Biocontrol of Postharvest Fruit Fungal Diseases by Bacterial Antagonists: A Review

 , and
, and
Abstract
:1. Introduction
2. Biocontrol as a Method for Decreasing Phytopathogen Abundance
2.1. Selection and Identification of a Biological Control Agent
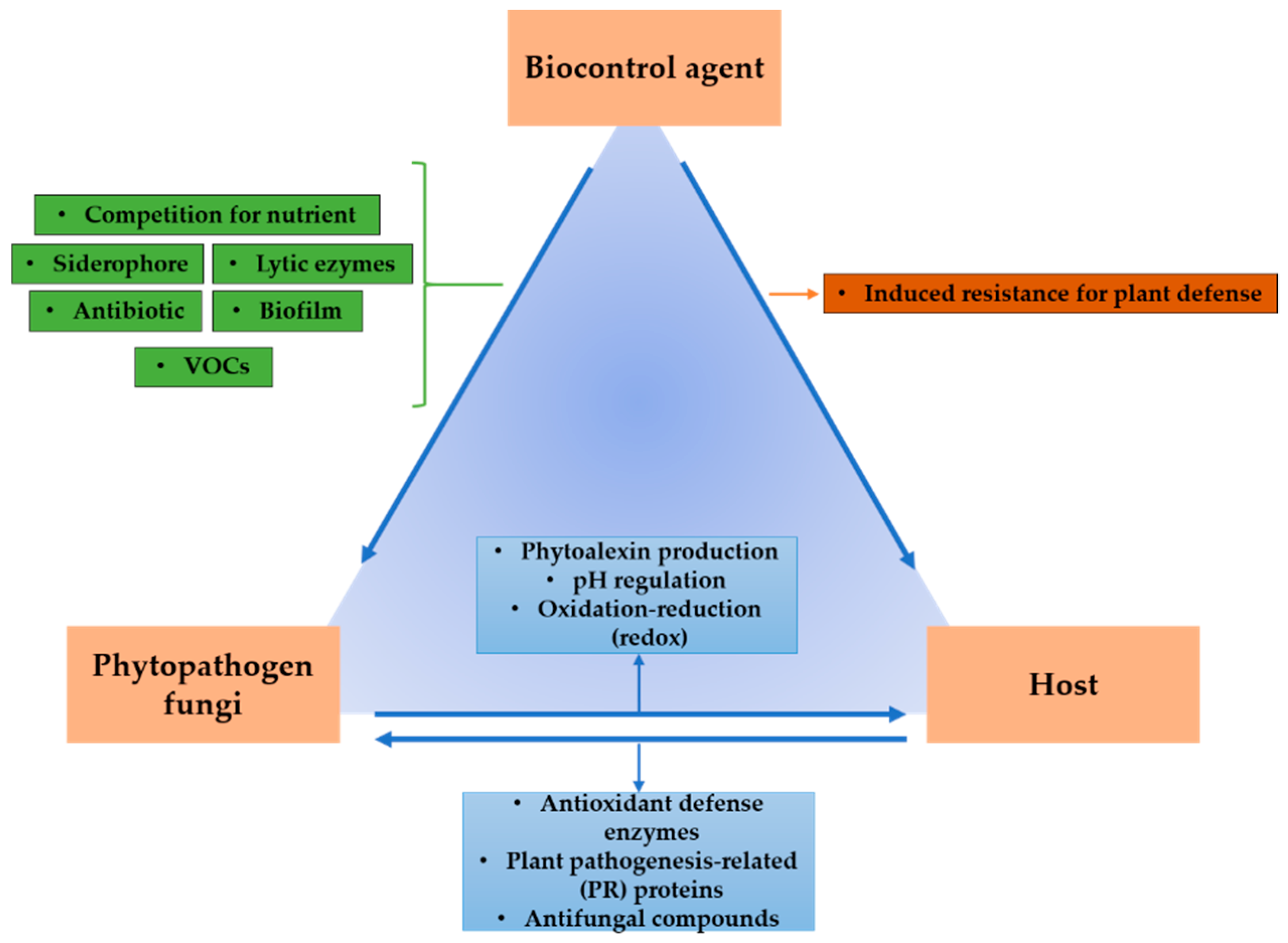
2.2. Antagonistic Mechanisms of Bacteria Utilized as Biocontrol Agents
2.2.1. Competition for Space and Nutrients
2.2.2. Siderophores
2.2.3. Parasitism (Lithic Enzyme Production)
2.2.4. Formation of Biofilms and Quorum Sensing
2.2.5. Antibiosis
2.2.6. Induction of Resistance
2.3. Commercializing Biological Products
2.4. Search for New Antagonists
2.5. Application of Biocontrol Agents in Preharvest Stages
2.6. Application of Biocontrol Agents Postharvest
3. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Zhang, H.; Mahunu, G.K.; Castoria, R.; Apaliya, M.T.; Yang, Q. Argumentation of biocontrol agents with physical methods against postharvest diseases of fruits and vegetables. Trends Food Sci. Technol. 2017, 69, 36–45. [Google Scholar] [CrossRef]
- Nunes, C.A. Biological control of postharvest diseases of fruit. Eur. J. Plant Pathol. 2012, 133, 181–196. [Google Scholar] [CrossRef]
- Droby, S.; Wisniewski, M.; Macarisin, D.; Wilson, C. Twenty years of postharvest biocontrol Research: Is it time for a new paradigm? Postharvest Biol. Technol. 2009, 52, 137–145. [Google Scholar] [CrossRef]
- Cuthbert, R.N.; Dick, J.T.; Callaghan, A.; Dickey, J.W. Biological control agent selection under environmental change using functional responses, abundances and fecundities; the relative control potential (RCP) metric. Biol. Control 2018, 121, 50–57. [Google Scholar] [CrossRef]
- Safdarpour, F.; Khodakaramian, G. Assessment of antagonistic and plant growth promoting activities of tomato endophytic bacteria in challenging with Verticillium dahliae under in-vitro and in-vivo conditions. Biol. J. Microorg. 2019, 7, 77–90. [Google Scholar]
- Dukare, A.S.; Sangeeta, P.; Eyarki, N.V.; Ram, K.G.; Rajbir, S.; Kalyani, S.; Rajesh, K.V. Exploitation of microbial antagonists for the control of postharvest diseases of fruits: A review. Crit. Rev. Food Sci. Nutr. 2018. [Google Scholar] [CrossRef] [PubMed]
- Beneduzi, A.; Ambrosini, A.; Passaglia, L.M. Plant growth-promoting rhizobacteria (PGPR): their potential as antagonists and biocontrol agents. Genet. Mol. Biol. 2012, 35, 1044–1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghazanfar, M.U.; Hussan, M.; Hamid, M.I.; Ansari, S.U. Utilization of biological control agents for the management of postharvest pathogens of tomato. Pak. J. Bot. 2016, 48, 2093–2100. [Google Scholar]
- Torres, M.J.; Brandan, C.P.; Petroselli, G.; Erra-Balsells, R.; Audisio, M.C. Antagonistic effects of Bacillus subtilis subsp., subtilis and B. amyloliquefaciens against Macrophomina phaseolina: SEM study of fungal changes and UV-MALDI-TOF MS analysis of their bioactive compounds. Microbiol. Res. 2016, 182, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Mcdonald, B.A. Using dynamic diversity to achieve durable diseases resistance in agricultural ecosystems. Trop. Plant Pathol. 2014, 39, 191–196. [Google Scholar] [CrossRef]
- Marin, A.; Atarés, L.; Chialt, A. Improving function of biocontrol agents incorporated in antifungal fruit coatings: A review. Biocontrol Sci. Technol. 2017, 27, 1220–1241. [Google Scholar] [CrossRef]
- Gotor-Vila, A.; Teixidó, N.; Casals, C.; Torres, R.; De Cal, A.; Guijarro, B.; Usall, J. Biological control of brown rot in Stone fruit using Bacillus amyloliquefaciens CPA-8 under field conditions. Crop Prot. 2017, 102, 72–80. [Google Scholar] [CrossRef]
- Nikolić, I.; Berić, T.; Dimkić, I.; Popović, T.; Lozo, J.; Fira, D.; Stanković, S. Biological control of Pseudomonas syringae pv. aptata on sugar beet with Bacillus pumilus SS-10.7 and Bacillus amyloliquefaciens (SS-12.6 and SS-38.4) strains. J. Appl. Microbiol. 2019, 126, 165–176. [Google Scholar]
- Paz, I.C.P.; Santin, R.D.C.M.; Guimarães, A.M.; da Rosa, O.P.P.; Quecine, M.C.; e Silva, M.D.C.P.; Azevedo, J.L.; Matsumura, A.T.S. Biocontrol of Botrytis cinerea and Calonectria gracilis by eucalypts growth promoters Bacillus spp. Microb. Pathog. 2018, 121, 106–109. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Sui, Y.; Wisniewski, M.; Droby, S.; Liu, Y. Utilization of antagonistic yeast to manage postharvest fungal diseases of fruit. Int. J. Food Microbiol. 2013, 167, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, Y.; Fu, X.; Li, Y.; Wang, Q. Isolation and characterization of Bacillus amyloliquefaciens PG12 for the biological control of apple ring rot. Postharvest Biol. Technol. 2016, 115, 113–121. [Google Scholar] [CrossRef]
- Aiello, D.; Restuccia, C.; Stefani, E.; Vitale, A.; Cirvilleri, G. Postharvest biocontrol ability of Pseudomonas synxantha against Monilinia fructicola and Monilinia fructigena on stone fruit. Postharvest Biol. Technol. 2019, 149, 83–89. [Google Scholar] [CrossRef]
- Wallace, R.L.; Hirkala, D.L.; Nelson, L.M. Mechanisms of action of three isolates of Pseudomonas fluorescens active against postharvest grey mold decay of apple during commercial storage. Biol. Control 2018, 117, 13–20. [Google Scholar] [CrossRef]
- Kong, Q.; Shan, S.; Liu, Q.; Wang, Q.; Yu, F. Biocontrol of Aspergillus flavus on peanut kernels by use of a strain of Marine Bacillus megaterium. Int. J. Food Microbiol. 2010, 139, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Montiel, L.G.; Zulueta-Rodriguez, R.; Angulo, C.; Rueda-Puente, E.O.; Quiñonez-Aguilar, E.E.; Galicia, R. Marine yeasts and bacteria as biological control agents against anthracnose on mango. J. Phytopathol. 2017, 165, 833–840. [Google Scholar] [CrossRef]
- Fu, G.; Huang, S.; Ye, Y.; Wu, Y.; Cen, Z.; Lin, S. Characterization of a bacterial biocontrol strain B106 and its efficacy in controlling banana leaf spot and post-harvest anthracnose diseases. Biol. Control 2010, 55, 1–10. [Google Scholar] [CrossRef]
- Khleekorn, S.; Wongrueng, S. Evaluation of antagonistic bacteria inhibitory to Colletotrichum musae on banana. J. Agric. Technol. 2014, 10, 383–390. [Google Scholar]
- Calvo, H.; Marco, P.; Blanco, D.; Oria, R.; Venturini, M.E. Potential of new strain Bacillus amyloliquefaciens BUZ-14 as a biocontrol agent of postharvest fruit diseases. Food Microbiol. 2017, 63, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Balaraju, K.; Jeon, Y. Effects of rhizobacteria Paenibacillus polymyxa APEC136 and Bacillus subtilis APEC170 on biocontrol of postharvest pathogens of apple fruits. J. Zhejiang Univ-Sci. B 2016, 17, 931–940. [Google Scholar] [CrossRef] [PubMed]
- Panebianco, S.; Vitale, A.; Polizzi, G.; Scala, F.; Cirvilleri, G. Enhanced control of postharvest citrus fruit decay by means of the combined use of compatible biocontrol agents. Biol. Control 2015, 84, 19–27. [Google Scholar] [CrossRef]
- Kasfi, K.; Taheri, P.; Jafarpour, B.; Tarighi, S. Identification of epiphytic yeast and bacteria with potential for biocontrol of gray mold diseases on table grapes caused by Botrytis cinerea. Span. J. Agric. Res. 2018, 16, 1–16. [Google Scholar] [CrossRef]
- Bull, C.T.; Stack, J.P.; Smilanick, J.L. Pseudomonas syringae Strains ESC-10 and ESC-11 survive in wounds on citrus and control green and blue molds of citrus. Biol. Control 1997, 8, 81–88. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, Z.; Zhu, P.; Liu, Y.; Zhang, Z.; Mastuda, Y.; Toyoda, H.; Xu, L. Postharvest biological control of melon pathogens using Bacillus subtilis EXWB1. J. Plant Pathol. 2010, 92, 645–652. [Google Scholar]
- Bahadou, S.A.; Ouijja, A.; Karfach, A.; Tahiri, A.; Lahlali, R. New potential bacterial antagonists for the biocontrol of fire blight disease (Erwinia amylovora) in Morocco. Microb. Pathog. 2018, 117, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Thokchom, E.; Thakuria, D.; Kalita, M.C.; Sharma, C.K.; Talukdar, N.C. Root colonization by host-specific rhizobacteria alters indigenous root endophyte and rhizosphere soil bacterial communities and promotes the growth of mandarin orange. Eur. J. Soil Biol. 2017, 79, 48–56. [Google Scholar] [CrossRef]
- Di Francesco, A.; Mari, M. Biological control of postharvest diseases by microbial antagonists: How many mechanisms of action? Eur. J. Plant Pathol. 2016, 145, 711–717. [Google Scholar] [CrossRef]
- Hernandez-Montiel, L.G.; Gutierrez-Perez, E.D.; Murillo-Amador, B.; Vero, S.; Chiquito-Contreras, R.G.; Rincon-Enriquez, G. Mechanisms employed by Debaryomyces hansenii in biological control of anthracnose disease on papaya fruit. Postharvest Biol. Technol. 2018, 139, 31–37. [Google Scholar] [CrossRef]
- Adrees, H.; Haider, M.S.; Anjum, T.; Akram, W. Inducing systemic resistance in cotton plants against charcoal root rot pathogen using indigenous rhizospheric bacterial strains and chemical elicitors. Crop Prot. 2019, 115, 75–83. [Google Scholar] [CrossRef]
- Asari, S.; Matzén, S.; Petersen, M.A.; Bejai, S.; Meijer, J. Multiple effects of Bacillus amyloliquefaciens volatile compounds: Plant growth promotion and growth inhibition of phytopathogens. FEMS Microbiol. Ecol. 2016, 92, fiw070. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.M.; Lee, Y.H. Genes involved in nutrient competition by Pseudomonas putida JBC17 to suppress green mold in postharvest satsuma mandarin. J. Basic Microbiol. 2015, 55, 898–906. [Google Scholar] [CrossRef] [PubMed]
- Terpilowska, S.; Siwicki, A.K. Cell cycle and transmembrane mitochondrial potential analysis after treatment with chromium (iii), iron (iii), molybdenum (iii) or nickel (ii) and their mixtures. Toxicol. Res. 2019. [Google Scholar] [CrossRef]
- Golonka, R.; San Yeoh, B.; Vijay-Kumar, M. The iron tug-of-war between bacterial siderophores and innate immunity. J. Innate Immun. 2019. [Google Scholar] [CrossRef] [PubMed]
- Carroll, C.S.; Moore, M.M. Ironing out siderophore biosynthesis: A review of non-ribosomal peptide synthetase (NRPS)-independent siderophore synthetases. Crit. Rev. Biochem. Mol. Biol. 2018, 53, 356–381. [Google Scholar] [CrossRef] [PubMed]
- Cordova-Albores, L.C.; Zapotitla, E.S.; Ríos, M.Y.; Barrera-Necha, L.L.; Hernández-López, M.; Bautista-Banos, S. Microscopic study of the morphology and metabolic activity of Fusarium oxysporum f. sp. gladioli treated with Jatropha curcas oil and derivatives. J. Microsc. Ultrastruct. 2016, 4, 28–35. [Google Scholar] [PubMed]
- Zeng, J.; Xu, T.; Cao, L.; Tong, C.; Zhang, X.; Luo, D.; Han, S.; Pang, P.; Fu, W.; Yan, J.; et al. The role of iron competition in the antagonistic action of the rice endophyte Streptomyces sporocinereus OsiSh-2 against the pathogen Magnaporthe oryzae. Microb. Ecol. 2018, 76, 1021–1029. [Google Scholar] [CrossRef] [PubMed]
- Drehe, I.; Simonetti, E.; Ruiz, J.A. Contribution of the siderophores pyoverdine and enantio-pyochelin to fitness in soil of Pseudomonas protegens Pf-5. Curr. Microbiol. 2018, 75, 1560–1565. [Google Scholar] [CrossRef] [PubMed]
- Andreolli, M.; Zapparoli, G.; Angelini, E.; Lucchetta, G.; Lampis, S.; Vallini, G. Pseudomonas protegens MP12: A plant growth-promoting endophytic bacterium with broad-spectrum antifungal activity against grapevine phytopathogens. Microbiol. Res. 2019, 219, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Katiyar, D.; Hemantaranjan, A.; Dwivedi, P. Plant growth promoting rhizobacteria and their roles as fungal biocontrol agents: An overview. J. Plant Sci. Res. 2018, 34, 127–136. [Google Scholar] [CrossRef]
- De Los Santos-Villalobos, S.; Barrera-Galicia, G.C.; Miranda-Salcedo, M.A.; Peña-Cabriales, J.J. Burkholderia cepacia XXVI siderophore with biocontrol capacity against Colletotrichum gloeosporioides. World J. Microbiol. Biotechnol. 2013, 28, 2615–2623. [Google Scholar] [CrossRef] [PubMed]
- Kesaulya, H.; Hasinun, J.V.; Tuhumury, G.N.C. Potential of Bacillus spp. produces siderophores in suppressing the wilt diseases of banana plants. In Proceedings of the International Symposium on Food and Agro-Biodiversity, Semarang, Indonesia, 26–27 September 2017. [Google Scholar]
- Sasirekha, B.; Srividya, S. Siderophore production by Pseudomonas aeruginosa PF6, a biocontrol strain for Rhizoctonia solani and Colletotrichum gloeosporioides causing diseases in chilli. Agric. Nat. Resour. 2016, 50, 250–256. [Google Scholar]
- Yu, X.; Ai, C.; Xin, L.; Zhou, G. The siderophore-producing bacterium, Bacillus subtilis CAS15, has a biocontrol effect on Fusarium wilt and promotes the growth of pepper. Eur. J. Soil Biol. 2011, 47, 138–145. [Google Scholar] [CrossRef]
- Santoyo, G.; Valencia-Cantero, E.; Orozco-Mosqueda, M.C.; Peña-Cabriles, J.J.; Farias-Rodríguez, R. Role of siderophores in antagonic activity of Pseudomonas fluorescens ZUM80 against plant fungi. Terra Latinoam. 2010, 10, 53–60. [Google Scholar]
- Sanz, F.M.I.; Benuzzi, D.; Calvente, V.; Calvo, J.; Sansone, G.; Cerutti, S.; Raba, J. The use of siderophores for improving the control of postharvest in stored fruits and vegetables. In Microbial Pathogens and Strategies for Combating Them: Science, Technology and Education; Méndez-Vilas, A., Ed.; Formatex: Badajoz, Spain, 2013; pp. 1385–1394. [Google Scholar]
- Acebedo, G.Y.; Hernández-Rodríguez, A.; Rives, R.N.; Velázquez del Valle, M.G.; Hernández-Lauzardo, A.N. Perspectivas del uso de bacterias rizosféricas en el control de Pyricularia grisea (Cooke Sacc.) en el cultivo del arroz (Oryza sativa L). Rev. Colomb. Biotecnol. 2011, 13, 16–22. [Google Scholar]
- Spadaro, D.; Droby, S. Development of biocontrol products for postharvest diseases of fruits: The importance of elucidating the mechanisms of action of yeast antagonists. Trends Foods Sci. Technol. 2016, 47, 39–49. [Google Scholar] [CrossRef]
- Seidl, V. Chitinase of filamentous fungi a large group old the diverse proteins with multiple physiological functions. Fungal Biol. Rev. 2008, 22, 36–42. [Google Scholar] [CrossRef]
- Stoykov, Y.M.; Paulov, A.I.; Krastanov, A.I. Chitinase biotechnology: Production, purification and application. Eng. Life Sci. 2015, 15, 30–38. [Google Scholar] [CrossRef]
- Barret, A.J.; Rawlings, N.D.; Woesnner, J.F. The Handbook of Proteolytic Enzymes, 2nd ed.; Academic Press: London, UK, 2003. [Google Scholar]
- Yu, T.; Wang, L.; Yin, Y.; Wang, Y.; Zheng, X. Effect of chitin on the antagonistic activity of Cryotococcus laurentii against Penicillium expansum in pear fruit. Int. J. Food Microbiol. 2008, 122, 44–48. [Google Scholar] [CrossRef] [PubMed]
- Shivakumar, S.; Karmali, A.N.; Ruhimbana, C. Partial purification, characterization, and kinetic studies of a low-molecular-weight, alkali-tolerant chitinase enzyme Bacillus subtilis JN032305, a potential biocontrol strain. Prep. Biochem. Biotechnol. 2014, 44, 1–37. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, P.; Tozlu, E.; Kotan, R.; Kotan, M.S. Potential of some bacteria for biological control of postharvest citrus green mold caused by Penicillum digitatum. Plant Prot. Sci. 2017, 53, 1–10. [Google Scholar]
- Chi, M.; Li, G.; Liu, Y.; Liu, G.; Li, G.; Li, M.; Zhang, X.; Sun, Z.; Sui, Y.; Liu, J. Increase in antioxidant enzyme activity, stress tolerance and biocontrol efficacy of Pichia kudriavzevii with the transition from a yeast-like to biofilm morphology. Biol. Control 2015, 90, 113–119. [Google Scholar] [CrossRef]
- Che, Y.; Yan, F.; Chai, Y.; Lu, H.; Kolter, R.; Losick, R.; Guo, J.H. Biocontrol of tomato wilt disease by Bacillus subtilis isolates from natural environments depends on conserved genes mediating biofilm formation. Environ. Microbiol. 2013, 15, 848–864. [Google Scholar]
- Beauregard, P.B.; Chai, Y.; Vlamakis, H.; Losick, R.; Kolter, R. Bacillus subtilis biofilm induction by plant polysaccharides. Proc. Natl. Acad. Sci. USA 2013, 110, E1621–E1630. [Google Scholar] [CrossRef] [PubMed]
- Vlamakis, H.; Aguilar, C.; Losick, R.; Kolter, R. Control of cell fate the formation of an architecturally complex bacterial community. Genes Dev. 2008, 22, 945–953. [Google Scholar] [CrossRef] [PubMed]
- Ostrowski, A.; Meheter, A.; Prescitt, A.; Kiley, T.B.; Stanley-Wall, N.R. YuaB functions synergistically with the exopolysaccharide and TasA amyloid fibers to allow biofilm formation by Bacillus subtilis. J. Bacteriol. 2011, 193, 4821–4831. [Google Scholar] [CrossRef] [PubMed]
- Branda, S.S.; Chu, F.; Kearns, D.B.; Losick, R.; Kolter, R. A major protein component of the Bacillus subtilis biofilm matrix. Mol. Microbiol. 2006, 59, 1229–1238. [Google Scholar] [CrossRef] [PubMed]
- Romero, D.; Vlamakis, H.; Losick, R.; Kolter, R. An accessory protein required for anchoring and assembly of amyloid fibers in B. subtilis. Mol. Microbiol. 2011, 80, 1155–1168. [Google Scholar] [CrossRef] [PubMed]
- Branda, S.S.; Gonzales-Pastor, J.E.; Ben-Yehuda, S.; Losick, R.; Kolter, R. Fruiting body formation by Bacillus subtilis. Proc. Natl. Acad. Sci. USA 2001, 98, 11621–11626. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Iwano, M. BsIA (YuaB) forms a hydrophobic layer on the surface on the Bacillus subtilis biofilms. Mol. Microbiol. 2012, 85, 51–66. [Google Scholar] [CrossRef] [PubMed]
- Kaufrnan, G.; Liu, W.; Williams, D.M.; Choo, Y.; Gopinadhan, M.; Samudrala, N.; Sarfati, R.; Yan, E.C.Y.; Regan, L.; Osuji, C.O. Flat drops, elastic sheets, and microcapsules by interfacial assembly of a bacterial biofilm protein, BsIA. Langmuir 2017, 33, 13590–13597. [Google Scholar]
- Haggag, W.M.; Timmusk, S. Colonization of peanut roots by biofilm-forming Paenibacillus polymyxa initiates biocontrol against crown rot diseases. J. Appl. Microbiol. 2008, 104, 961–969. [Google Scholar] [CrossRef] [PubMed]
- Bianciotto, V.; Andreotti, S.; Balstrini, R.; Bonfante, P.; Perotto, S. Mucoide mutants of the biocontrol strain Pseudomonas fluorescens CHA0 show increased ability in biofilm formation on mycorrhizal and nomycorrhizal carrot roots. Mol. Plant Microbe Interact. 2001, 14, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Bais, H.P.; Fall, R.; Vivanco, J.M. Biocontrol of Bacillus subtilis against infection of Arabidopsis roots by Pseudomona syringae is facilitated by biofilm formation and surfaction production. Plant Physiol. 2004, 134, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Torres, R.; Solsana, C.; Viñas, I.; Usall, J.; Plaza, P.; Teixidó, N. Optimization of packaging and storage conditions of a freeze dried Pantoea agglomerans formulation for controlling postharvest diseases in fruit. J. Appl. Microbiol. 2014, 117, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Batista, F.M.; Duarte, M.M.H.; Tremocoldi, A.R.; Pascholati, S.F. Potential of antimicrobial volatile organic compounds to control Sclerotinis sclerotiorum in vean seeds. Pesq. Agropec. Bras. 2011, 4, 137–142. [Google Scholar]
- Guevara-Avendaño, E.; Bejarano-Bolívar, A.A.; Kiel-Martínez, A.L.; Ramírez-Vázquez, M.; Méndez-Bravo, A.; von Wobeser, E.A.; Sanchez-Rangel, D.; Guerrero-Analco, J.A.; Eskalen, A.; Reverchon, F. Avocado rhizobacteria emit volatile organic compounds with antifungal activity against Fusarium solani, Fusarium sp. associated with Kuroshio shot hole borer, and Colletotrichum gloeosporioides. Microbiol. Res. 2019, 219, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Layton, C.; Maldonado, E.; Monroy, L.; Corrales, L.C.; Sánchez, L.C. Bacillus spp.; perspectiva de su efecto biocontrolador mediante antibiosis en cultivos afectados por fitopatógenos. Cienc. Bioméd. 2011. [Google Scholar] [CrossRef]
- Velázquez-Becerra, C.; Macías-Rodríguez, L.I.; López-Bucio, J.; Flores-Cortez, I.; Santoyo, G.; Hernández-Soberano, C.; Valencia-Cantero, E. The rhizobacterium Arthrobacter agilis produces dimethylhexadecylamine, a compound that inhibits growth of phytopathogenic fungi in vitro. Protoplasma 2013, 250, 1251–1262. [Google Scholar] [CrossRef] [PubMed]
- Raza, W.; Ling, N.; Yang, L.; Huang, Q.; Shen, Q. Response of tomato wilt pathogen Ralstonia solanacearum to the volatile organic compounds produced by biocontrol strain Bacillus amyloliquefaciens SQR-9. Sci. Rep. 2015, 6, 24856. [Google Scholar] [CrossRef] [PubMed]
- Jamalizadeh, M.; Etebarian, H.R.; Aminian, H.; Alizadeh, A. A review of mechanisms of action of biological control organisms against post-harvest fruit spoilage. Bull. OEPP/EPPO 2011, 41, 65–71. [Google Scholar] [CrossRef]
- Hacquard, S.; Spaepen, S.; Garrido-Oter, R.; Schulze-Lefert, P. Interplay between innate immunity and the plant microbiota. Annu. Rev. Phytopathol. 2017, 55, 565–589. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Nolan, T.M.; Song, G.; Liu, S.; Xie, Z.; Chen, J.; Schnable, P.S.; Walley, J.W.; Yin, Y. FERONIA receptor kinase contributes to plant immunity by suppressing jasmonic acid signaling in Arabidopsis thaliana. Curr. Biol. 2018, 28, 3316–3324. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Xia, W.; Cao, P.; Xiao, Z.A.; Zhang, Y.; Liu, M.; Zhan, C.; Wang, N. Integrated transcriptome analysis reveals plant hormones jasmonic acid and salicylic acid coordinate growth and defense responses upon fungal infection in poplar. Biomolecules 2019, 9, E12. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, B.; Marques, A.; Ramos, C.; Neng, R.N.; Nogueira, M.F.J.; Saravia, A.J.; Nunes, L.M. Chemical compositions and antibacterial and antioxidant properties of commercial essential oils. Ind. Crop. Prod. 2013, 43, 587–595. [Google Scholar] [CrossRef]
- Holert, J.; Cardenas, E.; Bergstrand, L.H.; Zaikova, E.; Hahn, A.S.; Hallam, S.J.; Mohn, W.W. Metagenomes Reveal global distribution of bacterial steroid catabolism in natural, engineered, and host environments. mBio 2018, 9, e0234517. [Google Scholar] [CrossRef] [PubMed]
- Droby, S.; Wisniewski, M.; Teixidó, N.; Spadaro, D.; Jijakli, M.H. The science, development, and commercialization of postharvest biocontrol products. Postharvest Biol. Technol. 2016, 122, 22–29. [Google Scholar] [CrossRef] [Green Version]
- Blachinsky, D.; Antonov, J.; Bercovitz, A.; El-ad, B.; Feldman, K.; Husid, A.; Lazare, M.; Marcov, N.; Shamai, I.; Droby, S.; et al. Commercial applications of shemer for the control of pre- and post-harvest diseases. IOBC/WPRS Bull. 2007, 30, 75–78. [Google Scholar]
- Teixidó, N.; Torres, R.; Viñas, I.; Abadias, M.; Usall, J. Biological control of postharvest diseases in fruit and vegetables. In Protective Cultures, Antimicrobial Metabolites and Bacteriophages for Food and Beverage Biopreservation; Woodhead Publishing: Cambridge, UK, 2011; pp. 364–402. [Google Scholar]
- Janisiewicz, W.J.; Korsten, L. Biological control of postharvest diseases of fruits. Ann. Rev. Phytopathol. 2002, 40, 411–441. [Google Scholar] [CrossRef] [PubMed]
- Janisiewicz, W.J.; Peterson, D.J. Susceptibility of the stem pull area of mechanically harvest apples to blue mold decay and its control with a biocontrol agent. Plant Dis. 2004, 88, 662–664. [Google Scholar] [CrossRef] [PubMed]
- Demoz, B.T.; Korsten, L. Bacillus subtilis attachment, colonization, and survival on avocado flowers and its mode of action on stem-end rot pathogens. Biol. Control 2006, 37, 68–74. [Google Scholar] [CrossRef]
- Lahlali, R.; Raffaele, B.; Jijakli, M.H. UV protectants for Candida oleophila (strain O), a biocontrol agent of postharvest fruit diseases. Plant Pathol. 2011, 60, 288–295. [Google Scholar] [CrossRef]
- Massart, S.; Jijakli, M.H. Pichia anomala and Candida oleophila in biocontrol of postharvest diseases of fruits: 20 years of fundamental and practical Research. In Post-Harvest Pathology, Plant Pathology in the 21st Century; Prusky, D., Gullino, M.L., Eds.; Springer International Publishing: Basel, Switzerland, 2014; pp. 111–122. [Google Scholar]
- Lima, G.; Sanzani, S.M.; Curtis, F.; Ippolito, A. Biological control of postharvest diseases. In Advances in Postharvest Fruit and Vegetables Technology; Wills, R.B.H., Golding, J., Eds.; CRC Press: Boca Raton, FL, USA, 2015; pp. 65–81. [Google Scholar]
- Usall, T.; Torres, R.; Teixidó, N. Biological control of postharvest diseases on fruit a suitable alternative? Curr. Opin. Food Sci. 2016, 11, 51–55. [Google Scholar] [CrossRef]
- Maida, I.; Chiellini, C.; Mengoni, A.; Bosi, E.; Firenzuoli, F.; Fondi, M.; Fani, R. Antagonistic interactions between endophytic cultivable bacterial communities isolated from the medicinal plant Echinacea purpurea. Environ. Microbiol. 2016, 18, 2357–2365. [Google Scholar] [CrossRef] [PubMed]
- Goudjal, Y.; Toumatia, O.; Yekkour, A.; Sabaou, N.; Mathieu, F.; Zitouni, A. Biocontrol of Rhizoctonia solani damping-off and promoting of tomato plant growth by endophytic actinomycete isolated from native plants of Algerian Sahara. Microbiol. Res. 2014, 169, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Ab Rahman, S.F.S.; Singh, E.; Pieterse, C.M.; Schenk, P.M. Emerging microbial biocontrol strategies for plant pathogens. Plant Sci. 2018, 267, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Chenniappan, C.; Narayanasamy, M.; Daniel, G.M.; Ramaraj, G.B.; Ponnusamy, P.; Sekar, J.; Ramalingam, P.V. Biocontrol efficiency of native plant growth promoting rhizobacteria against rhizome rot disease of turmeric. Biol. Control 2019, 129, 55–64. [Google Scholar] [CrossRef]
- Larran, S.; Simón, M.R.; Moreno, M.V.; Santamarina, S.M.P.; Perelló, A. Endophytes from wheat as biocontrol agents against tan spot disease. Biol. Control 2016, 92, 17–23. [Google Scholar] [CrossRef]
- Comby, M.; Gacoin, M.; Robineau, M.; Rabenoelina, F.; Ptas, S.; Dupont, J.; Prozi, C.; Baillieul, F. Screening of wheat endophytes as biological control agents against Fusarium head blight using two different in vitro test. Microbiol. Res. 2017, 202, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Santoyo, G.; Moreno-Hagelsieb, G.; Orozco-Mosqueda, M.C.; Glick, B.R. Plant growth-promoting bacterial endophytes. Microbiol. Res. 2016, 183, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Chen, J.; Hu, G.; Yu, J.; Zhu, X.; Lin, Y.; Chen, S.; Yuan, J. Statistical Research on the bioactivity of new marine natural products discovered during the 28 years from 1985 to 2012. Mar. Drugs 2015, 13, 202–221. [Google Scholar] [CrossRef] [PubMed]
- Santos, S.M.C.; Polonio, J.C.; Quecine, M.C.; Almeida, T.T.; Bogas, A.C.; Pamphile, J.A.; Pereira, J.O.; Asrolfi-Fiho, S.; Azevedo, J.L. Endophytic cultivable bacteria community obtained from the Paullinia cupana seed in Amazonas and Bahia regions and its antagonistic effects against Colletotrichum gloeosporioides. Microb. Pathog. 2016, 98, 16–22. [Google Scholar]
- Sinimol, S.; Sarika, A.R.; Nair, A.J. Diversity and antagonistic potential of marine microbes collected from south-west coast of India. 3 Biotech. 2016, 6, 7. [Google Scholar] [CrossRef] [PubMed]
- Bibi, F.; Nasser, M.I.; Hassan, A.M.; Yasir, M.; Al-Ghamdi, A.A.K.; Azhar, E.I. Diversity and antagonistic potential of bacteria isolated from marine grass Halodule uninervis. 3 Biotech. 2018, 8, 48. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Montiel, L.G.; Larralde-Corona, C.P.; Vero, S.; Lopez-Aburto, M.G.; Ochoa, J.L.; Ascencio-Valle, F. Characterization of yeast Debaryomyces hansenii for biological control of blue mold decay of Mexican lemon. CyTA J. Food. 2010, 8, 49–56. [Google Scholar] [CrossRef]
- Aloui, H.; Liccardello, F.; Khawldia, K.; Hamdi, M.; Restuccia, C. Physical properties and antifungal activity of bioactive films containing Wickerhamomyces anomalus killer yeast and their application for preservation of oranges and control of postharvest green mold caused by Penicillium digitatum. Int. J. Food Microbiol. 2015, 200, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Rivas-Garcia, T.; Murillo-Amador, B.; Nieto-Garibay, A.; Chiquito-Contreras, R.; Rincon-Enriquez, G.; Hernandez-Montiel, L.G. Effect of ulvan on the biocontrol activity of Debaryomyces hansenii and Stenotrophomonas rhizophila against fruit rot of Cucumis melo L. Agronomy 2018, 8, 273. [Google Scholar] [CrossRef]
- Bibi, F.; Strobel, G.A.; Naseer, M.I.; Yasir, M.; Khalaf Al-Ghamdi, A.A.; Azhar, E.I. Halophytes-associated endophytic and rhizospheric bacteria: Diversity, antagonism and metabolite production. Biocontrol Sci. Technol. 2018, 28, 192–213. [Google Scholar] [CrossRef]
- Cong, Z.; Huang, X.; Liu, Y.; Liu, Y.; Wang, P.; Liao, S.; Yang, B.; Zhou, X.; Huang, D.; Wang, J. Cytotoxic anthracycline and antibacterial tirandamycin analogues from a marine-derived Streptomyces sp. SCSIO 41399. J. Antibiot. 2019, 72, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Gomes, E.D.B.; Dias, L.R.L.; Rita de Cassia, M. Actinomycetes bioactive compounds: Biological control of fungi and phytopathogenic insect. Afr. J. Biotechnol. 2018, 17, 552–559. [Google Scholar]
- Lopes, M.R.; Nadjara, K.M.; Pompeo, F.L.; Silva, A.C.; Kupper, K.C. Saccharomyces cerevisiae: A novel and efficient biological control agent for Colletotrichum acutatum during pre-harvest. Microbiol. Res. 2015, 175, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Silva, Y.Y.; De Costa, D.M. Potential of pre-harvest application of Burkholderia spinosa for biological control of epiphytic and pathogenic microorganisms on the phyllosphere of banana (Musa spp.). Trop. Agric. Res. 2014, 25, 443–454. [Google Scholar] [CrossRef]
- Camañas, T.P.; Viñas, I.; Usall, J.; Torres, R.; Anguera, M.; Teixidó, N. Control of postharvest diseases on citrus fruit by preharvest applications of biocontrol agent Pantoea agglomerans CPA-2 Part II. Effectiveness of different cell formulations. Postharvest Biol. Technol. 2008, 49, 96–106. [Google Scholar] [CrossRef]
- Qin, X.; Xiao, H.; Xue, C.; Yu, Z.; Yang, R.; Cai, Z.; Si, L. Biocontrol of gray mold in grapes with the yeast Hanseniaspora uvarum alone and in combination with salicylic acid or sodium bicarbonate. Postharvest Biol. Technol. 2015, 100, 160–167. [Google Scholar] [CrossRef]
- Han, J.; Chen, D.; Huang, J.; Li, X.; Zhou, W.W.; Gao, W.; Jia, Y. Antifungal activity and biocontrol potential of Paenibacillus polymyxa HT16 against white rot pathogen (Coniella diplodiella Speq.) in table grapes. Biocontrol Sci. Technol. 2015, 25, 1120–1132. [Google Scholar] [CrossRef]


{kind=link}
| Bacterial | Diseases | Phytopathogens | Hosts | Inhibition (%) | References |
|---|---|---|---|---|---|
| Bacillus amyloliquefaciens | Brown rot | Monilinia sp. | Apple | 75 | [16] |
| Pseudomonas synxantha | Brown rot | Monilinia fructicola | Peach | 70 | [17] |
| Pseudomonas fluorescens | Blue mold | Penicillium expansum | Apple | 88 | [18] |
| Bacillus megaterium | Damping-off | Aspergillus flavus | Peanut | 41 | [19] |
| Stenotrophomonas rhizophila | Anthracnose | Colletotrichum gloeosporioides | Mango | 89 | [20] |
| Bacillus subtilis | Anthracnose | Colletotrichum musae | Banana | 72 | [21] |
| Pantoea agglomerans | Anthracnose | Colletotrichum musae | Banana | 94 | [22] |
| Bacillus amyloliquefaciens | Blue mold | Penicillium expansum | Apple | 80 | [23] |
| Paenibacillus polymyxa | Anthracnose | Colletotrichum gloeosporioides | Apple | 83 | [24] |
| Pseudomonas syringae | Green mold | Penicillium digitatum | Citrus | 28 | [25] |
| Bacillus sp. | Gray mold | Botrytis cinerea | Grape | 50 | [26] |
| Pseudomonas syringae | Blue and green mold | Pseudomonas digitatum and Pseudomonas italicum | Citrus | 31 | [27] |
| Bacillus subtilis | Rot | Alternaria alternata | Melon | 77 | [28] |
| Genetically stable |
| Effective at low concentrations |
| Non-demanding in terms of required nutrients |
| Survival capability under adverse environmental conditions (including low/high temperature, storage with controlled atmospheres) |
| Capability of adhering to and colonizing the host surface |
| Showing more than one antagonistic mechanism against phytopathogens that need to be controlled |
| Effective against a range of phytopathogens |
| Production in a low-cost growth medium |
| Preparation in a simple formulation that can be stored for long time periods |
| Environmentally friendly |
| No production of secondary metabolites or harmful toxins |
| Bacteria | Disease | Siderophore Type | Phytopathogen | Host | Reference |
|---|---|---|---|---|---|
| Pseudomonas sp. | Damping-off | Pyoverdine | Pythium ultimum | Cucumber | [43] |
| Burkholderia cepacia | Anthracnose | Pyoverdine | Colletotrichum gloeosporioides | Mango | [44] |
| Bacillus sp. | Bacterial wilt | Bacilibactin | Ralstonia solanacearum | Banana | [45] |
| Pseudomonas aeruginosa | Damping-off | Pyoverdine | Rhizoctonia solani | Pepper | [46] |
| Bacillus subtilis | Anthracnose | Bacilibactin | Colletotrichum sp. | Pepper | [47] |
| Pseudomonas fluorescens | Anthracnose | Pyoverdine | Colletotrichum gloeosporioides | Avocado | [48] |
| Rahnella aquatilis | Molds | Enterochelin | Penicillium expansum | Fruit | [49] |
| Pseudomonas sp. | Piriculariosis | Pyoverdine | Pyricularia grisea | Rice | [50] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carmona-Hernandez, S.; Reyes-Pérez, J.J.; Chiquito-Contreras, R.G.; Rincon-Enriquez, G.; Cerdan-Cabrera, C.R.; Hernandez-Montiel, L.G. Biocontrol of Postharvest Fruit Fungal Diseases by Bacterial Antagonists: A Review. Agronomy 2019, 9, 121. https://doi.org/10.3390/agronomy9030121
Carmona-Hernandez S, Reyes-Pérez JJ, Chiquito-Contreras RG, Rincon-Enriquez G, Cerdan-Cabrera CR, Hernandez-Montiel LG. Biocontrol of Postharvest Fruit Fungal Diseases by Bacterial Antagonists: A Review. Agronomy. 2019; 9(3):121. https://doi.org/10.3390/agronomy9030121
Chicago/Turabian StyleCarmona-Hernandez, Saul, Juan J. Reyes-Pérez, Roberto G. Chiquito-Contreras, Gabriel Rincon-Enriquez, Carlos R. Cerdan-Cabrera, and Luis G. Hernandez-Montiel. 2019. "Biocontrol of Postharvest Fruit Fungal Diseases by Bacterial Antagonists: A Review" Agronomy 9, no. 3: 121. https://doi.org/10.3390/agronomy9030121
APA StyleCarmona-Hernandez, S., Reyes-Pérez, J. J., Chiquito-Contreras, R. G., Rincon-Enriquez, G., Cerdan-Cabrera, C. R., & Hernandez-Montiel, L. G. (2019). Biocontrol of Postharvest Fruit Fungal Diseases by Bacterial Antagonists: A Review. Agronomy, 9(3), 121. https://doi.org/10.3390/agronomy9030121