Improving the Conservation and Use of Traditional Germplasm through Breeding for Local Adaptation: The Case of the Castellfollit del Boix Common Bean (Phaseolus vulgaris L.) Landrace

 ,
, 
 and
and
Abstract
:1. Introduction
2. Materials and Methods
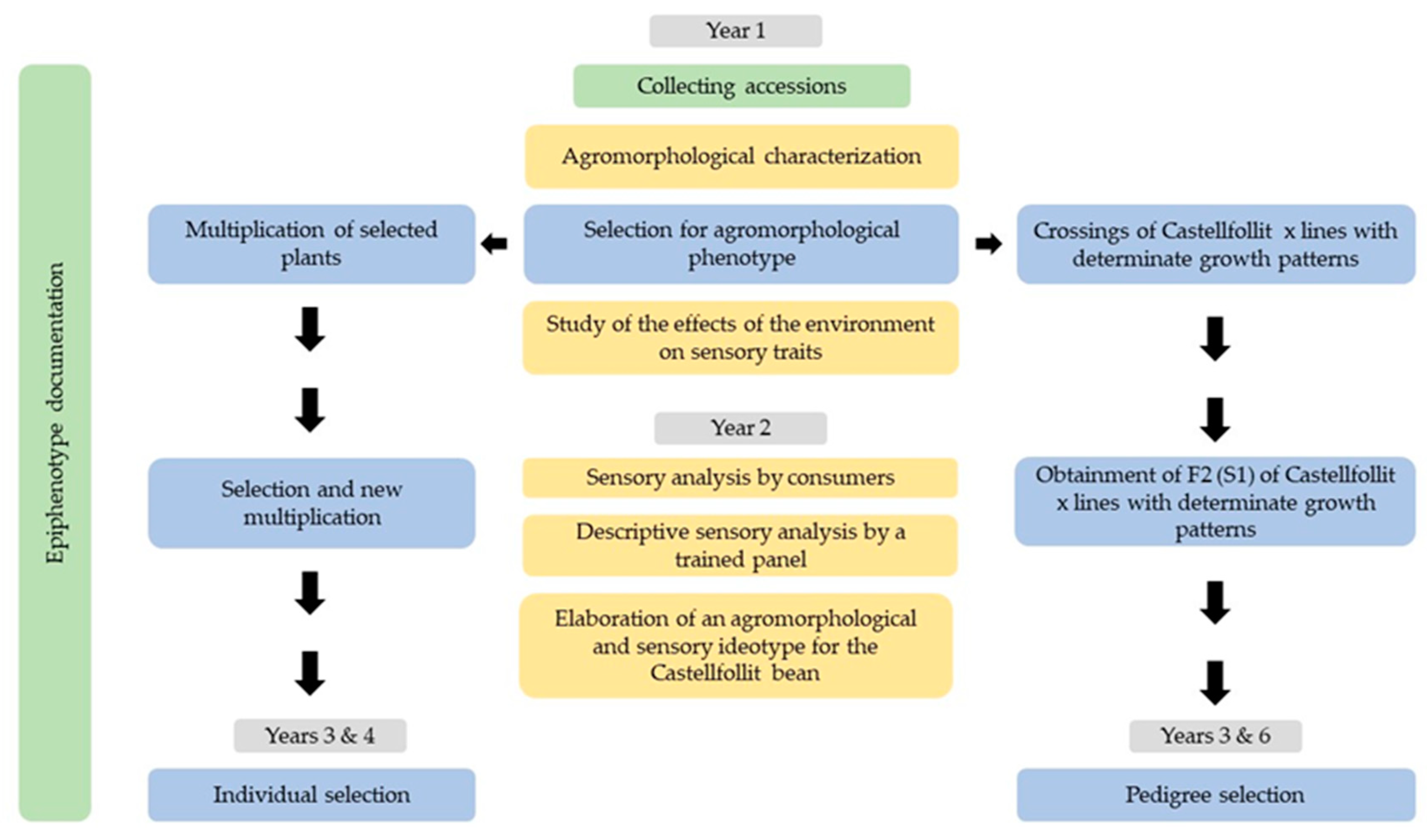
2.1. The Castellfollit del Boix Breeding Project
2.2. Collecting Accessions and Characterizing the Epiphenotype
2.3. From Characterizing the Phenotypes to the Elaboration of the Ideotype
2.3.1. Characterizing the Phenotypes
2.3.2. Effects of the Environment on Sensory Traits
2.3.3. Consumer Taste Test
2.3.4. Elaboration of the Ideotype
2.4. Selection, Crossings, and Breeding Program
2.4.1. Individual Selection Program
2.4.2. Pedigree Selection
2.5. Statistical Analysis of the Data and Index to Measure Conservation Status and Conditions Favorable for Conservation
2.5.1. Statistical Analyses
2.5.2. Measuring Conservation Status and Conditions Favorable for Conservation
3. Results
3.1. Collecting Accessions, Phenotypic and Epiphenotypic Characterization, Phenotypic Selection, Study of Preferences, and Elaboration of the Ideotype
3.2. Selection
3.2.1. Individual
3.2.2. Pedigree Selection
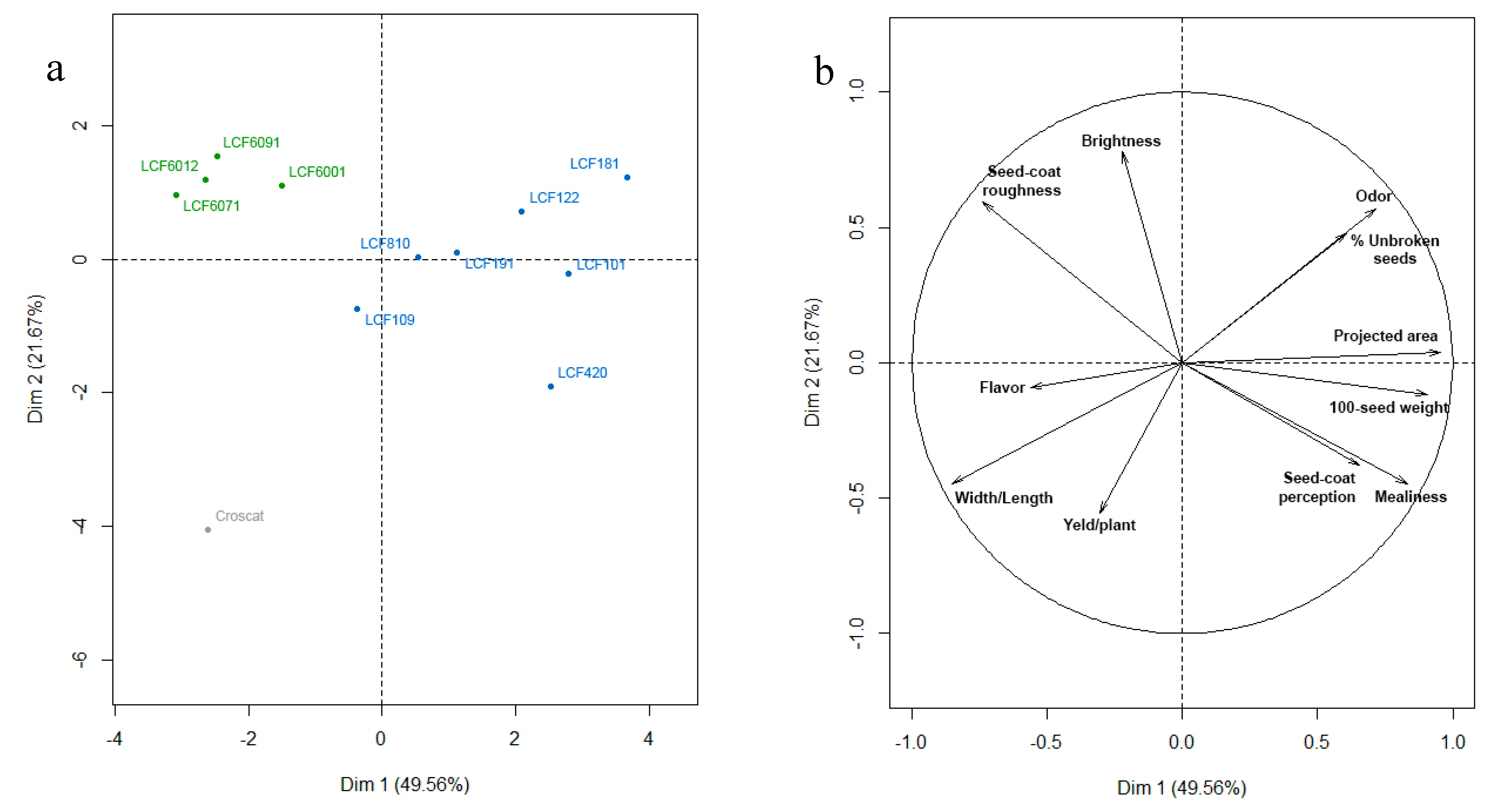
3.2.3. The Selected Germplasm
3.3. Effects on Conservation and the Predisposition to Conservation during the Intervention Process
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Louwaars, N.P. Plant breeding and diversity: A troubled relationship? Euphytica 2018, 214, 114–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borlaug, N.E. The green revolution revisited and the road ahead. In Proceedings of the Special 30th Anniversary Lecture, Oslo, Norway, 8 September 2000. [Google Scholar]
- Gollin, D.; Morris, M.; Ekbyerlee, A. Technology adoption in intensive post-green revolution systems. Am. J. Agric. Econ. 2005, 87, 1310–1316. [Google Scholar] [CrossRef]
- Stout, J. Handbook of Energy for World Agriculture; Elsevier: New York, NY, USA, 1990. [Google Scholar]
- Casañas, F.; Simó, J.; Casals, J.; Prohens, J. Towards an evolved concept of landrace. Front. Plant Sci. 2017, 8, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO. The Second Report on the State of the World’s Plant Genetic Resources for Food and Agriculture; FAO: Rome, Italy, 2010. [Google Scholar]
- Sánchez, E.; Sifres, A.; Casañas, F.; Nuez, F. Common bean (Phaseolus vulgaris L.) landraces in Catalonia, a Mesoamerican germplasm hotspot to be preserved. J. Hortic. Sci. Biotech. 2007, 82, 529–534. [Google Scholar] [CrossRef]
- Young, A. Tour in Catalonia. In Annals of Agriculture and Other Useful Arts; Arthur Young: London, UK, 1787. [Google Scholar]
- Singh, S.P. A key for identification of different growth habits of Phaseolus vulgaris L. Annu. Rep. Bean Improv. Coop. 1982, 25, 92–95. [Google Scholar]
- Díez, M.J.; de la Rosa, L.; Martín, I.; Guasch, L.; Cartea, M.E.; Mallor, C.; Casals, J.; Simó, J.; Rivera, A.; Anastasio, G.; et al. Plant Genebanks: Present Situation and Proposals for Their Improvement. The Case of the Spanish Network. Front. Plant Sci. 2018, 9, 1794. [Google Scholar] [CrossRef] [Green Version]
- ISO. 8589 Sensory Analysis—General Guidance for the Design of Test Rooms; International Organization for Standardization: Génève, Switzerland, 2007. [Google Scholar]
- Romero del Castillo, R.; Costell, E.; Plans, M.; Simó, J.; Casañas, F. A standardized method of preparing common beans (Phaseolus vulgaris L.) for sensory analysis. J. Sens. Stud. 2012, 27, 188–195. [Google Scholar] [CrossRef]
- Romero del Castillo, R.; Valero, J.; Casañas, F.; Costell, E. Training, validation and maintenance of a panel to evaluate the texture of dry beans (Phaseolus vulgaris L.). J. Sens. Stud. 2008, 23, 303–319. [Google Scholar] [CrossRef]
- Meilgaard, M.C.; Civille, G.V.; Carr, B.T. Sensory Evaluation Techniques, 4th ed.; CRC Press: Boca Raton, FL, USA, 2007. [Google Scholar]
- Dawson, J.C.; Murphy, K.M.; Jones, S. Decentralized selection and participatory approaches in plant breeding for low-input systems. Euphytica 2008, 160, 143–154. [Google Scholar] [CrossRef]
- Almirall, A.; Bosch, L.; Romero del Castillo, R.; Rivera, A.; Casañas, F. ‘Croscat’ common bean (Phaseolus vulgaris L.), a prototypical cultivar within the ‘Tavella Brisa’ type. HortScience 2010, 45, 432–433. [Google Scholar] [CrossRef] [Green Version]
- Plans, M.; Simó, J.; Casañas, F.; Sabaté, J. Near-infrared spectroscopy analysis of seed coats of common beans (Phaseolus vulgaris L.): A potential tool for breeding and quality evaluation. J. Agric. Food Chem. 2012, 60, 706–712. [Google Scholar] [CrossRef] [PubMed]
- Plans, M.; Simó, J.; Casañas, F.; Romero del Castillo, R.; Rodriguez-Saona, L.E.; Sabaté, J. Estimating sensory properties of common beans (Phaseolus vulgaris L.) by near infrared spectroscopy. Food Res. Int. 2014, 56, 55–62. [Google Scholar] [CrossRef]
- Coyne, D.P. Photoperiodism: Inheritance and linkage studies in Phaseolus vulgaris. J. Hered. 1967, 58, 313–314. [Google Scholar] [CrossRef]
- Koinange, E.M.K.; Singh, S.P.; Gepts, P. Genetic control of the domestication syndrome in common-bean. Crop Sci. 1996, 36, 1037–1145. [Google Scholar] [CrossRef] [Green Version]
- O’Mahony, M. Sensory Evaluation of Food. In Statistical Methods and Procedures; Marcel Dekker: New York, NY, USA, 1986. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2010. [Google Scholar]
- Mendiburu, F. Agricolae: Statistical Procedures for Agricultural Research; R Package Version 1.0–9; R Foundation for Statistical Computing: Vienna, Austria, 2010. [Google Scholar]
- FMA. Manual de bones pràctiques. Mongeta de Castellfollit del Boix. 2019. Available online: http://fundaciomiquelagusti.com/es/2017/03/04/projecte-de-recuperacio-i-millora-genetica-de-la-mongeta-de-castellfollit-del-boix/ (accessed on 20 October 2019).
- Sans, S.; Casals, J.; Simó, J. Improving the Commercial Value of the ‘Calçot’ (Allium cepa L.) Landrace: Influence of Genetic and Environmental Factors in Chemical Composition and Sensory Attributes. Front. Plant Sci. 2018, 9, 1465. [Google Scholar] [CrossRef] [PubMed]
- Santalla, M.; de Ron, A.M.; Voysest, O. European Bean Market Classes. In Catalogue of Bean Genetic Resources; Amurrio, M., Santalla, M., de Ron, A.M., Eds.; PHASELIEUFAIRPL97-3463-MBG-CSIC; Fundación Pedro Barrié de la Maza: Pontevedra, Spain, 2001; pp. 79–94. [Google Scholar]
- Casañas, F.; Pujolà, M.; Romero del Castillo, R.R.; Almirall, A.; Sánchez, E.; Nuez, F. Variability in some texture characteristics and chemical composition of common beans (Phaseolus vulgaris L.). J. Sci. Food Agric. 2006, 86, 2445–2449. [Google Scholar] [CrossRef] [Green Version]
- De la Iglesia, B.; Potter, J.F.; Poulter, N.R.; Robins, M.M.; Skinner, J. Performance of the ASSIGN cardiovascular disease risk score on a UK cohort of patients from general practice. Heart 2011, 97, 491–499. [Google Scholar] [CrossRef]
- Kant, A.K. Indexes of overall diet quality: A review. J. Am. Diet. Assoc. 1996, 8, 85–791. [Google Scholar]
- Ross, T.; Summer, J. A simple, spreadsheet-based, food safety risk assessment tool. Int. J. Food Microbiol. 2002, 77, 39–53. [Google Scholar] [CrossRef]
- Bitocchi, E.; Nanni, L.; Rossi, M.; Rau, D.; Bellucci, E.; Giardini, A.; Buonamici, A.; Vendramin, G.G.; Papa, R. Introgression from modern hybrid varieties into landrace populations of maize (Zea mays ssp. mays L. in central Italy. Mol. Ecol. 2009, 18, 603–621. [Google Scholar] [CrossRef]
- Van Heerwaarden, J.; Doebley, J.; Briggs, W.H.; Glaubitz, J.C.; Goodman, M.M.; Sanchez Gonzalez, J.J.; Ross-Ibarra, J. Genetic signals of origin, spread, and introgression in a large sample of maize landraces. Proc. Natl. Acad. Sci. USA 2011, 108, 1088–1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Union. Regulation No 1151/2012 of the European Parliament and of the Council of 21 November 2012 on quality schemes for agricultural products and foodstuffs. Off. J. Eur. Union 2012, 14, 12. [Google Scholar]
- Prada, D. Molecular population genetics and agronomic alleles in seed banks: Searching for a needle in a haystack? J. Exp. Bot. 2009, 60, 2541–2552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University Illinois Press: Urbana, IL, USA, 1963. [Google Scholar]
- Nei, M. Analysis of gene diversity in subdivided populations. Proc. Natl. Acad. Sci. USA 1973, 70, 3321–3323. [Google Scholar] [CrossRef] [Green Version]
- Witcombe, J.R.; Joshi, A.; Joshi, K.D.; Sthapit, B.R. Farmer participatory crop improvement. I. Varietal selection and breeding methods and their impact on biodiversity. Exp. Agric. 1996, 32, 445–460. [Google Scholar] [CrossRef]
- Pimentel, D.; Culliney, T.W.; Buttler, I.W.; Reinemann, D.J.; Beckman, K.B. Low-input sustainable agriculture using ecological management practices. Agric. Ecosyst. Environ. 1989, 27, 3–24. [Google Scholar] [CrossRef]
- Lammerts van Bueren, E.T.; Østergård, H.; Goldringer, I.; Scholten, O. Plant breeding for organic and sustainable, low-input agriculture: Dealing with genotype–environment interactions. Euphytica 2008, 163, 321–322. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Items | Definition |
|---|---|
| Controlled multiplication of germplasm | Degree of multiplication of germplasm done in an isolated and controlled way, with conservative selection |
| Morphological characterization | Degree of characterization of the morphological variability found in the landrace |
| Agronomic characterization | Degree of characterization of the agronomic variability found in the landrace |
| Sensory characterization | Degree of characterization of the sensory variability found in the landrace |
| Chemical characterization | Degree of characterization of the chemical variability found in the landrace |
| Genetic characterization | Degree of characterization of the genetic variability found in the landrace |
| Characterization of environmental effects | Degree of characterization of the effects of the environment on the variability found in the landrace |
| Edaphic description | Degree of characterization of the soils in the territory where the landrace has traditionally been cultivated |
| Climatic description | Degree of characterization of the climate in the territory where the landrace has traditionally been cultivated |
| Historical/cultural documentation | Degree of historical/cultural characterization of the uses of the landrace |
| Documentation of traditional cultivation methods | Degree of knowledge and documentation of the traditional methods of cultivating the landrace |
| Bringing management technologies up to date | Degree of exploration of new farming technologies and mechanization tried and/or incorporated into cultivation of the landrace |
| Associations among farmers | Degree of association among farmers for the purpose of exploiting and commercializing the landrace |
| European Union geographical quality label | State of the process toward obtaining a European quality label |
| External quality control | Degree of external quality control of the product’s sensory traits |
| Farmers’ preferences | Degree of formulation of farmers’ preferences |
| Consumers’ preferences | Degree of formulation of consumers’ preferences |
| Use in restaurants | Degree of use of the product in restaurants (mainly in the area of cultivation) |
| Use in transformed products | Degree of transformation of the product (canning, etc.) |
| Collective promotion in events | Degree of promotion of the product at fairs and events |
| Accession | Virus 1 | Growth Habit 2 | Appearance 3 | Days to Flowering | Number of Pod/Plant | Yield (g/plant) | 100 Seeds Weight (g) |
|---|---|---|---|---|---|---|---|
| CF7102 | 2.11 ± 0.25 | 1.51 ± 0.24 | 1.22 ± 0.09 | 50.56 ± 1.01 | 38.56 ± 4.85 | 51.78 ± 8.64 | 33.46 ± 1.45 |
| CF114 | 2.00 ± 0.18 | 1.50 ± 0.22 | 1.52 ± 0.18 | 48.88 ± 0.01 | 33.01 ± 4.82 | 40.13 ± 0.67 | 34.12 ± 1.70 |
| CF201 | 2.50 ± 0.15 | 1.57 ± 0.17 | 1.21 ± 0.11 | 48.12 ± 0.31 | 30.14 ± 3.88 | 37.29 ± 7.72 | 32.81 ± 2.77 |
| CF214 | 2.14 ± 0.26 | 1.64 ± 0.24 | 1.57 ± 0.17 | 48.88 ± 0.02 | 34.14 ± 5.51 | 52.12 ± 6.59 | 40.33 ± 2.58 |
| CF301 | 2.32 ± 0.17 | 1.45 ± 0.14 | 1.45 ± 0.08 | 50.36 ± 0.76 | 46.09 ± 4.84 | 51.18 ± 5.68 | 31.48 ± 1.92 |
| CF314 | 2.42 ± 0.37 | 1.51 ± 0.18 | 1.33 ± 0.21 | 49.63 ± 0.84 | 47.00 ± 4.90 | 54.33 ± 8.01 | 28.50 ± 1.59 |
| CF401 | 1.63 ± 0.13 | 1.17 ± 0.08 | 1.22 ± 0.09 | 49.38 ± 0.56 | 43.78 ± 3.64 | 55.67 ± 5.34 | 34.89 ± 0.92 |
| CF106 | 1.83 ± 0.17 | 1.33 ± 0.17 | 1.17 ± 0.08 | 49.87 ± 0.74 | 43.56 ± 6.05 | 63.89 ± 7.56 | 36.48 ± 1.22 |
| CF501 | 2.38 ± 0.18 | 1.63 ± 0.23 | 1.31 ± 0.09 | 49.44 ± 0.63 | 37.38 ± 5.79 | 47.01 ± 9.61 | 33.33 ± 1.87 |
| CF514 | 2.08 ± 0.20 | 1.53 ± 0.32 | 1.33 ± 0.17 | 49.63 ± 0.84 | 43.33 ± 4.88 | 61.17 ± 7.84 | 39.17 ± 2.26 |
| CF614 | 2.29 ± 0.15 | 1.14 ± 0.14 | 1.07 ± 0.07 | 51.11 ± 1.04 | 47.57 ± 5.45 | 64.14 ± 6.87 | 35.20 ± 1.58 |
| CF701 | 2.23 ± 0.18 | 1.41 ± 0.14 | 1.27 ± 0.08 | 50.10 ± 0.68 | 43.42 ± 3.49 | 54.75 ± 6.12 | 36.36 ± 3.36 |
| CF714 | 1.86 ± 0.14 | 1.07 ± 0.07 | 1.43 ± 0.17 | 50.16 ± 0.92 | 53.29 ± 8.31 | 65.14 ± 9.75 | 37.86 ± 3.78 |
| CF7002 | 2.51 ± 0.13 | 1.35 ± 0.15 | 1.15 ± 0.08 | 51.11 ± 0.84 | 45.00 ± 5.32 | 51.91 ± 5.48 | 31.73 ± 2.12 |
| CF109 | 2.31 ± 0.23 | 1.31 ± 0.13 | 1.25 ± 0.13 | 50.01 ± 0.82 | 32.63 ± 2.71 | 39.88 ± 5.42 | 32.91 ± 2.43 |
| Initial population | 2.67 ± 0.02 | 0.94 ± 0.01 | 1.13 ± 0.01 | 56.44 ± 0.07 | 29.20 ± 0.28 | 40.73 ± 0.52 | 29.61 ± 0.18 |
| Selected plants | 2.17 ± 0.06 | 1.40 ± 0.04 | 1.29 ± 0.03 | 49.81 ± 0.22 | 41.25 ± 1.56 | 52.67 ± 2.09 | 34.56 ± 0.73 |
| Locality | Odor 1 | Flavor | Seed-Coat Perception | Mealiness | Seed-Coat Roughness | Brightness | % Whole Seeds after Cooking |
|---|---|---|---|---|---|---|---|
| Loc 1 | 3.2 a | 35 b | 0.8 b | 4.3 a | 4.3 a | 6.2 b | 72.1 a |
| Loc 2 | 2.8 a | 3.0 b | 1.9 a | 4.5 a | 4.2a | 7.2 a | 64.5 b |
| Loc 3 | 3.2 a | 4.6 a | 1.1 b | 4.3 a | 4.7 a | 6.4 b | 65.8 b |
| Accession | Odor 1 | Flavor | Seed-Coat Perception | Mealiness | Seed-Coat Roughness | Brightness | % Whole Seeds after Cooking |
|---|---|---|---|---|---|---|---|
| CF109 | 3.3 a | 3.7 a | 0.8 a | 3.5 a | 5.2 a | 5.4 a | 74.8 a |
| CF106 | 2.5 b | 3.5 a | 1.6 b | 4.7 b | 6.0 b | 6.7 b | 62.0 a |
| Inbred Line | Yield (g/plant) | 100-Seed Weight (g) | Projected Area of the Seed (mm2) | Seed Width/Length |
|---|---|---|---|---|
| LCF101 | 37.7 cd | 31.5 ab | 66.4 bcd | 0.67 b |
| LCF122 | 27.8 e | 34.5 a | 70.4 bcd | 0.68 ab |
| LCF181 | 28.4 e | 35.9 a | 80.7 a | 0.63 c |
| LCF191 | 43.3 bc | 26.5 b | 58.1 d | 0.67 b |
| LCF420 | 52.6 a | 37.7 a | 73 ab | 0.68 ab |
| LCF810 | 33.4 de | 27.2 b | 61.7 cd | 0.68 ab |
| LCF109 | 47.0 ab | 25.4 b | 56.2 d | 0.7 a |
| Inbred Line | Odor 1 | Flavor | Seed-Coat Perception | Mealiness | Seed-Coat Roughness | Brightness | % Whole Seeds after Cooking |
|---|---|---|---|---|---|---|---|
| LCF101 | 4.9 a | 5.7 abc | 3.0 ab | 4.7 a | 4.9 b | 6.5 a | 77.5 a |
| LCF122 | 5.1 a | 6.5 a | 1.3 cd | 4.5 a | 5.7 ab | 6.0 ab | 72.5 a |
| LCF181 | 5.5 a | 6.3 ab | 2.1 bc | 4.8 a | 6.2 a | 5.9 ab | 65.7 ab |
| LCF191 | 4.7 a | 5.2 bc | 2.2 bc | 3.9 a | 5.2 ab | 6.9 a | 76.8 a |
| LCF420 | 4.7 a | 4.8 c | 3.9 a | 4.3 a | 5.1 ab | 6.7 a | 46.6 b |
| LCF810 | 4.6 a | 6.3 ab | 1.7 cd | 4.0 a | 6.0 ab | 5.9 ab | 68.2 ab |
| LCF109 | 4.5 a | 6.2 ab | 0.8 d | 3.8 a | 5.8 ab | 5.2 b | 78.6 a |
| Cotyledon | Seed Coat | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Inbred Line | Starch | Protein | Amylose | Apparent Amylose | Ash | Ca | Mg | Dietary Fiber | P | Uronic Acid | |
| LCF101 | 383.70 a | 257.79 a | 273.90 a | 328.21 a | 68.31 ab | 18.33 b | 3.40 ab | 769.45 a | 0.23 ab | 128.49 a | |
| LCF122 | 397.89 a | 248.34 a | 301.91 a | 328.84 a | 74.17 ab | 20.83 ab | 3.29 abc | 789.85 a | 0.18 ab | 125.27 a | |
| LCF181 | 362.76 a | 284.96 a | 273.58 a | 312.09 a | 65.66 b | 17.01 b | 3.18 bc | 808.36 a | 0.11 b | 130.00 a | |
| LCF191 | 366.33 a | 269.90 a | 312.35 a | 333.27 a | 60.85 b | 14.56 b | 3.47 ab | 817.60 a | 0.30 a | 125.79 a | |
| LCF420 | 386.60 a | 260.40 a | 289.85 a | 328.22 a | 76.58 ab | 20.86 ab | 3.32 abc | 822.11 a | 0.23 ab | 123.45 a | |
| LCF810 | 388.20 a | 251.48 a | 282.00 a | 330.35 a | 71.68 ab | 19.28 ab | 3.72 a | 804.95 a | 0.21 ab | 121.54 a | |
| LCF109 | 361.94 a | 277.62 a | 265.63 a | 319.80 a | 88.26 a | 25.66 a | 2.95 c | 772.91 a | 0.18 ab | 122.26 a | |
| Inbred | Days to Flowering | General Value 1 | Number of Pods/Plant | Pod Length (cm) | Yield (g/Plant) | 100 Seeds Weight (g) | Projected Area of the Seed (mm2) | Seed Width/length |
|---|---|---|---|---|---|---|---|---|
| LCF6001 | 49 b | 2.9 a | 62 bc | 10.5 ab | 56.4 b | 23.4 c | 46.2 b | 0.67 bc |
| LCF6012 | 57 c | 1.8 b | 60 c | 10.9 a | 65.1 ab | 27.4 ab | 48.6 b | 0.67 b |
| LCF6071 | 43 a | 2.4 ab | 67 b | 10.8 a | 67.9 ab | 24.2 bc | 47.2 b | 0.66 bc |
| LCF6091 | 43 a | 2 ab | 63 bc | 10.9 a | 63.1 b | 25.5 abc | 50.5 b | 0.66 bc |
| LCF109 | 46 ab | 2.1 ab | 61 c | 10.3 ab | 74.9 a | 28.9 a | 58.9 a | 0.65 c |
| Croscat | 54 c | 1.9 ab | 76 a | 9.7 b | 71.3 ab | 26.5 abc | 45.9 b | 0.73 a |
| Inbred | Odor 1 | Flavor | Seed-Coat Perception | Mealiness | Seed-Coat Roughness | Brightness | % Whole Seeds after Cooking |
|---|---|---|---|---|---|---|---|
| LCF6001 | 4.4 a | 5.6 a | 1.2 ab | 2.3 b | 7.1 a | 7.3 a | 50 b |
| LCF6012 | 4.7 a | 4.4 b | 2.0 a | 2.9 ab | 6.7 b | 6.6 ab | 62 b |
| LCF6071 | 4.5 a | 5.6 a | 1.2 ab | 2.4 b | 7.2 a | 7.3 a | 58 b |
| LCF6091 | 4.6 a | 5.5 a | 1.3 ab | 2.3 b | 7.3 a | 7.2 a | 60 b |
| LCF109 | 4.4 a | 5.1 ab | 0.9 b | 3.3 a | 4.7 c | 4.7 c | 91 a |
| Croscat | 4.1 b | 6.2 c | 2.1 a | 3.8 a | 4.6 c | 3.1 d | 18 c |
| Landrace and species | Number In Situ Accessions T0 | Number In Situ Accessions T1 | Number Ex Situ Accessions T0 | Number Ex Situ Accessions T1 | Number Selected Varieties T0 | Number Selected Varieties T1 | Number Varieties Derived T0 | Number Varieties Derived T1 | CPI T0 | CPI T1 |
|---|---|---|---|---|---|---|---|---|---|---|
| Castellfollit del Boix bean (Phaseolus vulgaris L.) | 15 | 4 | 15 | 15 | 0 | 4 | 0 | 4 | 0.21 ± 0.05 | 0.65 ± 0.03 |
| Ganxet bean (Phaseolus vulgaris L.) | 257 | >140 | 3 | 257 | 0 | 28 | 0 | 0 | 0.23 ± 0.04 | 0.80 ± 0.01 |
| Tavella Brisa bean (Phaseolus vulgaris L.) | 17 | <10 | 17 | 17 | 0 | 0 | 0 | 4 | 0.30 ± 0.02 | 0.80 ± 0.06 |
| Blanca Tardana de Lleida onion (Allium cepa L.) | 16 | 10 | 1 | 14 | 0 | 2 | 0 | 3 | 0.36 ± 0.01 | 0.75 ± 0.02 |
| Coll de Nargó onion (Allium cepa L.) | 4 | 4 | 0 | 4 | 0 | 1 | 0 | 0 | 0.10 ± 0.02 | 0.29 ± 0.03 |
| Espigall del Garraf (Brassica oleracea L.) | 7 | 4 | 4 | 5 | 0 | 1 | 0 | 0 | 0.20 ± 0.06 | 0.52 ± 0.00 |
| “D’escairar” maize (Zea mays L.) | 6 | 6 | 6 | 6 | 0 | 0 | 0 | 0 | 0.15 ± 0.03 | 0.15 ± 0.03 |
| Mandó tomato (Solanum lycopersicum L.) | 1 | 1 | 0 | 1 | 0 | 6 | 0 | 0 | 0.07 | 0.64 |
| Penjar tomato (Catalonia) (Solanum lycopersicum L.) | 350 | 50 | 60 | 298 | 3 | 6 | 0 | 8 | 0.29 | 0.54 |
| Penjar tomato (Valencia) (Solanum lycopersicum L.) | 50 | 53 | 69 | 72 | 3 | 3 | 3 | 3 | 0.4 | 0.66 |
| Pera Girona tomato (Solanum lycopersicum L.) | 60 | 10 | 25 | 62 | 0 | 1 | 0 | 2 | 0.14 | 0.42 |
| Valencià tomato (Solanum lycopersicum L.) | >100 | >100 | 131 | 146 | 0 | 7 | 0 | 7 | 0.28 | 0.56 |
| Almagro eggplant (Solanum melongena L.) | 25 | 20 | 4 | 18 | 0 | 1 | 0 | 0 | 0.36 | 0.68 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Casals, J.; Rivera, A.; Rull, A.; Romero del Castillo, R.; Sabaté, J.; Sans, S.; Soler, S.; Díez, M.J.; Casañas, F.; Prohens, J.; et al. Improving the Conservation and Use of Traditional Germplasm through Breeding for Local Adaptation: The Case of the Castellfollit del Boix Common Bean (Phaseolus vulgaris L.) Landrace. Agronomy 2019, 9, 889. https://doi.org/10.3390/agronomy9120889
Casals J, Rivera A, Rull A, Romero del Castillo R, Sabaté J, Sans S, Soler S, Díez MJ, Casañas F, Prohens J, et al. Improving the Conservation and Use of Traditional Germplasm through Breeding for Local Adaptation: The Case of the Castellfollit del Boix Common Bean (Phaseolus vulgaris L.) Landrace. Agronomy. 2019; 9(12):889. https://doi.org/10.3390/agronomy9120889
Chicago/Turabian StyleCasals, Joan, Ana Rivera, Aurora Rull, Roser Romero del Castillo, Josep Sabaté, Silvia Sans, Salvador Soler, María José Díez, Francesc Casañas, Jaime Prohens, and et al. 2019. "Improving the Conservation and Use of Traditional Germplasm through Breeding for Local Adaptation: The Case of the Castellfollit del Boix Common Bean (Phaseolus vulgaris L.) Landrace" Agronomy 9, no. 12: 889. https://doi.org/10.3390/agronomy9120889
APA StyleCasals, J., Rivera, A., Rull, A., Romero del Castillo, R., Sabaté, J., Sans, S., Soler, S., Díez, M. J., Casañas, F., Prohens, J., & Simó, J. (2019). Improving the Conservation and Use of Traditional Germplasm through Breeding for Local Adaptation: The Case of the Castellfollit del Boix Common Bean (Phaseolus vulgaris L.) Landrace. Agronomy, 9(12), 889. https://doi.org/10.3390/agronomy9120889