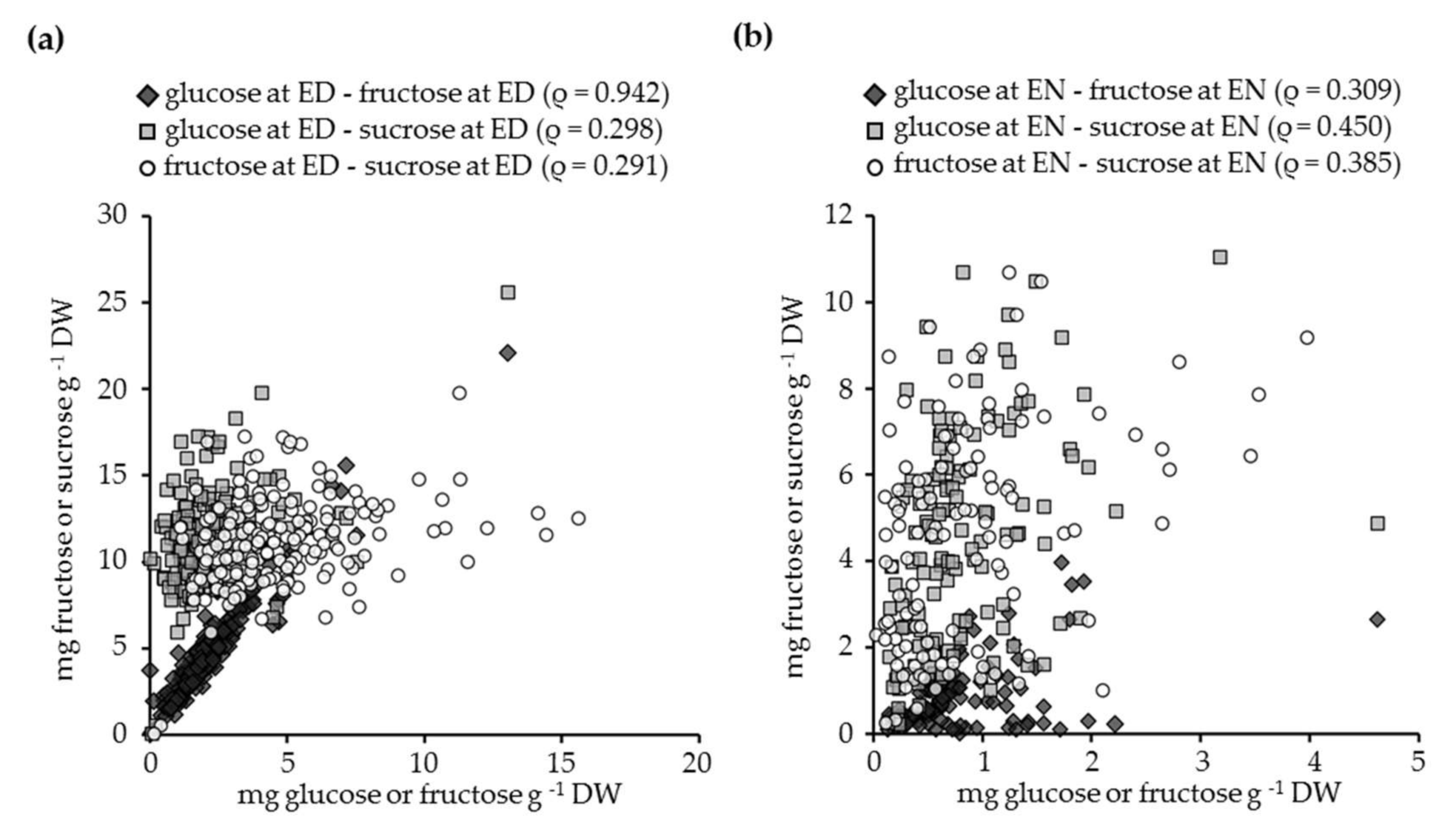
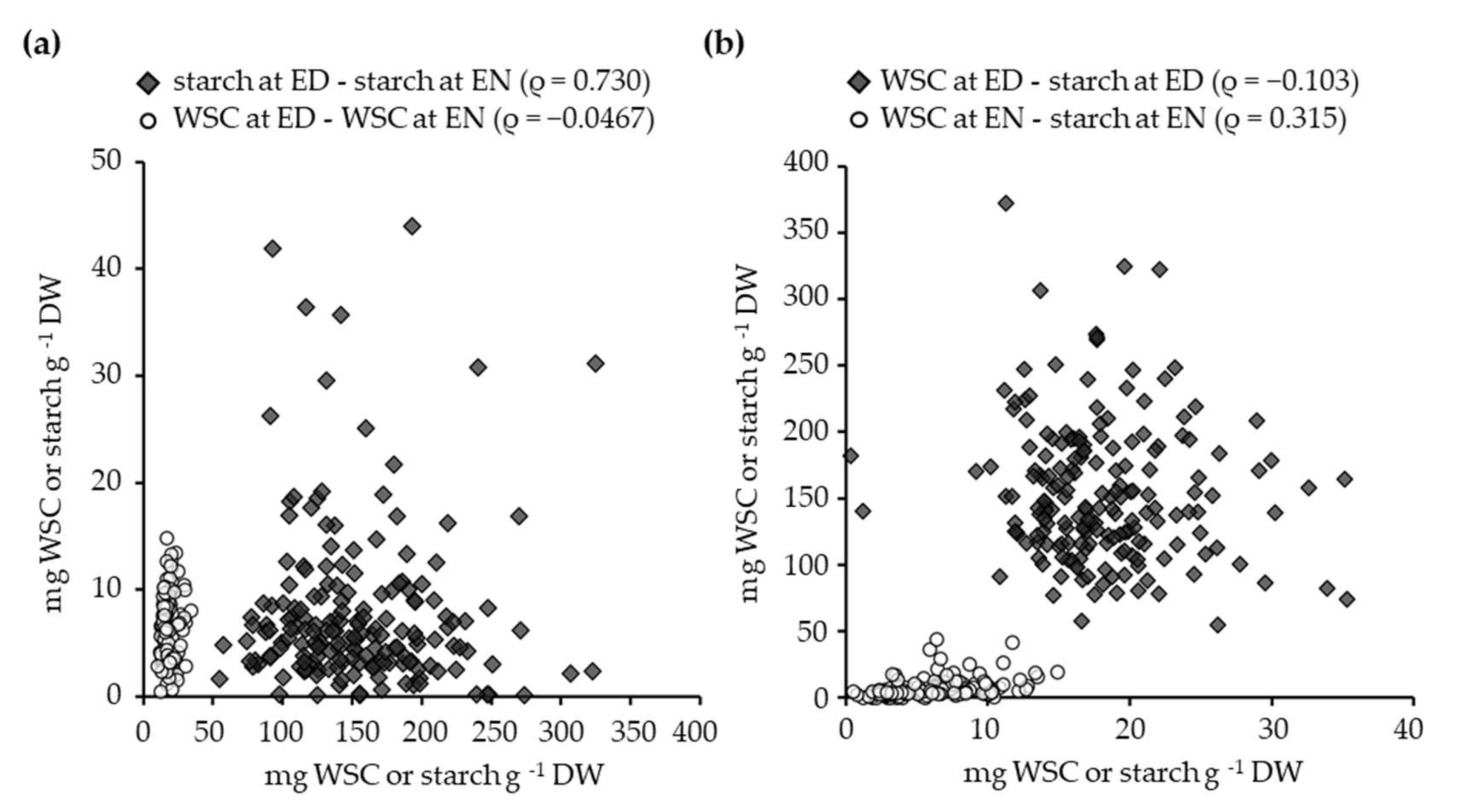
3.3. The Relationship between Glucose, Fructose, Sucrose, and Starch Contents in White Clover Leaves
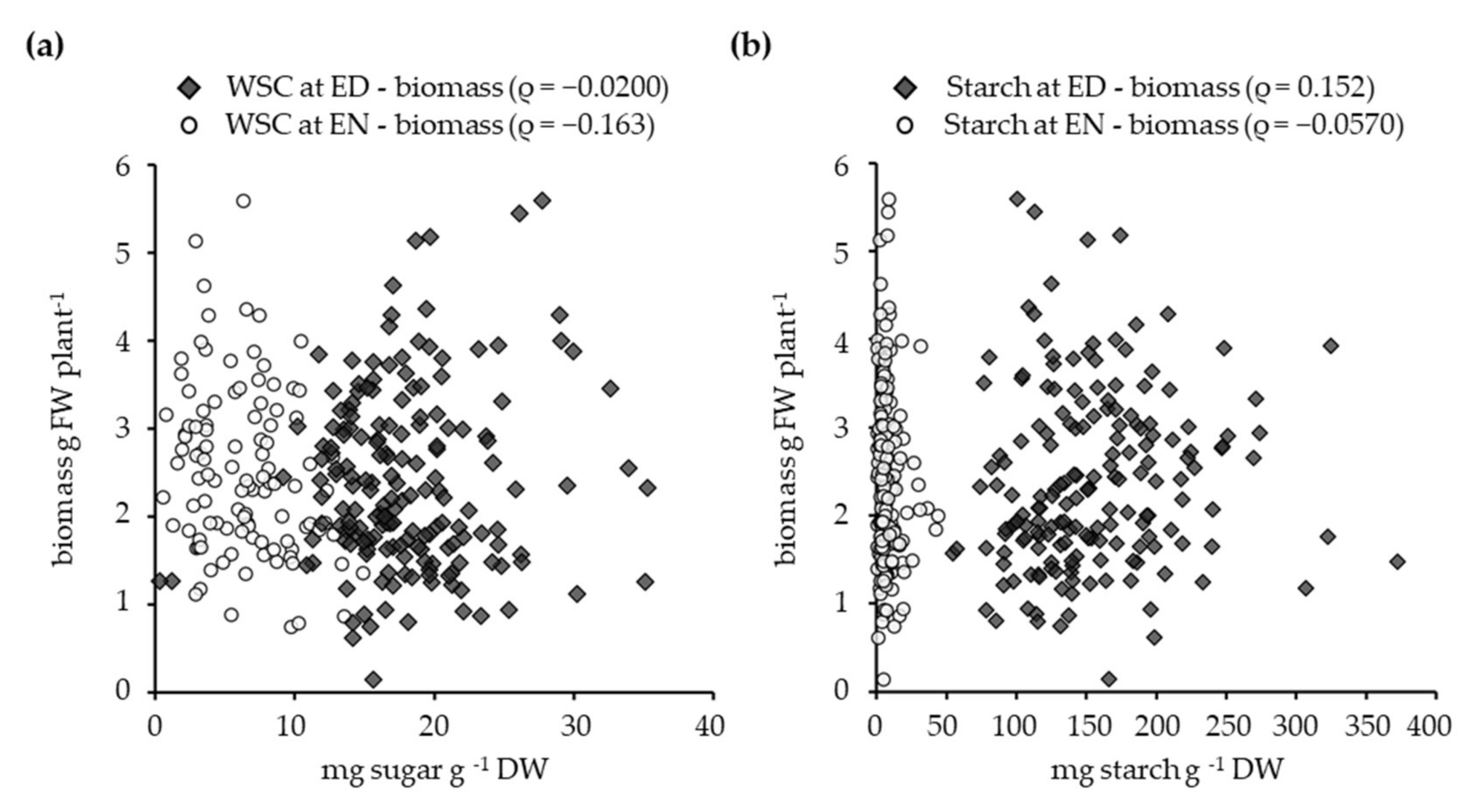
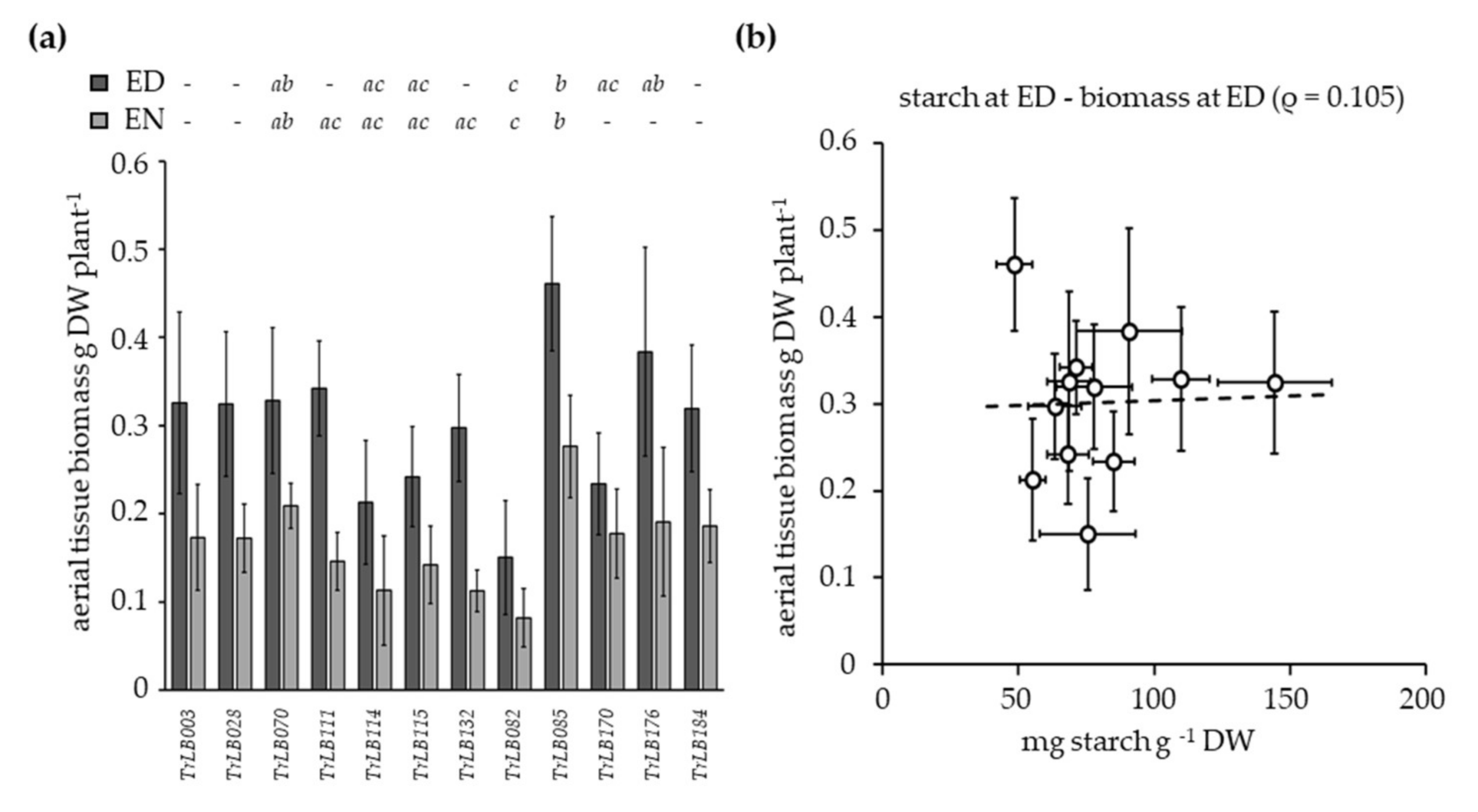
3.4. The Relationship between Leaf Biomass, Water-Soluble Carbohydrates, and Starch Contents
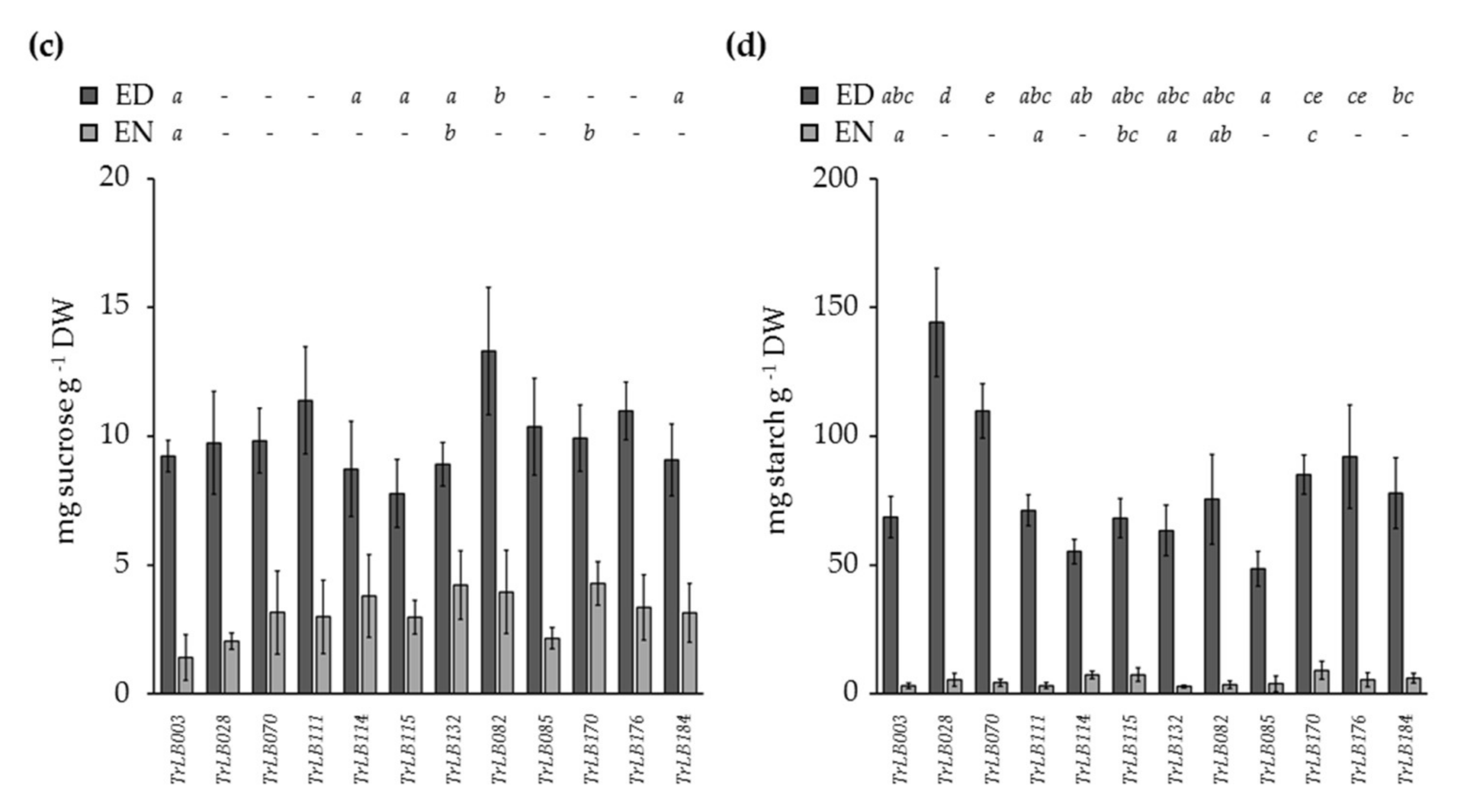
3.5. Genetic Variation in Leaf Diel Leaf Glucose, Fructose, Sucrose, and Starch Contents
3.6. The Pattern of Starch Content in White Clover Leaves
4. Discussion
4.1. Defining NSC Content as a Trait in White Clover
4.2. Toward the Development of a Field Trait
4.3. Relationship between Leaf NSC Content, Plant Growth, and Whole Plant Carbohydrate Homeostasis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Robertson, G.P.; Vitousek, P.M. Nitrogen in agriculture: Balancing the cost of an essential resource. Annu. Rev. Environ. Resour. 2009, 34, 97–125. [Google Scholar] [CrossRef]
- Paustian, K.; Lehmann, J.; Ogle, S.; Reay, D.; Robertson, G.P.; Smith, P. Climate-smart soils. Nature 2016, 532, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Steinfeld, H.; Gerber, P.; Wassenaar, T.; Castel, V.; de Haan, C. Livestock’s Long Shadow: Environmental Issues and Options; Food & Agriculture Organization: Rome, Italy, 2006. [Google Scholar]
- Kingston-Smith, A.; Marshall, A.; Moorby, J. Breeding for genetic improvement of forage plants in relation to increasing animal production with reduced environmental footprint. Animal 2013, 7, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Nocek, J.E.; Tamminga, S. Site of digestion of starch in the gastrointestinal tract of dairy cows and its effect on milk yield and composition. J. Dairy Sci. 1991, 74, 3598–3629. [Google Scholar] [CrossRef]
- Zeeman, S.C.; Kossmann, J.; Smith, A.M. Starch: Its metabolism, evolution, and biotechnological modification in plants. Annu. Rev. Plant Biol. 2010, 61, 209–234. [Google Scholar] [CrossRef] [PubMed]
- Stitt, M.; Zeeman, S.C. Starch turnover: Pathways, regulation and role in growth. Curr. Opin. Plant Biol. 2012, 15, 282–292. [Google Scholar] [CrossRef] [PubMed]
- Vijn, I.; Smeekens, S. Fructan: More than a reserve carbohydrate? Plant Physiol. 1999, 120, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Turner, L.; Cairns, A.; Armstead, I.; Ashton, J.; Skøt, K.; Whittaker, D.; Humphreys, M. Dissecting the regulation of fructan metabolism in perennial ryegrass (Lolium perenne) with quantitative trait locus mapping. New Phytol. 2006, 169, 45–58. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, A.; Dhont, C.; Bipfubusa, M.; Chalifour, F.-P.; Drouin, P.; Beauchamp, C.J. Improving salt stress responses of the symbiosis in alfalfa using salt-tolerant cultivar and rhizobial strain. Appl. Soil Ecol. 2015, 87, 108–117. [Google Scholar] [CrossRef]
- Brito, A.; Tremblay, G.; Bertrand, A.; Castonguay, Y.; Bélanger, G.; Michaud, R.; Lafrenière, C.; Martineau, R.; Berthiaume, R. Alfalfa baleage with increased concentration of nonstructural carbohydrates supplemented with a corn-based concentrate did not improve production and nitrogen utilization in early lactation dairy cows. J. Dairy Sci. 2014, 97, 6970–6990. [Google Scholar] [CrossRef] [PubMed]
- Turner, L.; Pollock, C. Changes in stolon carbohydrates during the winter in four varieties of white clover (Trifolium repens L.) with contrasting hardiness. Ann. Bot. 1998, 81, 97–107. [Google Scholar] [CrossRef]
- Ruckle, M.E.; Meier, M.A.; Frey, L.; Eicke, S.; Kölliker, R.; Zeeman, S.C.; Studer, B. Diurnal leaf starch content: An orphan trait in forage legumes. Agronomy 2017, 7, 16. [Google Scholar] [CrossRef]
- Claessens, A.; Castonguay, Y.; Bertrand, A.; Bélanger, G.; Tremblay, G. Breeding for improved nonstructural carbohydrates in alfalfa. In Breeding in a World of Scarcity; Springer International Publishing Switzerland: Cham, Switzerland, 2016; pp. 231–235. [Google Scholar]
- Boller, B.C.; Nösberger, J. Effects of temperature and photoperiod on stolon characteristics, dry matter partitioning, and nonstructural carbohydrate concentration of two white clover ecotypes. Crop Sci. 1983, 23, 1057–1062. [Google Scholar] [CrossRef]
- Humphreys, M. Water-soluble carbohydrates in perennial ryegrass breeding. Grass Forage Sci. 1989, 44, 237–244. [Google Scholar] [CrossRef]
- Moorby, J.; Evans, R.; Scollan, N.; MacRae, J.; Theodorou, M. Increased concentration of water-soluble carbohydrate in perennial ryegrass (Lolium perenne L.). Evaluation in dairy cows in early lactation. Grass Forage Sci. 2006, 61, 52–59. [Google Scholar] [CrossRef]
- Livingston, D.P.; Hincha, D.K.; Heyer, A.G. Fructan and its relationship to abiotic stress tolerance in plants. Cell. Mol. Life Sci. 2009, 66, 2007–2023. [Google Scholar] [CrossRef] [PubMed]
- Elgersma, A.; Hassink, J. Effects of white clover (Trifolium repens L.) on plant and soil nitrogen and soil organic matter in mixtures with perennial ryegrass (Lolium perenne L.). Plant Soil 1997, 197, 177–186. [Google Scholar] [CrossRef]
- Lowell, M.E. Post-Harvest Physiological Changes in Forage Plants; Post-Harvest Physiology and Preservation of Forages; Crop Science Society of America: Madison, WI, USA, 1995; pp. 1–19. [Google Scholar]
- Wilkinson, J. Losses in the conservation and utilisation of grass and forage crops. Ann. Appl. Biol. 1981, 98, 365–375. [Google Scholar] [CrossRef]
- Orr, R.J.; Penning, P.D.; Harvey, A.; Champion, R.A. Diurnal patterns of intake rate by sheep grazing monocultures of ryegrass or white clover. Appl. Anim. Behav. Sci. 1997, 52, 65–77. [Google Scholar] [CrossRef]
- Lüscher, A.; Mueller-Harvey, I.; Soussana, J.F.; Rees, R.M.; Peyraud, J.L. Potential of legume-based grassland–livestock systems in europe: A review. Grass Forage Sci. 2014, 69, 206–228. [Google Scholar] [CrossRef] [PubMed]
- Williams, T.; Abberton, M.; Rhodes, I. Performance of white clover varieties combined in blends and alone when grown with perennial ryegrass under sheep and cattle grazing. Grass Forage Sci. 2003, 58, 90–93. [Google Scholar] [CrossRef]
- Nyfeler, D.; Huguenin-Elie, O.; Suter, M.; Frossard, E.; Connolly, J.; Lüscher, A. Strong mixture effects among four species in fertilized agricultural grassland led to persistent and consistent transgressive overyielding. J. Appl. Ecol. 2009, 46, 683–691. [Google Scholar] [CrossRef]
- Gregorini, P. Diurnal grazing pattern: Its physiological basis and strategic management. Anim. Prod. Sci. 2012, 52, 416–430. [Google Scholar] [CrossRef]
- Nkrumah, J.; Okine, E.; Mathison, G.; Schmid, K.; Li, C.; Basarab, J.; Price, M.; Wang, Z.; Moore, S. Relationships of feedlot feed efficiency, performance, and feeding behavior with metabolic rate, methane production, and energy partitioning in beef cattle. J. Anim. Sci. 2006, 84, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Thalmann, M.; Santelia, D. Starch as a determinant of plant fitness under abiotic stress. New Phytol. 2017, 214, 943–951. [Google Scholar] [CrossRef] [PubMed]
- Hostettler, C.; Kölling, K.; Santelia, D.; Streb, S.; Kötting, O.; Zeeman, S.C. Analysis of starch metabolism in chloroplasts. In Chloroplast Research in Arabidopsis: Methods and Protocols, Volume II; Jarvis, R.P., Ed.; Humana Press: Totowa, NJ, USA, 2011; pp. 387–410. [Google Scholar]
- Zhang, N.; Gibon, Y.; Jason, G.W.; Nick, K.L.; Li, P.; Dedow, L.; Chen, C.; So, Y.-S.; Kremling, K.; Bradbury, P. Genome-wide association of carbon and nitrogen metabolism in the maize nested association mapping population. Plant Physiol. 2015, 168, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Boller, B.; Tanner, P. Sortenblatt Munida Weissklee (4n): Trifolium Repens l. Ertragsstark und Sehr Grossblättrig; Hrsg. Agroscope INH: Zürich, Switzerland, 2014; Volume 1. [Google Scholar]
- Team, R.C. The R Project for Statistical Computing. Available online: www.r-project.org/ (accessed on 3 February 2015).
- Studer, B.; Boller, B.; Bauer, E.; Posselt, U.K.; Widmer, F.; Kölliker, R. Consistent detection of qtls for crown rust resistance in italian ryegrass (Lolium multiflorum Lam.) across environments and phenotyping methods. Theor. Appl. Genet. 2007, 115, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, B.B.; Gruissem, W.; Jones, R.L. Biochemistry & Molecular Biology of Plants; American Society of Plant Physiologists: Rockville, MD, USA, 2000; Volume 40. [Google Scholar]
- Ruan, Y.-L. Sucrose metabolism: Gateway to diverse carbon use and sugar signaling. Annu. Rev. Plant Biol. 2014, 65, 33–67. [Google Scholar] [CrossRef] [PubMed]
- Sturm, A.; Tang, G.-Q. The sucrose-cleaving enzymes of plants are crucial for development, growth and carbon partitioning. Trends Plant Sci. 1999, 4, 401–407. [Google Scholar] [CrossRef]
- Smith, A.M.; Stitt, M. Coordination of carbon supply and plant growth. Plant Cell Environ. 2007, 30, 1126–1149. [Google Scholar] [CrossRef] [PubMed]
- Gibon, Y.; Usadel, B.; Blaesing, O.E.; Kamlage, B.; Hoehne, M.; Trethewey, R.; Stitt, M. Integration of metabolite with transcript and enzyme activity profiling during diurnal cycles in arabidopsis rosettes. Genome Biol. 2006, 7, R76. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.M. Starch in the arabidopsis plant. Starch-Stärke 2012, 64, 421–434. [Google Scholar] [CrossRef]
- Cabrera-Bosquet, L.; Fournier, C.; Brichet, N.; Welcker, C.; Suard, B.; Tardieu, F. High-throughput estimation of incident light, light interception and radiation-use efficiency of thousands of plants in a phenotyping platform. New Phytol. 2016, 212, 269–281. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, J.R.; Kossmann, J.; Ritte, G. Leaf starch degradation comes out of the shadows. Trends Plant Sci. 2005, 10, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Masclaux, C.; Valadier, M.-H.; Brugière, N.; Morot-Gaudry, J.-F.; Hirel, B. Characterization of the sink/source transition in tobacco (Nicotiana tabacum L.) shoots in relation to nitrogen management and leaf senescence. Planta 2000, 211, 510–518. [Google Scholar] [CrossRef] [PubMed]
- Mondal, M.H.; Brun, W.A.; Brenner, M.L. Effects of sink removal on photosynthesis and senescence in leaves of soybean (Glycine max L.) plants. Plant Physiol. 1978, 61, 394–397. [Google Scholar] [CrossRef] [PubMed]
- Pfister, B.; Lu, K.-J.; Eicke, S.; Feil, R.; Lunn, J.E.; Streb, S.; Zeeman, S.C. Genetic evidence that chain length and branch point distributions are linked determinants of starch granule formation in arabidopsis. Plant Physiol. 2014, 165, 1457–1474. [Google Scholar] [CrossRef] [PubMed]
- Tardieu, F.; Tuberosa, R. Dissection and modelling of abiotic stress tolerance in plants. Curr. Opin. Plant Biol. 2010, 13, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Araus, J.L.; Cairns, J.E. Field high-throughput phenotyping: The new crop breeding frontier. Trends Plant Sci. 2014, 19, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Manzanares, C.; Yates, S.; Ruckle, M.; Nay, M.; Studer, B. Tilling in forage grasses for gene discovery and breeding improvement. New Biotechnol. 2016, 33, 594–603. [Google Scholar] [CrossRef] [PubMed]
- Gibon, Y.; Blaesing, O.E.; Hannemann, J.; Carillo, P.; Höhne, M.; Hendriks, J.H.; Palacios, N.; Cross, J.; Selbig, J.; Stitt, M. A robot-based platform to measure multiple enzyme activities in arabidopsis using a set of cycling assays: Comparison of changes of enzyme activities and transcript levels during diurnal cycles and in prolonged darkness. Plant Cell 2004, 16, 3304–3325. [Google Scholar] [CrossRef] [PubMed]
- Dreccer, M.F.; Barnes, L.R.; Meder, R. Quantitative dynamics of stem water soluble carbohydrates in wheat can be monitored in the field using hyperspectral reflectance. Field Crops Res. 2014, 159, 70–80. [Google Scholar] [CrossRef]
- Yendrek, C.; Tomaz, T.; Montes, C.M.; Cao, Y.; Morse, A.M.; Brown, P.J.; McIntyre, L.; Leakey, A.; Ainsworth, E. High-throughput phenotyping of maize leaf physiology and biochemistry using hyperspectral reflectance. Plant Physiol. 2016, 176. [Google Scholar] [CrossRef]
- Sulpice, R.; Pyl, E.-T.; Ishihara, H.; Trenkamp, S.; Steinfath, M.; Witucka-Wall, H.; Gibon, Y.; Usadel, B.; Poree, F.; Piques, M.C. Starch as a major integrator in the regulation of plant growth. Proc. Natl Acad. Sci. USA 2009, 106, 10348–10353. [Google Scholar] [CrossRef] [PubMed]
- Vriet, C.; Welham, T.; Brachmann, A.; Pike, M.; Pike, J.; Perry, J.; Parniske, M.; Sato, S.; Tabata, S.; Smith, A.M. A suite of lotus japonicus starch mutants reveals both conserved and novel features of starch metabolism. Plant Physiol. 2010, 154, 643–655. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| End of Day | End of Night | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Glucose | Fructose | Sucrose | Starch | Total NSC | Biomass | Glucose | Fructose | Sucrose | Starch | Total NSC | Biomass | |
| Mean | 2.01 | 3.09 | 9.86 | 79.9 | 94.8 | 0.298 | 2.16 | 1.38 | 3.12 | 5.28 | 15.1 | 0.165 |
| Genotypic Variance | 2.51 | 6.91 | 15.6 | 4830 | 4690 | 0.0429 | 1.65 | 0.792 | 6.03 | 37.2 | 33.8 | 0.0182 |
| Repeatability | 0.824 | 0.867 | 0.753 | 0.946 | 0.94 | 0.796 | 0.591 | 0.646 | 0.709 | 0.799 | 0.76 | 0.805 |
| F-value | 4.70 * | 6.51 * | 3.04 * | 17.4 * | 15.6 * | 3.91 * | 1.44 | 1.70 | 2.44 | 3.97 * | 3.24 * | 4.13 * |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruckle, M.E.; Bernasconi, L.; Kölliker, R.; Zeeman, S.C.; Studer, B. Genetic Diversity of Diurnal Carbohydrate Accumulation in White Clover (Trifolium repens L.). Agronomy 2018, 8, 47. https://doi.org/10.3390/agronomy8040047
Ruckle ME, Bernasconi L, Kölliker R, Zeeman SC, Studer B. Genetic Diversity of Diurnal Carbohydrate Accumulation in White Clover (Trifolium repens L.). Agronomy. 2018; 8(4):47. https://doi.org/10.3390/agronomy8040047
Chicago/Turabian StyleRuckle, Michael E., Lucia Bernasconi, Roland Kölliker, Samuel C. Zeeman, and Bruno Studer. 2018. "Genetic Diversity of Diurnal Carbohydrate Accumulation in White Clover (Trifolium repens L.)" Agronomy 8, no. 4: 47. https://doi.org/10.3390/agronomy8040047
APA StyleRuckle, M. E., Bernasconi, L., Kölliker, R., Zeeman, S. C., & Studer, B. (2018). Genetic Diversity of Diurnal Carbohydrate Accumulation in White Clover (Trifolium repens L.). Agronomy, 8(4), 47. https://doi.org/10.3390/agronomy8040047