Development and Evaluation of a Leaf Disease Damage Extension in Cropsim-CERES Wheat

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
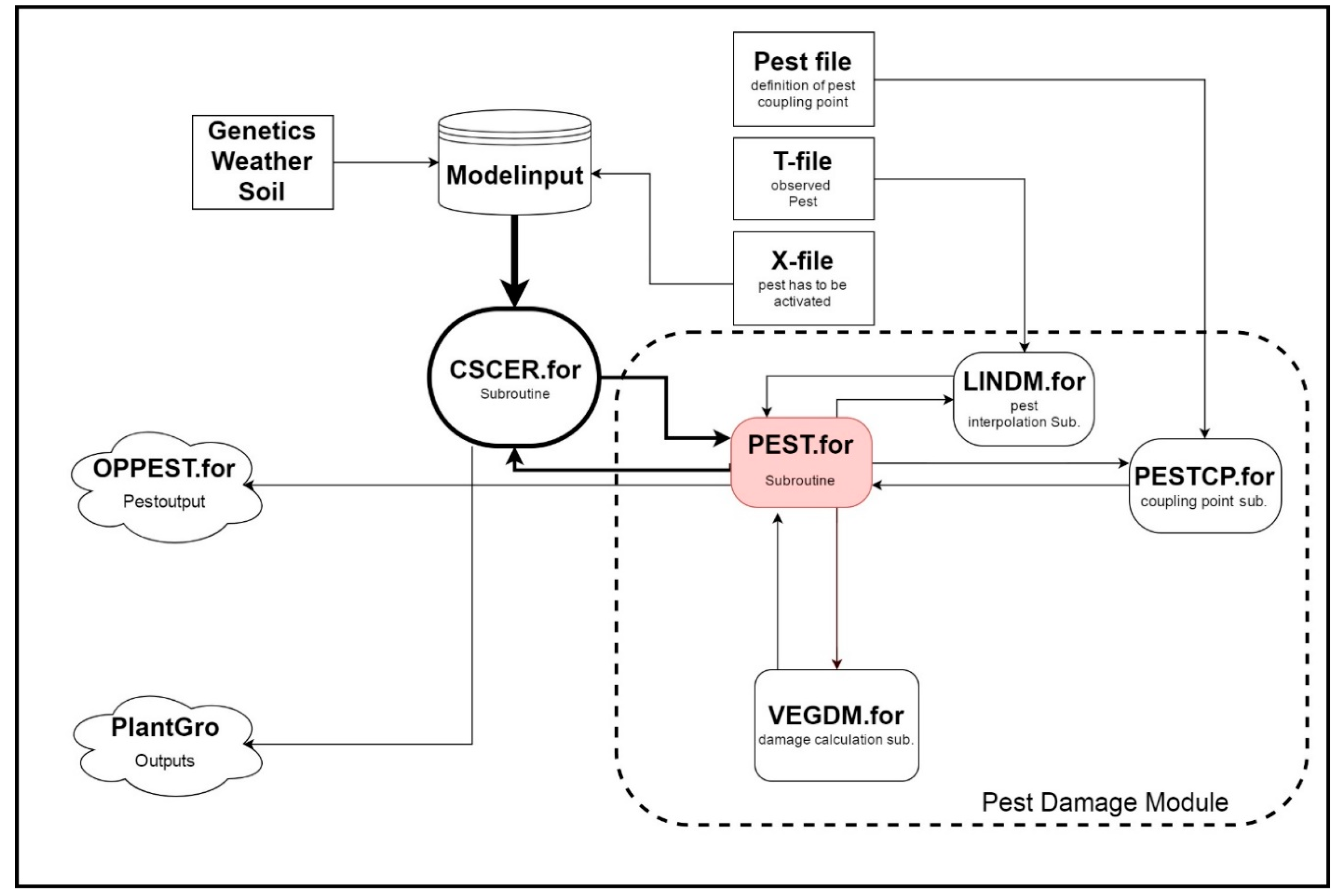
2.1. Model Development
2.2. Field Trials
2.3. Model Calibration and Evaluation
Statistical Evaluation
3. Results and Discussion
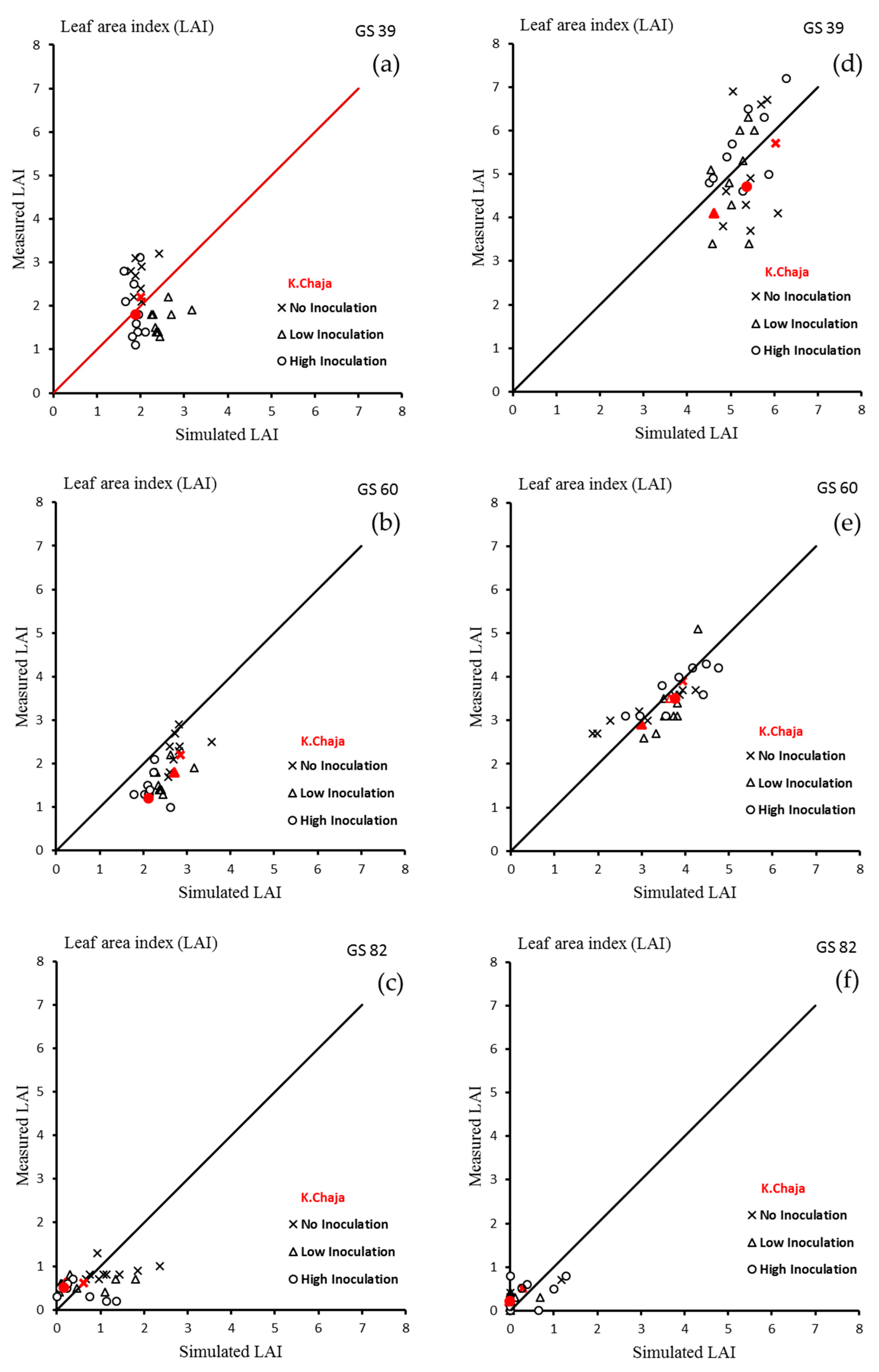
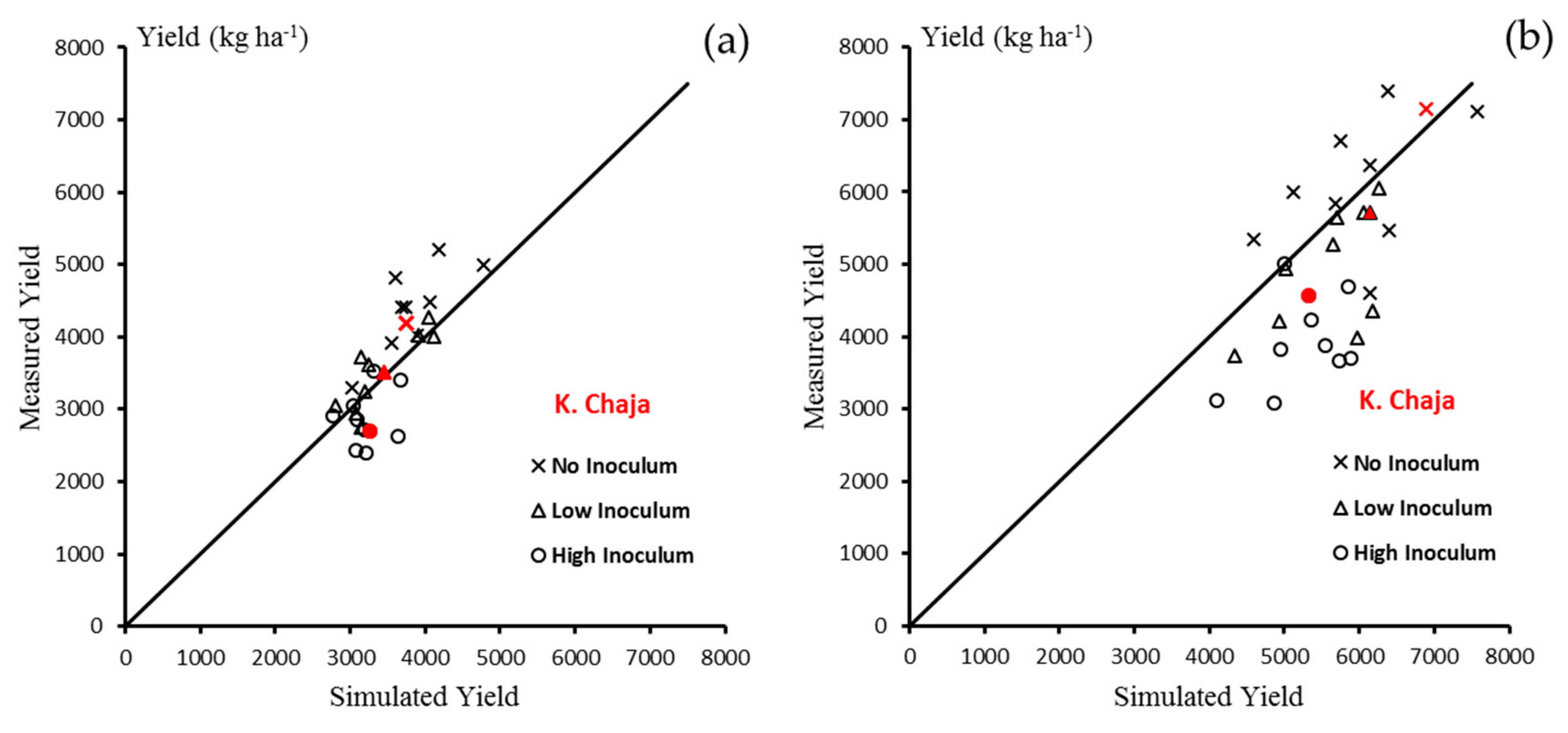
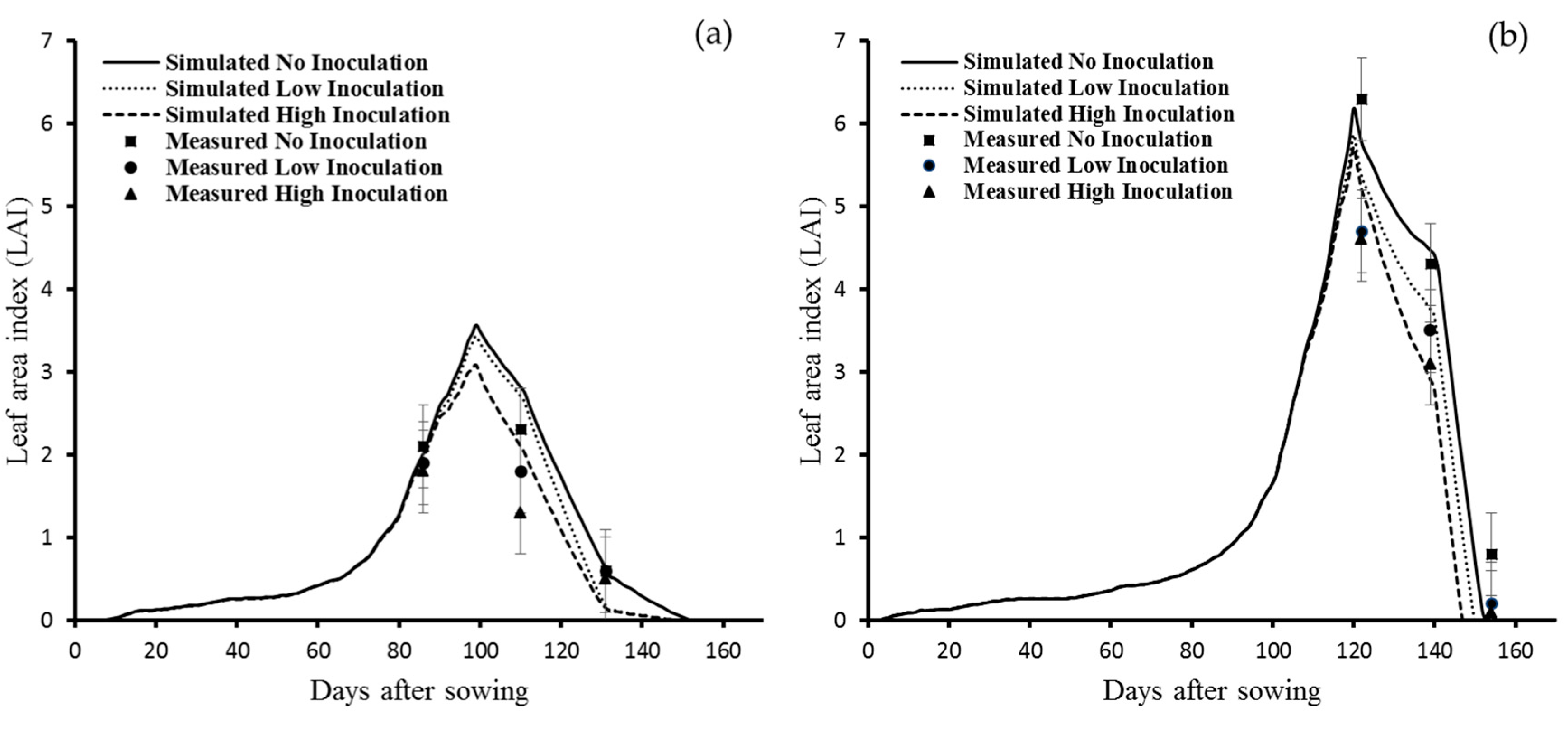
3.1. Model Calibration for La Plata
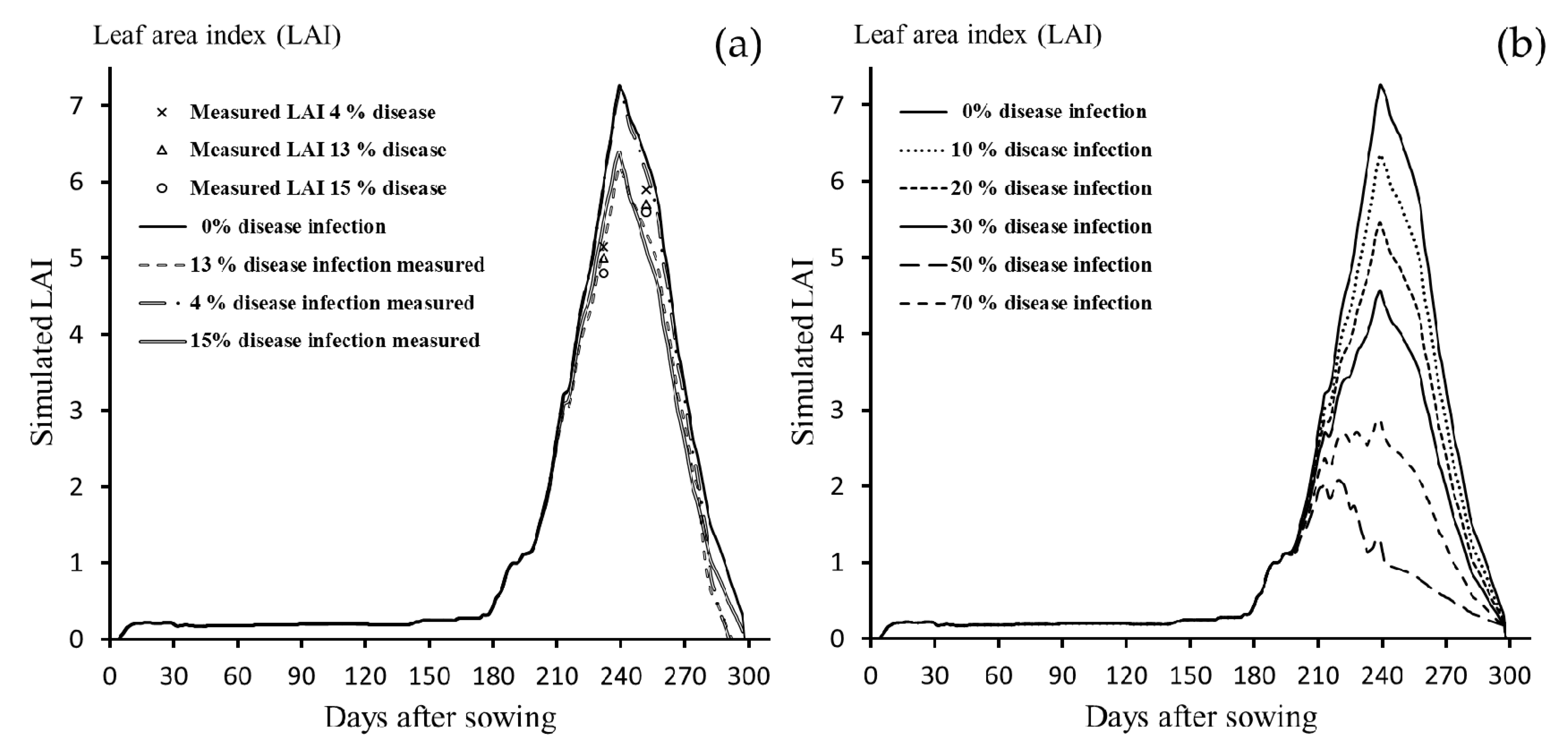
3.1.1. Leaf Area Index
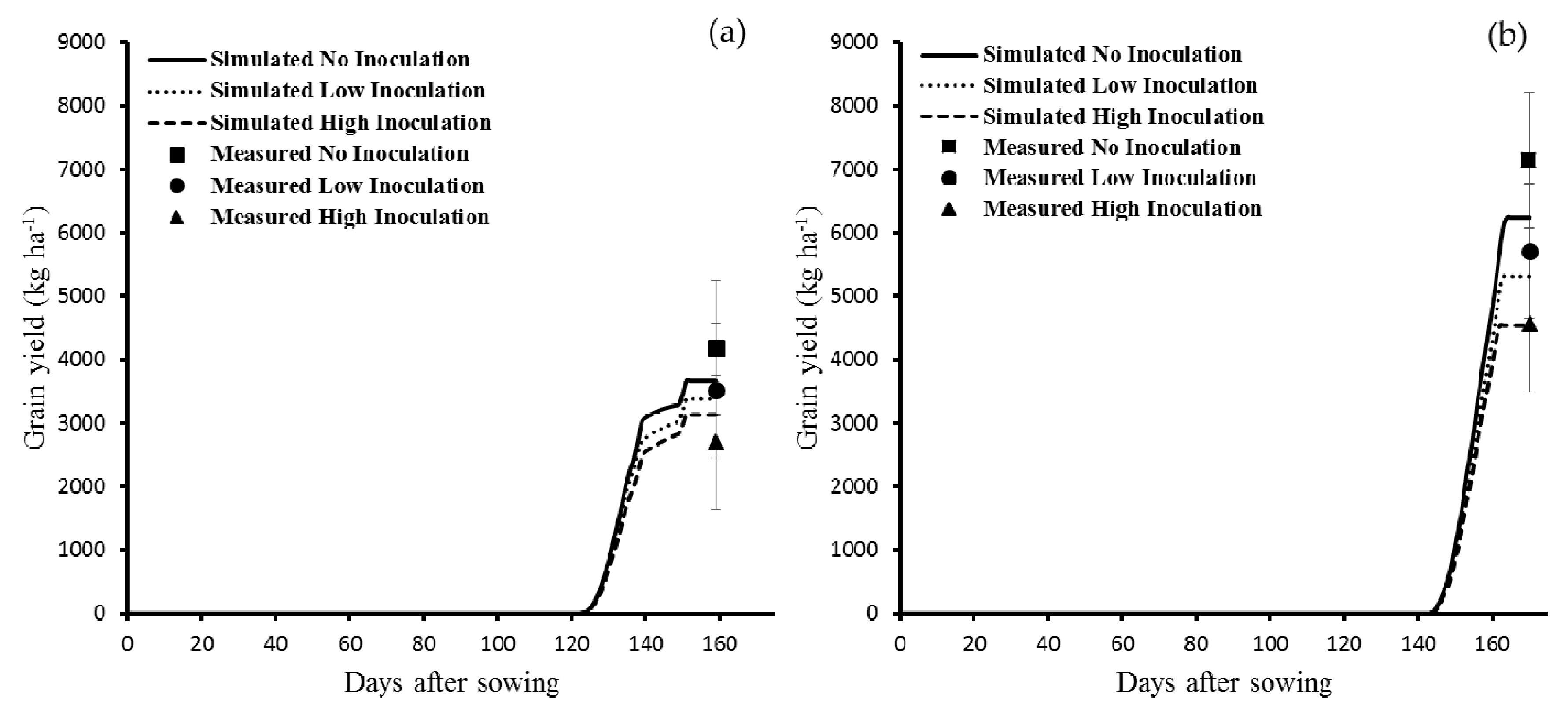
3.1.2. Yield
3.2. Model Validation for La Plata
3.2.1. LAI
3.2.2. Yield
3.3. Sensitivity Analysis
3.4. Future Model Applications
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Singh, R.P.; Singh, P.K.; Rutkoski, J.; Hodson, D.P.; He, X.; Jørgensen, L.N.; Hovmøller, M.S.; Huerta-Espino, J. Disease Impact on Wheat Yield Potential and Prospects of Genetic Control. Annu. Rev. Phytopathol. 2016, 54, 303–322. [Google Scholar] [CrossRef] [PubMed]
- Olesen, J.E.; Trnka, M.; Kersebaum, K.C.; Skjelvåg, A.O.; Seguin, B.; Peltonen-Sainio, P.; Rossi, F.; Kozyra, J.; Micale, F. Impacts and adaptation of European crop production systems to climate change. Eur. J. Agron. 2011, 34, 96–112. [Google Scholar] [CrossRef]
- Eigenbrode, S.D.; Binns, W.P.; Huggins, D.R. Confronting Climate Change Challenges to Dryland Cereal Production: A Call for Collaborative. Prod. Engagem. 2018, 5. [Google Scholar] [CrossRef]
- Ahanger, R.A.; Bhat, H.A.; Bhat, T.A.; Ganie, S.A.; Lone, A.A.; Wani, I.A.; Ganai, S.A.; Haq, S.; Khan, O.A.; Junaid, M.J.; Bhat, T.A. Impact of Climate Change on Plant Diseases. Int. J. Modern Plant & Anim. Sci. USA 2013, 3, 105–115. [Google Scholar]
- Weiss, M.V. Compendium of Wheat Diseases, 2nd ed.; APS Press: St. Paul, MN, USA, 1987. [Google Scholar]
- Forrer, H.R.; Zadoks, J.C. Yield reduction in wheat in relation to leaf necrosis caused by Septoria tritici. Neth. J. Plant Pathol. 1983, 89, 87–98. [Google Scholar] [CrossRef]
- Eyal, Z. The septoria tritici and stagonospora nodorum blotch diseases of wheat. Eur. J. Plant Pathol. 1999, 105, 629–641. [Google Scholar] [CrossRef]
- Bearchell, S.J.; Fraaije, B.A.; Shaw, M.W.; Fitt, B.D.L.; Cowling, E.B. Wheat Archive Links Long-Term Fungal Pathogen Population Dynamics to Air Pollution. Proc. Natl. Acad. Sci. USA 2005, 102, 5438–5442. [Google Scholar] [CrossRef] [PubMed]
- Eyal, Z.; Amiri, Z.; Wahl, I. Physiological Specialization of Septoria tritici. Phytopathology 1973, 63, 1087–1091. [Google Scholar] [CrossRef]
- Fones, H.; Gurr, S. The impact of Septoria tritici Blotch disease on wheat: An EU perspective. Fungal Genet. Biol. 2015, 79, 3–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraaije, B.; Cools, H.J.; Fountaine, J.; Lovell, D.J.; Motteram, J.; West, J.S.; Lucas, J. A Role of Ascospores in Further Spread of QoI-Resistant Cytochrome b Alleles (G143A) in Field Populations of Mycosphaerella graminicola. Phytopathology 2005, 95, 933–941. [Google Scholar] [CrossRef] [PubMed]
- Estep, L.K.; Torriani, S.F.F.; Zala, M.; Anderson, N.P.; Flowers, M.D.; Mcdonald, B.A.; Mundt, C.C.; Brunner, P.C. Emergence and early evolution of fungicide resistance in North American populations of Zymoseptoria tritici. Plant Pathol. 2015, 64, 961–971. [Google Scholar] [CrossRef] [Green Version]
- Simón, M.R.; Cordo, C.A.; Castillo, N.S.; Struik, P.C.; Börner, A. Population Structure of Mycosphaerella graminicola and Location of Genes for Resistance to the Pathogen: Recent Advances in Argentina. Int. J. Agron. 2012, 2012, 680275. [Google Scholar] [CrossRef]
- Rodrigo, S.; Cuello-Hormigo, B.; Gomes, C.; Santamaria, O.; Costa, R.; Poblaciones, M.J. Influence of fungicide treatments on disease severity caused by Zymoseptoria tritici, and on grain yield and quality parameters of bread-making wheat under Mediterranean conditions. Eur. J. Plant Pathol. 2014, 141, 99–109. [Google Scholar] [CrossRef]
- Keating, B.A.; Carberry, P.S.; Hammerb, G.L.; Probert, M.E.; Robertson, M.J.; Holzworth, D.; Huth, N.I.; Hargreaves, J.N.G.; Meinke, H.; Hochman, Z.; et al. An overview of the crop model APSIM. Eur. J. Agron. 2003, 18, 267–288. [Google Scholar] [CrossRef]
- Lv, Z.; Liu, X.; Cao, W.; Zhu, Y. Agricultural and Forest Meteorology Climate change impacts on regional winter wheat production in main wheat production regions of China. Agric. For. Meteorol. 2013, 171–172, 234–248. [Google Scholar] [CrossRef]
- Brisson, N.; Gary, C.; Justes, E.; Roche, R.; Mary, B.; Ripoche, D.; Zimmer, D.; Sierra, J.; Bertuzzi, P.; Burger, P.; et al. An overview of the crop model Stics. Eur. J. Agron. 2003, 18, 309–332. [Google Scholar] [CrossRef]
- Jamieson, P.D.; Semenov, M.A.; Brooking, I.R.; Francis, G.S. Sirius a mechanistic model of wheat respinse to environmental variation. Eur. J. Agron. 1998, 8, 161–179. [Google Scholar] [CrossRef]
- Jones, J.W.; Hoogenboom, G.; Porter, C.H.; Boote, K.J.; Batchelor, W.D.; Hunt, L.A.; Wilkens, P.W.; Singh, U.; Gijsman, A.J.; Ritchie, J.T. The Dssat Cropping System Model. Eur. J. Agron. 2003, 18, 235–265. [Google Scholar] [CrossRef]
- Hunt, L.A.; Pararajasingham , S. CROPSIM—WHEAT: A model describing the growth and development of wheat. Can. J. Plant Sci. 1995, 619–632. [Google Scholar]
- Ritchie, J.T.; Singh, U.; Godwin, D.C.; Bowen, W.T. Cereal growth, development and yield. Underst. Opt. Agric. Prod. 1998, 79–98. [Google Scholar] [CrossRef]
- Ritchie, J.T.; Otter, S. Description and performance of CERES-Wheat: A user-orientes wheat yield model. ARS Wheat Yield Proj. 1985, 38, 159–175. [Google Scholar]
- Hoogenboom, G.; Jones, J.W.; Wilkens, P.W.; Porter, C.H.; Boote, K.J.; Hunt, U.S.; Lizaso, J.I.; White, J.W.; Uryasev, O.; Ogoshi, R.; et al. Decision Support System for Agrotechnology Transfer (DSSAT) [CD-ROM]; University of Hawaii: Honululu, HI, USA, 2015. [Google Scholar]
- Thorp, K.R.; Hunsaker, D.J.; French, A.N.; White, J.W.; Clarke, T.R. Evaluation of the CSM-CROPSIM-CERES-Wheat Model as a Tool for Crop Water Management. Trans. ASABE 2010, 53, 1–17. [Google Scholar] [CrossRef]
- Chipanshi, A.C.; Ripley, E.A.; Lawford, R.G. Large-scale simulation of wheat yields in a semi-arid environment using a crop-growth model. Agric. Syst. 1999, 59, 57–66. [Google Scholar] [CrossRef]
- Savin, R.; Satorre, E.H.; Hall, A.J.; Slafer, G.A. Assessing strategies for wheat cropping in the monsoonal climate of the Pampas using the CERES-Wheat simulation model. Field Crops Res. 1995, 42, 81–91. [Google Scholar] [CrossRef]
- Sardinia, S.; Dettori, M.; Cesaraccio, C.; Motroni, A.; Spano, D.; Duce, P. Field Crops Research Using CERES-Wheat to simulate durum wheat production and phenology. Field Crops Res. 2011, 120, 179–188. [Google Scholar] [CrossRef]
- Bannayan, M.; Crout, N.M.J.; Hoogenboom, G. Application of the CERES-Wheat model for within-season prediction of winter wheat yield in the United Kingdom. Agron. J. 2003, 95, 114–125. [Google Scholar] [CrossRef]
- Gbegbelegbe, S.; Cammarano, D.; Asseng, S.; Robertson, R.; Chung, U.; Adam, M.; Abdalla, O.; Payne, T.; Reynolds, M.; Sonder, K.; et al. Baseline simulation for global wheat production with CIMMYT mega-environment specific cultivars. Field Crops Res. 2017, 202, 122–135. [Google Scholar] [CrossRef]
- Batchelor, W.D.; Jones, J.W.; Boote, K.J.; Porter, C.H. Pest and Disease Damage Module; University of Florida: Gainsville, FL, USA, 2004. [Google Scholar]
- Boote, K.J.; Bennet, J.M.; Jones, J.W.; Jowers, H.E. On-farming testing of peanut and soybean models in north Florida. Paper Am. Soc. Agric. Eng. USA 1989. Available online: http://agris.fao.org/agris-search/search.do?recordID=US9165910 (accessed on 10 January 2019).
- Batchelor, W.D.; Jones, J.W.; Boote, K.J.; Pinnschmidt, H.O. Extending the use of crop models to study pest damage. Trans. Am. Soc. Agric. Eng. Gen. Ed. 1993, 36, 551–558. [Google Scholar] [CrossRef]
- Boote, K.J.; Jones, J.W.; Hoogenboom, G.; Pickering, N.B. The CROPGRO model for grain legumes. In Understanding Options for Agricultural Production; Tsuji, G.Y., Hoogenboom, G., Thornton, P.K., Eds.; Springer: Dordrecht, The Netherlands, 1998; pp. 99–128. ISBN 978-94-017-3624-4. [Google Scholar]
- Andarzian, B.; Hoogenboom, G.; Bannayan, M.; Shirali, M.; Andarzian, B. Determining optimum sowing date of wheat using CSM-CERES-Wheat model. J. Saudi Soc. Agric. Sci. 2015, 14, 189–199. [Google Scholar] [CrossRef] [Green Version]
- Waggoner, P.E.; Berger, R.D. Defoliation, Disease, and Growth. Phytopathology 1987, 77, 1495–1497. [Google Scholar]
- Robert, C. Analysis and modelling of effects of leaf rust and Septoria tritici blotch on wheat growth. J. Exp. Bot. 2004, 55, 1079–1094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robert, C.; Bancal, M.O.; Lannou, C.; Ney, B. Quantification of the effects of Septoria tritici blotch on wheat leaf gas exchange with respect to lesion age, leaf number, and leaf nitrogen status. J. Exp. Bot. 2006, 57, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Vallet, A.; McDonald, M.C.; Solomon, P.S.; McDonald, B.A. Is Zymoseptoria tritici a hemibiotroph? Fungal Genet. Biol. 2015, 79, 29–32. [Google Scholar] [CrossRef] [PubMed]
- Castro, A.C.; Simón, M.R. Effect of tolerance to Septoria tritici blotch on grain yield, yield components and grain quality in Argentinean wheat cultivars. Crop Prot. 2016, 90, 66–76. [Google Scholar] [CrossRef]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Gröll, K. Use of Sensor Technologies to Estimate and Assess the Effects of Various Plant Diseases on Crop Growth and Development; Universty of Hohenheim: Stuttgart, Germany, 2008; Available online: http://opus.uni-hohenheim.de/volltexte/2008/296/ (accessed on 10 January 2019).
- Thomas, M.; Cook, R.; King, J. Factors affecting development of Septoria tritici in winter wheat and its effect on yield. Plant Pathol. 1989, 246–257. [Google Scholar] [CrossRef]
- Willmott, C.J. Some Comments on the Evaluation of Model Performance. J. Appl. Phys. 1982, 36, 1309–1313. [Google Scholar] [CrossRef]
- Jing, Q.; Qian, B.; Shang, J.; Huffman, T.; Liu, J.; Pattey, E.; Dong, T.; Tremblay, N.; Drury, C.F.; Ma, B.L.; et al. Assessing the options to improve regional wheat yield in eastern canada using the csm–ceres–wheat model. Agron. J. 2017, 109, 510–523. [Google Scholar] [CrossRef]
- Attia, A.; Rajan, N.; Xue, Q.; Nair, S.; Ibrahim, A.; Hays, D. Application of DSSAT-CERES-Wheat model to simulate winter wheat response to irrigation management in the Texas High Plains. Agric. Water Manag. 2016, 165, 50–60. [Google Scholar] [CrossRef]
- Robert, C.; Fournier, C.; Andrieu, B.; Ney, B. coupling a 3D virtual wheat (Triticum aestivum) plant model with a Septoria tritici epidemic model (Septo3D): A new approach to investigate plant-pathogen interactions linked to caonpy architecture. Funct. Plant Biol. 2008, 35, 997–1013. [Google Scholar] [CrossRef]
- Magarey, R.D.; Sutton, T.B.; Thayer, C.L. A Simple Generic Infection Model for Foliar Fungal Plant Pathogens. Phytopathology 2005, 95, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Donatelli, M.; Magarey, R.D.; Bregaglio, S.; Willocquet, L.; Whish, J.P.M.; Savary, S. Modelling the impacts of pests and diseases on agricultural systems. Agric. Syst. 2017, 155, 213–224. [Google Scholar] [CrossRef] [PubMed]
- King, J.E.; Jenkins, J.E.E.; Morgan, W.A. The estimation of yield losses in wheat from severity of infection by Septoria species. Plant Pathol. 1983, 32, 239–249. [Google Scholar] [CrossRef]
- Ziv, O.; Eyal, Z. Assessment of Yield Component Losses Caused in Plants of Spring Wheat Cultivars by Selected Isolates of Septoria tritici. Phytopathology 1977, 68, e796. [Google Scholar] [CrossRef]
- Bhathal, J.S.; Loughman, R.; Speijers, J. Yield reduction in wheat in relation to leaf disease from yellow (tan) spot and septoria nodorum blotch. Eur. J. Plant Pathol. 2003, 109, 435–443. [Google Scholar] [CrossRef]
- Bastiaans, L. Ecology and Epidemiology Ratio Between Virtual and Visual Lesion Size as a Measure to Describe Reduction in Leaf Photosynthesis of Rice Due to Leaf Blast. Phytopathology 1991, 81, 611–615. [Google Scholar] [CrossRef]
- Thorp, K.R.; Hunsaker, D.J.; French, A.N. Assimilating leaf area index estimates from remote sensing into the simulations of a cropping systems model. Trans. ASABE 2010, 53, 251–262. [Google Scholar] [CrossRef]
- Ficke, A.; Cowger, C.; Bergstrom, G.; Brodal, G. Understanding Yield Loss and Pathogen Biology to Improve Disease Management: Septoria Nodorum Blotch—A Case Study in Wheat. Plant Dis. 2018, 102, 696–707. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location La Plata | Clay Content % | Sand Content % | Silt Content % | LLL * | DUL ** | SAT *** |
|---|---|---|---|---|---|---|
| 0–30 cm | 20.7 | 28.9 | 50.4 | 0.226 | 0.457 | 0.561 |
| 30–60 cm | 20.7 | 28.9 | 50.4 | 0.226 | 0.457 | 0.561 |
| 60–90 cm | 20.7 | 28.9 | 50.4 | 0.226 | 0.457 | 0.561 |
| Location Ihinger Hof | ||||||
| 0–30 cm | 43.3 | 9.9 | 46.8 | 0.247 | 0.412 | 0.467 |
| 30–60 cm | 43.3 | 9.9 | 46.8 | 0.247 | 0.412 | 0.467 |
| 60–90 cm | 25.0 | 18.8 | 56.2 | 0.142 | 0.313 | 0.503 |
| Ihinger Hof | La Plata | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Parameter | Definition | Monopol | ACA801 | B75 Aniversario | Buck Brasil | Buck Guapo * Baguette 10 * Klein Zorro * Relmó Centinela * | Klein Escorpion | Klein Flecha | Klein Chaja |
| Cultivar file | |||||||||
| P1D | Percentage reduction in development rate in a photoperiod 10 h shorter than the threshold relative to that at the threshold | 110 | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| P1V | Days at optimum vernalizing temperature required to complete vernalisation | 90 | 20 | 20 | 20 | 20 | 20 | 20 | 20 |
| G1 | Kernel number per unit canopy weight at anthesis (kernels g−1) | 15 | 25 | 25 | 20 | 22 | 22 | 22 | 25 |
| G2 | Standard kernel size under optimum conditions (mg) | 48 | 25 | 30 | 30 | 30 | 35 | 30 | 39 |
| G3 | Standard, non-stressed dry weight of a single tiller at maturity (g) | 2.0 | 4.0 | 4.0 | 4.0 | 4.0 | 4.0 | 4.0 | 4.0 |
| P5 | Duration of the grain filling phase (°C d) | 600 | 420 | 430 | 420 | 420 | 420 | 420 | 420 |
| PHINT | Interval between successive leaf tip appearances (°C d) | 90 | 92 | 92 | 92 | 92 | 80 | 93 | 92 |
| Ecotype file | |||||||||
| P1 | Duration of phase end juvenile to terminal spikelet | 350 | 350 | 350 | 350 | 350 | 350 | 350 | 350 |
| P2 | Duration of phase from double ridges to the end of leaf growth (°C d) | 200 | 250 | 250 | 250 | 250 | 250 | 250 | 250 |
| P3 | Duration of phase from the end of leaf growth to the end of spike growth (°C d) | 300 | 220 | 220 | 220 | 220 | 220 | 220 | 220 |
| P4 | Duration of phase from the end of spike growth to the end of the grain fill lag (°C d) | 380 | 300 | 300 | 300 | 300 | 300 | 300 | 300 |
| PARUE | Conversion rate from photosynthetically active radiation to dry matter before the last leaf stage (g MJ−1) | 2.3 | 2.7 | 2.7 | 2.7 | 2.7 | 2.7 | 2.7 | 2.7 |
| SLAS | Specific leaf area, standard first leaf (cm2/g) | 400 | 450 | 450 | 450 | 450 | 450 | 450 | 450 |
| Original CCW | Modified CCW | ||||||
|---|---|---|---|---|---|---|---|
| Variable | Experiment | RMSE | d-Index | EF | RMSE | d-Index | EF |
| Leaf area index | La Plata 2010 | 1.19 | 0.33 | −2.69 | 0.69 | 0.51 | −1.07 |
| La Plata 2011 | 2.88 | 0.24 | −0.98 | 1.11 | 0.70 | 0.68 | |
| Yield | La Plata 2010 | 1144 | 0.47 | −1.19 | 499 | 0.81 | 0.58 |
| La Plata 2011 | 1755 | 0.50 | −1.19 | 1285 | 0.66 | −0.18 | |
| % Disease Infection | Simulated Yield kg ha−1 | Measured Yield kg ha−1 | % Yield Reduction | % LAI Reduction at Maximum LAI |
|---|---|---|---|---|
| 0 | 4384 | 0 | 0 | |
| 4 * | 4332 | 4409 | 1.2 | 0.8 |
| 10 | 4190 | 4.4 | 12.5 | |
| 13 * | 4159 | 3934 | 5.1 | 14.5 |
| 15 * | 4124 | 3965 | 5.9 | 12.0 |
| 20 | 3990 | 9.0 | 24.8 | |
| 30 | 3737 | 14.8 | 37.1 | |
| 50 | 3242 | 26.0 | 60.3 | |
| 70 | 2380 | 45.7 | 82.4 |
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Röll, G.; Batchelor, W.D.; Castro, A.C.; Simón, M.R.; Graeff-Hönninger, S. Development and Evaluation of a Leaf Disease Damage Extension in Cropsim-CERES Wheat. Agronomy 2019, 9, 120. https://doi.org/10.3390/agronomy9030120
Röll G, Batchelor WD, Castro AC, Simón MR, Graeff-Hönninger S. Development and Evaluation of a Leaf Disease Damage Extension in Cropsim-CERES Wheat. Agronomy. 2019; 9(3):120. https://doi.org/10.3390/agronomy9030120
Chicago/Turabian StyleRöll, Georg, William D. Batchelor, Ana Carolina Castro, María Rosa Simón, and Simone Graeff-Hönninger. 2019. "Development and Evaluation of a Leaf Disease Damage Extension in Cropsim-CERES Wheat" Agronomy 9, no. 3: 120. https://doi.org/10.3390/agronomy9030120
APA StyleRöll, G., Batchelor, W. D., Castro, A. C., Simón, M. R., & Graeff-Hönninger, S. (2019). Development and Evaluation of a Leaf Disease Damage Extension in Cropsim-CERES Wheat. Agronomy, 9(3), 120. https://doi.org/10.3390/agronomy9030120