Impact of Row Spacing, Sowing Density and Nitrogen Fertilization on Yield and Quality Traits of chia (Salvia Hispanica L.) Cultivated in southwestern Germany


Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Trial Setup
2.2.1. Row Spacing Trial
2.2.2. Sowing Density Trial
2.2.3. Fertilizer Trial
2.3. Experimental Procedure
2.3.1. Soil Mineral Nitrogen
2.3.2. Yield Traits
2.3.3. Quality Traits
2.4. Statistical Analysis
3. Results and Discussion
3.1. Plant Development
Impact of Day Length and Temperature
3.2. Yield Traits
3.2.1. Row Spacing Trial
3.2.2. Sowing Density Trial
3.2.3. N-Fertilizer Trial
3.2.4. Additional Factors Influencing Chia Seed Yield
3.3. Quality Traits
3.3.1. Row Spacing Trial
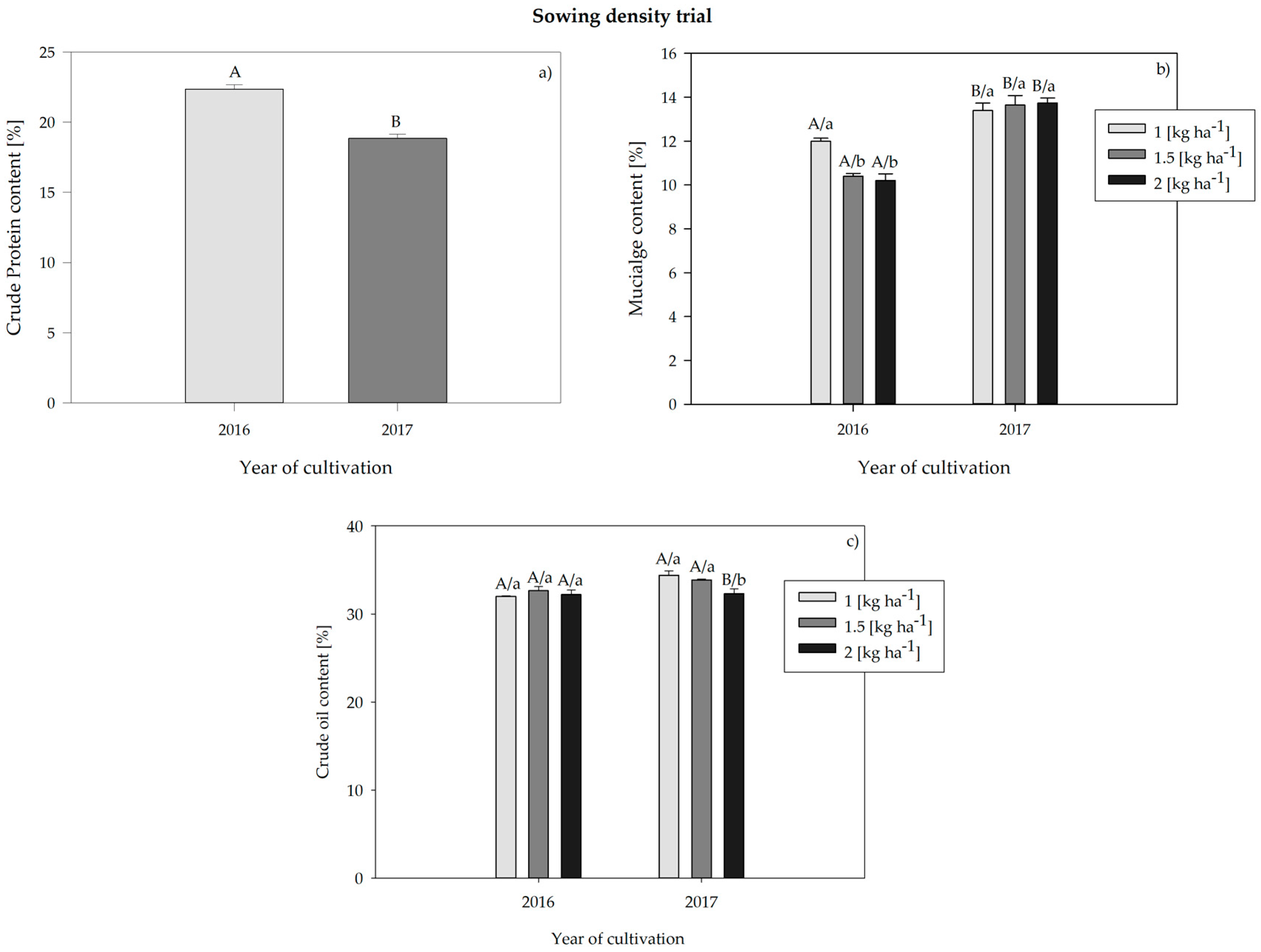
3.3.2. Sowing Density Trial
3.3.3. Fertilizer Trial
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jamboonsri, W.; Phillips, T.D.; Geneve, R.L.; Cahill, J.P.; Hildebrand, D.F. Extending the range of an ancient crop, Salvia hispanica L.—A new ω3 source. Genet. Resour. Crop Evol. 2012, 59, 171–178. [Google Scholar] [CrossRef]
- Mohd Ali, N.; Yeap, S.K.; Ho, W.Y.; Beh, B.K.; Tan, S.W.; Tan, S.G. The promising future of chia, Salvia hispanica L. J. Biomed. Biotechnol. 2012, 2012, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Us-Medina, U.; Julio, L.M.; Segura-Campos, M.R.; Ixtaina, V.Y.; Tomás, M.C. Development and characterization of spray-dried chia oil microcapsules using by-products from chia as wall material. Powder Technol. 2018, 334, 1–8. [Google Scholar] [CrossRef]
- Fernández-Ferrína, P.; Calvo-Turrientes, A.; Bande, B.; Artaraz-Miñóna, M.; Galán-Ladero, M.M. The valuation and purchase of food products that combine local, regional and traditional features: The influence of consumer ethnocentrism. Food Qual. Prefer. 2018, 64, 138–147. [Google Scholar] [CrossRef]
- Grimes, S.; Phillips, T.; Claupein, W.; Graeff-Hönninger, S. Chia^4: Is a new tetraploid chia (Salvia hispanica L.) genotype able to revolutionize European agriculture? In The Food System Approach: New Challenges for Education, Research and Industry; ISEKI Food Association: Stuttgart, Germany, 2018; p. 79. [Google Scholar]
- Hildebrand, D.; Jamboonsri, W. Phillips Early Flowering Chía and Uses Thereof. U.S. Patent 8586831, 19 October 2013. [Google Scholar]
- Grimes, S.J.; Phillips, T.D.; Hahn, V.; Capezzone, F.; Graeff-Hönninger, S. Growth, Yield Performance and Quality Parameters of Three Early Flowering Chia (Salvia Hispanica L.) Genotypes Cultivated in Southwestern Germany. Agriculture 2018, 8, 154. [Google Scholar] [CrossRef]
- Bochicchio, R.; Phillips, T.D.; Lovelli, S.; Labella, R.; Galgano, F.; Marisco, A.D.; Perniola, M.; Amato, M. Innovative Crop Productions for Healthy Food: The Case Chia (Salvia hispanica L.). In The Sustainability of Agro-Food and Natural Resource Systems in the Mediterranean Basin; Vastola, A., Ed.; Springer International Publishing: Basel, Switzerland, 2015; pp. 29–45. ISBN 9783319163574. [Google Scholar]
- Souza, R.S.; Chaves, L.H.G. Initial growth of chia (Salvia hispanica L.) submitted to nitrogen, phosphorus and potassium fertilization. Aust. J. Crop Sci. 2017, 11, 610–615. [Google Scholar] [CrossRef]
- Thiago, F.d.F.; Bruna, B.R.; Alexandre, L.M.; Dyogo, B.B.; Sidiane, C.-R.; Omari, D.F.D.; Guilherme, P.d.F.; Heloisa, F.C.M.-M. Development and production of chia (Salvia hispanica L.) in different space arrangements. Afr. J. Agric. Res. 2016, 11, 4384–4388. [Google Scholar] [CrossRef]
- Wojahn, R.E.; Bortolotto, R.P.; Zamberlan, J.F.; Koefender, J.; Tragnago, J.L.; Camera, J.N.; Pasini, M.P.B.; Salazar, R.F.S.; Damiani, F. Agronomic feasibility of growing chia in northwestern rio grande do sul. Holos 2018, 3, 112–122. [Google Scholar] [CrossRef]
- Bochicchio, R.; Rossi, R.; Labella, R.; Bitella, G.; Perniola, M.; Amato, M. Effect of sowing density and nitrogen top-dress fertilisation on growth and yield of chia (Salvia hispanica L.) in a mediterranean environment: First results. Ital. J. Agron. 2015, 10, 163–166. [Google Scholar] [CrossRef]
- Bilalis, D.; Tabaxi, I.; Zervas, G.; Tsiplakou, E.; Travlos, I.S.; Kakabouki, I.; Tsioros, S. Chia (Salvia Hispanica) Fodder Yield and Quality as Affected by Sowing Rates and Organic Fertilization. Commun. Soil Sci. Plant Anal. 2016, 47. [Google Scholar] [CrossRef]
- Karkanis, A.C.; Kontopoulou, C.K.; Lykas, C.; Kakabouki, I.; Petropoulos, S.A.; Bilalis, D. Efficacy and selectivity of pre- and post-emergence herbicides in chia (Salvia hispanica L.) under mediterranean semi-arid conditions. Not. Bot. Horti Agrobot. Cluj-Napoca 2018, 46, 183–189. [Google Scholar] [CrossRef]
- Mcmaster, G.S.; Wilhelm, W.W. Growing degree-days: One equation, two interpretations. Agric. For. Meteorol. 1997, 87, 291–300. [Google Scholar] [CrossRef]
- IUSS Working Group WRB. World reference base for soil resources 2006, first update 2007 A framework for international classification, correlation and communication. In World Soil Resources Reports No. 103; FAO: Rome, Italiy, 2007; pp. 1–128. ISBN 9789251055113. [Google Scholar]
- Bassler, R.; Hoffmann, G. Bestimmung von mineralischem, (Nitrat-) Stickstoff in Bodenprofilen, Nmin-Labormethode. In VDLUFA Methodenbuch Band I.; Deller, B., Ed.; VDLUFA-Verlag: Darmstadt, Germany, 1997; ISBN 978-3-941273-21-4. [Google Scholar]
- Muñoz, L.A.; Cobos, A.; Diaz, O.; Aguilera, J.M. Chia seeds: Microstructure, mucilage extraction and hydration. J. Food Eng. 2012, 108, 216–224. [Google Scholar] [CrossRef]
- Dumas, A. Stickstoffbestimmung nach Dumas. In Die Praxis des org. Chemikers; Gattermann-Wieland, Ed.; Walter de Gruyter: Nürnberg, Germany, 1962. [Google Scholar]
- AOAC. Micro-Kjeldahl Method. In Official Methods of Analysis; Cunniff, P., Ed.; Association of Official Analytical Chemists: Arlington VA, USA, 1995. [Google Scholar]
- European Commission. Regulation (EC). No. 152/III H procedure B laying down the methods of sampling and analysis for the official control of feed. Off. J. Eur. Union 2009, 54, 1–130. [Google Scholar]
- Metcalfe, L.D.; Schmitz, A.A.; Pelka, J.R. Rapid preparation of fatty acid esters from lipids for gas chromatographic Analysis. Anal. Chem. 1966, 38, 514–515. [Google Scholar] [CrossRef]
- DGF Fettsäuremethylester. In Einheitsmethoden zur Untersuchung von Fetten, Fettprodukten, Tensiden und verwandten Stoffen, 2nd ed.; Deutsche Gesellschaft für Fettwissenschaft e.V. (Ed.) Wissenschaftliche Verlagsgesellschaft mbH: Stuttgart, Germany, 2002. [Google Scholar]
- Thurnhofer, S.; Vetter, W. A gas chromatography/electron ionization-mass spectrometry-selected ion monitoring method for determining the fatty acid pattern in food after formation of fatty acid methyl esters. J. Agric. Food Chem. 2005, 53, 8896–8903. [Google Scholar] [CrossRef] [PubMed]
- Roy, J. SAS for Mixed Models. In Journal of Biopharmaceutical Statistics; Littell, R.C., Milliken, G.A., Stroup, W.W., Wolfinger, R.D., Schabenberger, O., Eds.; Taylor & Francis: London, UK, 2007; Volume 17, pp. 363–365. ISBN 1054340060100. [Google Scholar]
- Baginsky, C.; Arenas, J.; Escobar, H.; Garrido, M.; Valero, N.; Tello, D.; Pizarro, L.; Valenzuela, A.; Morale, L.; Silva, H. Growth and yield of chia (Salvia hispanica L.) in the Mediterranean and desert climates of Chile. Chil. J. Agric. Res. 2016, 76, 255–264. [Google Scholar] [CrossRef] [Green Version]
- Ayerza, R.; Coates, W. New industrial crops: Northwestern Argentina Regional Project. In Progress in New Crops; Janick, J., Ed.; ASHS Press: Alexandria, Egypt, 1996; pp. 46–51. [Google Scholar]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration—Guidelines for Computing Crop Water Requirements; FAO: Rome, Italy, 1998; Volume 56. [Google Scholar]
- ESRL Solar Calculation Details. Available online: https://www.esrl.noaa.gov/gmd/grad/solcalc/calcdetails.html (accessed on 15 January 2018).
- Coates, W.; Ayerza, R. Commercial production of chia in Northwestern Argentina. JAOCS J. Am. Oil Chem. Soc. 1998, 75, 1417–1420. [Google Scholar] [CrossRef]
- Ayerza, R.; Coates, W. Some quality components of four chia (Salvia hispanica L.) genotypes grown under tropical coastal desert ecosystem conditions. Asian J. Plant Sci. 2009, 8, 301–307. [Google Scholar] [CrossRef]
- Yeboah, S.; Owusu Danquah, E.; Lamptey, J.N.L.; Mochiah, M.B.; Lamptey, S.; Oteng-Darko, P.; Adama, I.; Appiah-Kubi, Z.; Agyeman, K. Influence of Planting Methods and Density on Performance of Chia (Salvia hispanica) and its Suitability as an Oilseed Plant. Agric. Sci. 2014, 2, 14–26. [Google Scholar] [CrossRef] [Green Version]
- Mary, J.; Veeranna, H.K.; Girijesh, G.K.; Dhananjaya, B.C.; Gangaprasad, S. Effect of Different Spacings and Fertilizer Levels on Growth Parameters and Yield of Chia (Salvia hispanica L.). Int. J. Pure Appl. Biosci. 2018, 6, 259–263. [Google Scholar] [CrossRef]
- Kayaçetin, F.; Önemli, F.; Yilmaz, G.; Kinay, A.; Hatipoğlu, H.; Kivilcim, M.N.; Kara, N.; Köse, A.; Sefaoğlu, F. Effect of Row Spacing on Yield, Yield Components and Crude Oil of Autumn and Spring Sowed Mustard (Sinapis arvensis L.) in Eight Locations of Turkey. J. Agric. Sci. 2018, 24, 483–499. [Google Scholar]
- Rahnama, A.; Bakhshandeh, A. Determination of Optimum Row-Spacing and Plant Density for Uni-branched Sesame in Khuzestan Province. J. Agric. Sci. Technol. 2006, 8, 25–33. [Google Scholar]
- Ayerza, R.; Coates, W. Chia: Rediscovering a Forgotten Crop of the Aztecs; University of Arizona Press: Tucson, AZ, USA, 2005; ISBN 978-0-8165-2488-4. [Google Scholar]
- Pozo Pozo, S.A. Alternativas Para el Control Químico de Malezas Anuales en el Cultivo de la Chía (Salvia hispánica) en la Granja ECAA, Provincia de Imbabura; Pontificia Universidad Católica del Ecuador: Ibarra, Ecuador, 2010. [Google Scholar]
- Coates, W. Whole and Ground Chia (Salvia hispanica L.) Seeds, Chia Oil—Effects on Plasma Lipids and Fatty Acids; Elsevier Inc.: Amsterdam, The Netherlands, 2011; ISBN 9780123756886. [Google Scholar]
- Deligios, P.A.; Carboni, G.; Farci, R.; Solinas, S.; Ledda, L. Low-input herbicide management: Effects on rapeseed production and profitability. Sustainability 2018, 10, 2258. [Google Scholar] [CrossRef]
- Amato, M.; Caruso, M.C.; Guzzo, F.; Galgano, F.; Commisso, M.; Bochicchio, R.; Labella, R.; Favati, F. Nutritional quality of seeds and leaf metabolites of Chia (Salvia hispanica L.) from Southern Italy. Eur. Food Res. Technol. 2015, 241, 615–625. [Google Scholar] [CrossRef]
- Mamun, F.; Ali, M.H.; Chowdhury, I.F.; Hasanuzzaman, M.; Matin, M.A. Performance of Rapeseed Mustard Varieties Grown Under Different Plant Density. Sci. Agric. 2014, 4. [Google Scholar] [CrossRef]
- Raghuvanshi, N.; Kumar, V. Advances in agronomical manipulation on Indian mustard (Brassica juncea Cezern and Coss.): An review. J. Pharmacogn. Phytochem. 2018, 7, 833–837. [Google Scholar]
- Sosa-Baldivia, A.; Ruiz Ibarra, G. Inadequate Nitrogen Fertilization: Main Cause of the Low Seed Yield on the Chia Crop (Salvia Hispanica l.). Biomed. J. Sci. Tech. Res. 2018, 2, 2–4. [Google Scholar] [CrossRef]
- Pascual-Villalobos, M.; Correal, E.; Molina, E.; Martínez, J. Evaluación y Selección de Especies Vegetales Productoras de CompuestosNaturales con Actividad Insecticida; Centro de Investigación y Desarrollo Agroalimentario (CIDA): Murcia, Spain, 1997. [Google Scholar]
- Muñoz, L.A.; Cobos, A.; Diaz, O.; Aguilera, J.M. Chia Seed (Salvia hispanica): An Ancient Grain and a New Functional Food. Food Rev. Int. 2013, 29, 394–408. [Google Scholar] [CrossRef]
- Ouzounidou, G.; Skiada, V.; Papadopoulou, K.K.; Stamatis, N.; Kavvadias, V.; Eleftheriadis, E.; Gaitis, F. Effects of soil pH and arbuscular mycorrhiza (AM) inoculation on growth and chemical composition of chia (Salvia hispanica L.) leaves. Braz. J. Bot. 2015, 38, 487–495. [Google Scholar] [CrossRef]
- Alpaslan, M.; Boydak, E.; Hayta, M.; Gerçek, S.; Simsek, M. Effect of row space and irrigation on seed composition of Turkish sesame (sesamum indicum L.). JAOCS J. Am. Oil Chem. Soc. 2001, 78, 933–935. [Google Scholar] [CrossRef]
- Eryiğit, T.; Yıldırım, B.; Kumlay, A.M.; Sancaktaroğlu, S. The Effects of Different Row Distances and Nitrogen Fertilizer Rates on Yield and Yield Components of Safflower (Carthamus tinctorious) Under Micro-Climate Conditions of Iğdır Plain—Turkey. In Proceedings of the 3rd International Conference on Biological, Chemical & Environmental Sciences, Kuala Lumpur, Malaysia, 21–22 September 2015; pp. 21–22. [Google Scholar]
- Ayerza, R.; Coates, W. Protein content, oil content and fatty acid profiles as potential criteria to determine the origin of commercially grown chia (Salvia hispanica L.). Ind. Crops Prod. 2011, 34, 1366–1371. [Google Scholar] [CrossRef]
- Ayerza, R. The seed’s protein and oil content, fatty acid composition, and growing cycle length of a single genotype of chia (Salvia hispanica L.) as affected by environmental factors. J. Oleo Sci. 2009, 58, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Capitani, M.I.; Spotorno, V.; Nolasco, S.M.; Tomás, M.C. Physicochemical and functional characterization of by-products from chia (Salvia hispanica L.) seeds of Argentina. LWT Food Sci. Technol. 2012, 45, 94–102. [Google Scholar] [CrossRef]
- Ayerza, R.; Coates, W. Omega-3 enriched eggs: The influence of dietary α-linolenic fatty acid source on egg production and composition. Can. J. Anim. Sci. 2001, 81, 355–362. [Google Scholar] [CrossRef] [Green Version]
- Ixtaina, V.Y.; Martínez, M.L.; Spotorno, V.; Mateo, C.M.; Maestri, D.M.; Diehl, B.W.K.; Nolasco, S.M.; Tomás, M.C. Characterization of chia seed oils obtained by pressing and solvent extraction. J. Food Compos. Anal. 2011, 24, 166–174. [Google Scholar] [CrossRef]
- Silveira Coelho, M.; de las Mercedes Salas-Mellado, M. Chemical Characterization of CHIA (Salvia hispanica L.) for Use in Food Products. J. Food Nutr. Res. 2014, 2, 263–269. [Google Scholar] [CrossRef] [Green Version]
- Tavares, L.S.; Junqueira, L.A.; de Oliveira Guimarães, Í.C.; de Resende, J.V. Cold extraction method of chia seed mucilage (Salvia hispanica L.): Effect on yield and rheological behavior. J. Food Sci. Technol. 2018, 55, 457–466. [Google Scholar] [CrossRef] [PubMed]
- El-Satar, M.A.A.; Ahmed, A.A.; Hassan, T.H.A. Response of seed yield and fatty acid compositions for some sunflower genotypes to plant spacing and nitrogen fertilization. Inf. Process. Agric. 2017, 4, 241–252. [Google Scholar] [CrossRef]
- Ixtaina, V.Y.; Nolasco, S.M.; Tomás, M.C. Physical properties of chia (Salvia hispanica L.) seeds. Ind. Crops Prod. 2008, 28, 286–293. [Google Scholar] [CrossRef]
- European Commission. Commission implementing decision authorizing the placing on the market of chia oil (Salvia hispanica) as a novel food ingredient under Regulation (EC) No 258/97 of the European Parliament and of the Council. Off. J. Eur. Union 2014, 353, 15–16. [Google Scholar]
- Bellaloui, N.; Mengistu, A.; Walker, E.R.; Young, L.D. Soybean seed composition as affected by seeding rates and row spacing. Crop Sci. 2014, 54, 1782–1795. [Google Scholar] [CrossRef]
- Boydak, E.; Alpaslan, M.; Hayta, M.; Gerçek, S.; Simsek, M. Seed Composition of Soybeans Grown in the Harran Region of Turkey As Affected by Row Spacing and Irrigation. J. Agric. Food Chem. 2002, 50, 4718–4720. [Google Scholar] [CrossRef] [PubMed]
- Marcinek, K.; Krejpcio, Z. Chia Seeds (Salvia Hispanica): Health Promoting Properties and Therapeutic Applications—A Review. Rocz Panstw Zakl Hig 2017, 68, 123–129. [Google Scholar] [PubMed]
- Álvarez-Chávez, L.M.; Valdivia-López, M.D.L.A.; Aburto-Juárez, M.D.L.; Tecante, A. Chemical characterization of the lipid fraction of Mexican chia seed (Salvia hispanica L.). Int. J. Food Prop. 2008, 11, 687–697. [Google Scholar] [CrossRef]
- Ciftci, O.N.; Przybylski, R.; Rudzińska, M. Lipid components of flax, perilla, and chia seeds. Eur. J. Lipid Sci. Technol. 2012, 114, 794–800. [Google Scholar] [CrossRef]
- Peltonen-sainio, P.; Jauhiainen, L.; Hyövelä, M.; Nissilä, E. Trade-off between oil and protein in rapeseed at high latitudes: Means to consolidate protein crop status ? Field Crop Res. 2011, 121, 248–255. [Google Scholar] [CrossRef]
- Wang, N.; Daun, J.K. Effects of variety and crude protein content on nutrients and anti-nutrients in lentils (Lens culinaris ). Food Chem. 2006, 95, 493–502. [Google Scholar] [CrossRef]
- Ayerza, R.; Coates, W. Chia Seeds: New Source of Omega-3 Fatty Acids, Natural Antioxidants, and Dietetic Fiber; Southwest Center for Natural Products Research & Commercialization, Office of Arid Lands Studies: Tucson, AZ, USA, 2001. [Google Scholar]
- de Falco, B.; Incerti, G.; Bochicchio, R.; Phillips, T.D.; Amato, M.; Lanzotti, V. Metabolomic analysis of Salvia hispanica seeds using NMR spectroscopy and multivariate data analysis. Ind. Crops Prod. 2017, 99, 86–96. [Google Scholar] [CrossRef]
- Capitani, M.I.; Ixtaina, V.Y.; Nolasco, S.M.; Tomás, M.C. Microstructure, chemical composition and mucilage exudation of chia (Salvia hispanica L.) nutlets from Argentina. J. Sci. Food Agric. 2013, 93, 3856–3862. [Google Scholar] [CrossRef] [PubMed]
- Ayerza, R.; Coates, W. Composition of chia (Salvia hispanica) grown in six tropical and subtropical ecosystems of South America. Trop. Sci. 2004, 44, 131–135. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Depth (cm) | Sand (%) | Silt (%) | Clay (%) | pH | Nmin a (kg ha−1) |
|---|---|---|---|---|---|---|
| 2016 | 30 | 7.85 | 66.95 | 25.2 | 7.45 | 6.54 |
| 60 | 5.10 | |||||
| 90 | 11.32 | |||||
| 2017 | 30 | 2.72 | 70.18 | 27.1 | 6.63 | 59.46 |
| 60 | 2.41 | 70.44 | 27.15 | 27.58 | ||
| 90 | 3.29 | 63.61 | 33.1 | 13.24 |
| Sowing Date | Flower Induction | Radiation (Wh/m2) c | Day Length (h) d | Mean Temp. (°C) e | Mean Temp. (°C) f | Harvest Maturity * | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| DAS a | Date | GDD b | DAS | Date | GDD | |||||
| June 1st 2016 | 65 | August 4th 2016 | 463.3 | 10,184 | 15.8 | 17.3 | 14.2 | 150 | October 28th | 971.75 |
| May 17th 2017 | 68 | July 23rd 2017 | 529.75 | 10,316 | 15.9 | 18.0 | 14.4 | 156 | October 19th | 948.3 |
| Trait | Year | F-Test | ||||
|---|---|---|---|---|---|---|
| Row Spacing (cm) | 2016 | 2017 | ||||
| Seed yield (kg ha−1) ‡ | ||||||
| 35 | 1171.33 A/a | 819.67 B/a | ||||
| 50 | 1170.33 A/a | 844.33 B/a | 0.0017 | 0.0004 | 0.3809 | 0.9284 |
| 75 | 1110.0 A/a | 704.00 B/a | ||||
| SEM = 35.12 | SEM = 144.42 | |||||
| TSM [g] | ||||||
| 35 | 1.23 A/a | 1.17 B/a | ||||
| 50 | 1.23 A/a | 1.17 B/a | <0.0001 | 0.6242 | 0.8895 | 0.7997 |
| 75 | 1.24 A/a | 1.17 B/a | ||||
| SEM = 0.010 | ||||||
| Trait | Year | F-Test | ||||
|---|---|---|---|---|---|---|
| Sowing Density [kg ha−1] | 2016 | 2017 | ||||
| Seed yield [kg ha−1] | ||||||
| 1 | 618.33 A/a | 819.67 B/a | ||||
| 1.5 | 880.01 A/b | 844.33 B/b | 0.1122 | 0.0094 | 0.0114 | 0.4283 |
| 2 | 748.00 A/ab | 704.00 B/ab | ||||
| SEM = 55.12 | ||||||
| TSM [g] ‡ | ||||||
| 1 | 1.22 A/a | 1.21 A/a | ||||
| 1.5 | 1.20 A/b | 1.18 A/b | 0.1268 | 0.6378 | 0.0125 | 0.8024 |
| 2 | 1.20 A/ab | 1.19 A/ab | ||||
| SEM = 0.016 | SEM = 0.004 | |||||
| Trait | Year | F-Test | ||||
|---|---|---|---|---|---|---|
| Fertilizer Rate [kg N ha−1] | 2016 | 2017 | ||||
| Seed yield (kg ha−1) | ||||||
| 0 | 745.67 A/a | 768.33 A/a | ||||
| 20 | 751.67 A/a | 841.33 A/a | 0.4333 | 0.0020 | 0.4036 | 0.8123 |
| 40 | 838.00 A/a | 847.00 A/a | ||||
| SEM = 66.10 | ||||||
| TSM [g] ‡ | ||||||
| 0 | 1.19 A/a | 1.14 B/a | ||||
| 20 | 1.20 A/a | 1.17 B/a | 0.0319 | 0.3148 | 0.1297 | 0.1561 |
| 40 | 1.20 A/a | 1.19 B/a | ||||
| SEM = 0.004 | SEM = 0.012 | |||||
| Trait | Year 2016 | Year 2017 | F-Test | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Row Spacing [cm] | 35 | 50 | 75 | 35 | 50 | 75 | SEM | ||||
| Palmitic acid | 6.79 A/a | 6.76 A/a | 6.80 A/a | 7.16 B/a | 7.04 B/a | 7.18 B/a | 0.082 | <0.0001 | 0.0825 | 0.4589 | 0.7897 |
| Stearic acid | 3.30 A/a | 3.12 A/b | 3.18 A/b | 3.58 B/a | 3.23 B/b | 3.22 B/b | 0.061 | 0.0306 | 0.0587 | 0.0053 | 0.1824 |
| Oleic acid | 6.78 A/a | 6.30 A/b | 6.57 A/ab | 6.56 B/a | 6.14 B/b | 6.13 B/ab | 0.195 | 0.0386 | 0.0318 | 0.0052 | 0.6164 |
| Vaccenic acid | 0.75 A/a | 0.76 A/a | 0.77 A/b | 0.81 B/a | 0.82 B/a | 0.85 B//b | 0.005 | <0.0001 | 0.1480 | 0.0012 | 0.4652 |
| Linoleic acid | 20.53 A/a | 19.93 A/ab | 19.79 A/b | 19.91 B/a | 19.98 A/a | 20.14 A/a | 0.157 | 0.5521 | 0.0013 | 0.2234 | 0.0389 |
| α-Linolenic acid | 61.85 A/a | 63.13 A/b | 62.88 A/ab | 61.98 A/a | 62.80 A/b | 62.50 A/ab | 0.308 | 0.4329 | 0.0004 | 0.0125 | 0.6697 |
| SFA | 10.09 A/a | 9.88 A/b | 9.98 A/ab | 10.74 B/a | 10.27 B/b | 10.40 B/ab | 0.108 | 0.0002 | 0.0193 | 0.0273 | 0.4359 |
| MUFA | 7.53 A/a | 7.06 A/b | 7.35 A/b | 7.37 A/a | 6.96 A/b | 6.97 A/b | 0.193 | 0.0880 | 0.0332 | 0.0070 | 0.6380 |
| PUFA | 82.38 A/a | 83.06 A/b | 82.67 A/ab | 81.89 A/a | 82.78 A/b | 82.63 A/ab | 0.212 | 0.1304 | 0.0233 | 0.0093 | 0.5911 |
| PUFA/SFA | 8.18 A/a | 8.41 A/b | 8.29 A/ab | 7.63 B/a | 8.07 B/b | 7.95 B/ab | 0.098 | 0.0004 | 0.0065 | 0.0164 | 0.5179 |
| ω6:ω3 | 0.33 A/a | 0.32 A/a | 0.32 A/a | 0.32 A/a | 0.32 A/a | 0.32 A/a | 0.004 | 0.8434 | 0.0042 | 0.1309 | 0.1019 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grimes, S.J.; Phillips, T.D.; Capezzone, F.; Graeff-Hönninger, S. Impact of Row Spacing, Sowing Density and Nitrogen Fertilization on Yield and Quality Traits of chia (Salvia Hispanica L.) Cultivated in southwestern Germany. Agronomy 2019, 9, 136. https://doi.org/10.3390/agronomy9030136
Grimes SJ, Phillips TD, Capezzone F, Graeff-Hönninger S. Impact of Row Spacing, Sowing Density and Nitrogen Fertilization on Yield and Quality Traits of chia (Salvia Hispanica L.) Cultivated in southwestern Germany. Agronomy. 2019; 9(3):136. https://doi.org/10.3390/agronomy9030136
Chicago/Turabian StyleGrimes, Samantha J., Timothy D. Phillips, Filippo Capezzone, and Simone Graeff-Hönninger. 2019. "Impact of Row Spacing, Sowing Density and Nitrogen Fertilization on Yield and Quality Traits of chia (Salvia Hispanica L.) Cultivated in southwestern Germany" Agronomy 9, no. 3: 136. https://doi.org/10.3390/agronomy9030136
APA StyleGrimes, S. J., Phillips, T. D., Capezzone, F., & Graeff-Hönninger, S. (2019). Impact of Row Spacing, Sowing Density and Nitrogen Fertilization on Yield and Quality Traits of chia (Salvia Hispanica L.) Cultivated in southwestern Germany. Agronomy, 9(3), 136. https://doi.org/10.3390/agronomy9030136