Variability among Young Table Grape Cultivars in Response to Water Deficit and Water Use Efficiency


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Treatments
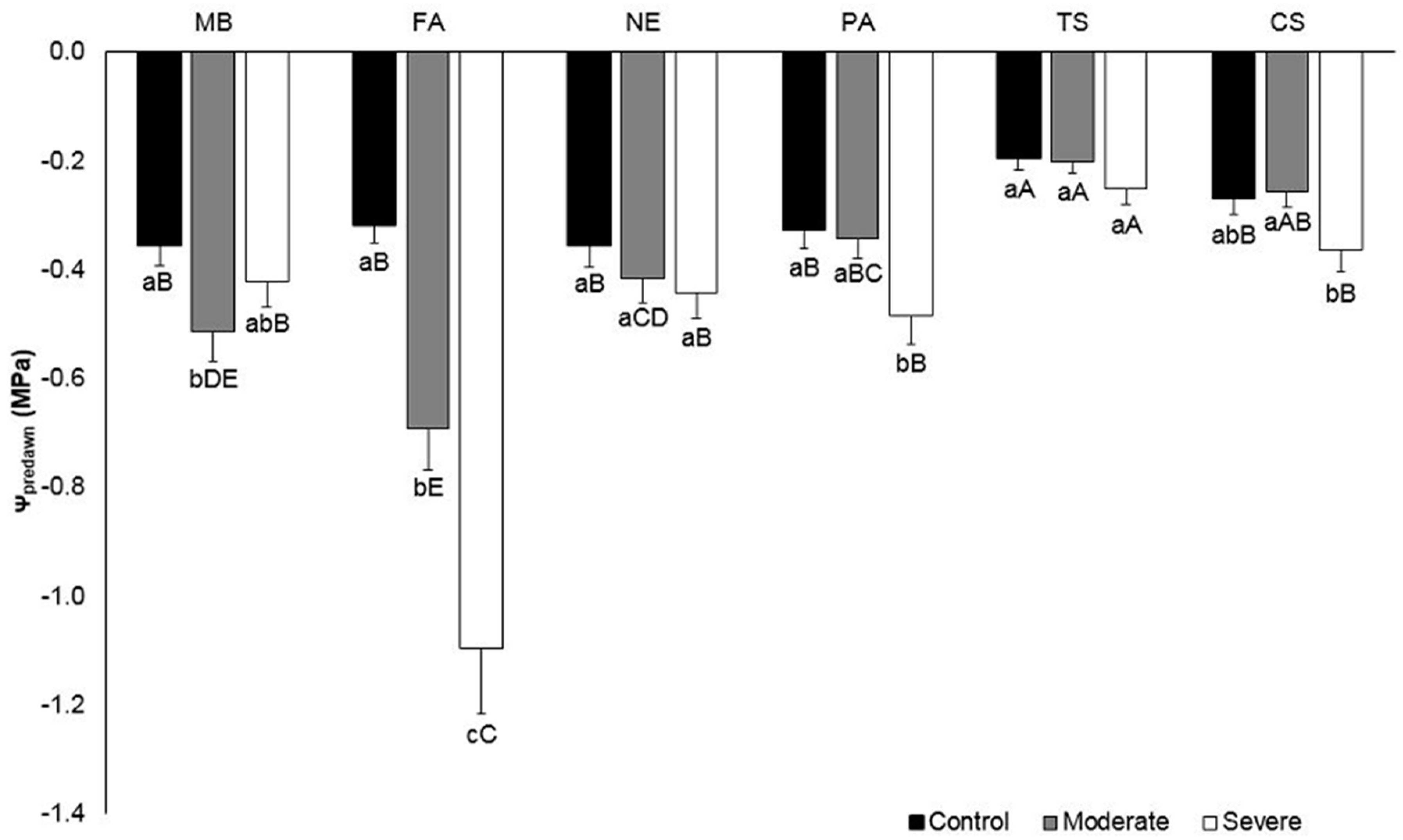
2.2. Plant Water Status
2.3. Gas Exchange Measurements
2.4. Plant Dry Weight and Leaf Area
2.5. Statistical Analysis
3. Results
3.1. Plant Water Status
3.2. Gas Exchange Measurement
3.3. Growth Parameters, Dry Mass Partitioning, and Plant WUE
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- IPCC. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2014. [Google Scholar]
- Costa, J.M.; Ortuño, M.F.; Chaves, M.M. Deficit irrigation as a strategy to save water: Physiology and potential application to horticulture. J. Integr. Plant Biol. 2007, 49, 1421–1434. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part B: Regional Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014; ISBN 9781107683860. [Google Scholar]
- Moriondo, M.; Bindi, M.; Fagarazzi, C.; Ferrise, R.; Trombi, G. Framework for high-resolution climate change impact assessment on grapevines at a regional scale. Reg. Environ. Chang. 2011, 11, 553–567. [Google Scholar] [CrossRef]
- Fraga, H.; Santos, J.A.; Malheiro, A.C.; Oliveira, A.A.; Moutinho-Pereira, J.; Jones, G.V. Climatic suitability of Portuguese grapevine varieties and climate change adaptation. Int. J. Climatol. 2016, 36, 1–12. [Google Scholar] [CrossRef]
- Fraga, H.; García de Cortázar Atauri, I.; Malheiro, A.C.; Santos, J.A. Modelling climate change impacts on viticultural yield, phenology and stress conditions in Europe. Glob. Chang. Biol. 2016, 22, 3774–3788. [Google Scholar] [CrossRef] [PubMed]
- Fraga, H.; Malheiro, A.C.; Moutinho-Pereira, J.; Santos, J.A. Future scenarios for viticultural zoning in Europe: Ensemble projections and uncertainties. Int. J. Biometeorol. 2013, 57, 909–925. [Google Scholar] [CrossRef] [PubMed]
- Permanhani, M.; Costa, J.M.; Conceição, M.A.F.; de Souza, R.T.; Vasconcellos, M.A.S.; Chaves, M.M. Deficit irrigation in table grape: Eco-physiological basis and potential use to save water and improve quality. Theor. Exp. Plant Physiol. 2016, 28, 85–108. [Google Scholar] [CrossRef]
- Pinillos, V.; Chiamolera, F.M.; Ortiz, J.F.; Hueso, J.J.; Cuevas, J. Post-veraison regulated deficit irrigation in “Crimson Seedless” tablegrape saves water and improves berry skin color. Agric. Water Manag. 2015, 165, 181–189. [Google Scholar] [CrossRef]
- Conesa, M.R.R.; de la Rosa, J.M.M.; Domingo, R.; Bañon, S.; Pérez-Pastor, A. Changes induced by water stress on water relations, stomatal behaviour and morphology of table grapes (cv. Crimson Seedless) grown in pots. Sci. Hortic. 2016, 202, 9–16. [Google Scholar] [CrossRef]
- Williams, L.E. Effects of applied water amounts at various fractions of evapotranspiration (ETc) on leaf gas exchange of Thompson Seedless grapevines. Aust. J. Grape Wine Res. 2012, 18, 100–108. [Google Scholar] [CrossRef]
- Tomás, M.; Medrano, H.; Pou, A.; Escalona, J.M.; Martorell, S.; Ribas-Carbó, M.; Flexas, J. Water-use efficiency in grapevine cultivars grown under controlled conditions: Effects of water stress at the leaf and whole-plant level. Aust. J. Grape Wine Res. 2012, 18, 164–172. [Google Scholar] [CrossRef]
- Bota, J.; Tomás, M.; Flexas, J.; Medrano, H.; Escalona, J.M. Differences among grapevine cultivars in their stomatal behavior and water use efficiency under progressive water stress. Agric. Water Manag. 2016, 164, 91–99. [Google Scholar] [CrossRef]
- Bota, J.; Flexas, J.; Medrano, H. Genetic variability of photosynthesis and water use in Balearic grapevine cultivars. Ann. Appl. Biol. 2001, 138, 353–361. [Google Scholar] [CrossRef]
- Tortosa, I.; Escalona, J.M.; Bota, J.; Tomas, M.; Hernandez, E.; Escudero, E.G.; Medrano, H. Exploring the genetic variability in water use efficiency: Evaluation of inter and intra cultivar genetic diversity in grapevines. Plant Sci. 2016, 251, 35–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomás, M.; Medrano, H.; Escalona, J.M.; Martorell, S.; Pou, A.; Ribas-Carbó, M.; Flexas, J. Variability of water use efficiency in grapevines. Environ. Exp. Bot. 2014, 103, 148–157. [Google Scholar] [CrossRef]
- Schulze, E. Carbon dioxide and water vapor exchane in repsponse to drought in the atmosphere and in the soil. Annu. Rev. Plant Physiol. 1986, 37, 247–274. [Google Scholar] [CrossRef]
- Chaves, M.M. Effects of water deficits on carbon assimilation. J. Exp. Bot. 1991, 42, 1–16. [Google Scholar] [CrossRef]
- Toumi, I.; M’Sehli, W.; Bourgou, S.; Jallouli, N.; Bensalem-Fnayou, A.; Ghorbel, A.; Mliki, A. Response of ungrafted and grafted grapevine cultivars and rootstocks (Vitis sp.) to water stress. J. Int. Sci. Vigne Vin. 2007, 41, 85–93. [Google Scholar] [CrossRef]
- Gómez-del-Campo, M.; Ruiz, C.; Baeza, P.; Lissarrague, J.R. Drought adaptation strategies of four grapevine cultivars (Vitis vinifera L.): Modification of the properties of the leaf area. J. Int. Sci. Vigne Vin. 2003, 37, 131–143. [Google Scholar] [CrossRef]
- Lavoie-Lamoureux, A.; Sacco, D.; Risse, P.-A.; Lovisolo, C. Factors influencing stomatal conductance in response to water availability in grapevine: A meta-analysis. Physiol. Plant. 2017. [Google Scholar] [CrossRef]
- Chaves, M.M.; Oliveira, M.M. Mechanisms underlying plant resilience to water deficits: Prospects for water-saving agriculture. J. Exp. Bot. 2004, 55, 2365–2384. [Google Scholar] [CrossRef]
- Morison, J.I.L.; Baker, N.R.; Mullineaux, P.M.; Davies, W.J. Improving water use in crop production. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 639–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medrano, H.; Tomás, M.; Martorell, S.; Flexas, J.; Hernández, E.; Rosselló, J.; Pou, A.; Escalona, J.-M.; Bota, J. From leaf to whole-plant water use efficiency (WUE) in complex canopies: Limitations of leaf WUE as a selection target. Crop J. 2015, 3, 220–228. [Google Scholar] [CrossRef] [Green Version]
- Merli, M.C.; Gatti, M.; Galbignani, M.; Bernizzoni, F.; Magnanini, E.; Poni, S. Water use efficiency in Sangiovese grapes (Vitis vinifera L.) subjected to water stress before veraison: Different levels of assessment lead to different conclusions. Funct. Plant Biol. 2015, 42, 198–208. [Google Scholar] [CrossRef]
- Scholander, P.F.; Hammel, H.T.; Bradstreet, E.D.; Hemmingsen, E.A. Sap Pressure in Vascular Plants: Negative hydrostatic pressure can be measured in plants. Science 1965, 148, 339–346. [Google Scholar] [CrossRef]
- Piepho, H.P. A SAS macro for generating letter displays of pairwise mean comparisons. Commun. Biometry Crop Sci. 2012, 7, 4–13. [Google Scholar]
- Ojeda, H.; Deloire, A.; Carbonneau, A. Influence of water deficits on grape berry growth. Vitis 2001, 40, 141–145. [Google Scholar]
- Costa, J.M.; Ortuño, M.F.; Lopes, C.M.; Chaves, M.M. Grapevine varieties exhibiting differences in stomatal response to water deficit. Funct. Plant Biol. 2012, 39, 179–189. [Google Scholar] [CrossRef]
- Ezzahouani, A.; Williams, L.E. Effect of irrigation amount and preharvest cutoff date on vine water status and productivity of Danlas grapevines. Am. J. Enol. Vitic. 2007, 58, 333–340. [Google Scholar]
- Wenter, A.; Zanotelli, D.; Montagnani, L.; Tagliavini, M.; Andreotti, C. Effects of an early-summer drought stress on leaf photosynthesis, growth and yields of grapevine in mountain conditions. Acta Hortic. 2017, 457–462. [Google Scholar] [CrossRef]
- Meggio, F.; Prinsi, B.; Negri, A.S.; Simone Di Lorenzo, G.; Lucchini, G.; Pitacco, A.; Failla, O.; Scienza, A.; Cocucci, M.; Espen, L. Biochemical and physiological responses of two grapevine rootstock genotypes to drought and salt treatments. Aust. J. Grape Wine Res. 2014, 20, 310–323. [Google Scholar] [CrossRef]
- Pou, A.; Flexas, J.; Alsina, M.D.M.; Bota, J.; Carambula, C.; De Herralde, F.; Galmés, J.; Lovisolo, C.; Jiménez, M.; Ribas-Carbó, M.; et al. Adjustments of water use efficiency by stomatal regulation during drought and recovery in the drought-adapted Vitis hybrid Richter-110 (V. berlandieri x V. rupestris). Physiol. Plant. 2008, 134, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Medrano, H.; Escalona, J.M.; Bota, J.; Gulías, J.; Flexas, J. Regulation of photosynthesis of C3 plants in response to progressive drought: Stomatal conductance as a reference parameter. Ann. Bot. 2002, 89, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Flexas, J.; Bota, J.; Escalona, J.-M.; Sampol, B.; Medrano, H. Effects of drought on photosynthesis in grapevines under field conditions: An evaluation of stomatal and mesophyll limitations. Funct. Plant Biol. 2002, 29, 461–471. [Google Scholar] [CrossRef]
- Chaves, M.M.; Zarrouk, O.; Francisco, R.; Costa, J.M.; Santos, T.; Regalado, A.P.; Rodrigues, M.L.; Lopes, C.M. Grapevine under deficit irrigation: Hints from physiological and molecular data. Ann. Bot. 2010, 105, 661–676. [Google Scholar] [CrossRef] [PubMed]
- Jara-Rojas, F.; Ortega-Farías, S.; Valdés-Gómez, H.; Acevedo-Opazo, C. Gas exchange relations of ungrafted grapevines (cv. Carménère) growing under irrigated field conditions. S. Afr. J. Enol. Vitic. 2015, 36, 231–242. [Google Scholar] [CrossRef]
- Flexas, J.; Medrano, H. Drought-inhibition of photosynthesis in C3 plants: Stomatal and non-stomatal limitations revisited. Ann. Bot. 2002, 89, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Ghaderi, N.; Talai, A.R.; Ebadi, A.; Lessani, H. The physiological response of three Iranian grape cultivars to progressive drought stress. J. Agric. Sci. Technol. 2011, 13, 601–610. [Google Scholar]
- Romero, P.; Fernández-Fernández, J.I.; Martinez-Cutillas, A. Physiological Thresholds for Efficient Regulated Deficit Irrigation Management in Winegrapes Under Semiarid Conditions: Soil-Plant-Water Relationships and Berry Composition. Acta Hortic. 2012, 171–178. [Google Scholar] [CrossRef]
- Tramontini, S.; Döring, J.; Vitali, M.; Ferrandino, A.; Stoll, M.; Lovisolo, C. Soil water-holding capacity mediates hydraulic and hormonal signals of near-isohydric and near-anisohydric Vitis cultivars in potted grapevines. Funct. Plant Biol. 2014, 41, 1119–1128. [Google Scholar] [CrossRef] [Green Version]
- Schultz, H.R. Differences in hydraulic architecture account for near-isohydric and anisohydric behaviour of two eld-grown. Plant Cell Environ. 2003, 26, 1393–1406. [Google Scholar] [CrossRef]
- Lovisolo, C.; Perrone, I.; Hartung, W.; Schubert, A. An abscisic acid-related reduced transpiration promotes gradual embolism repair when grapevines are rehydrated after drought. New Phytol. 2008, 180, 642–651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palliotti, A.; Tombesi, S.; Frioni, T.; Famiani, F.; Silvestroni, O.; Zamboni, M.; Poni, S. Morpho-structural and physiological response of container-grown Sangiovese and Montepulciano cvv. (Vitis vinifera) to re-watering after a pre-veraison limiting water deficit. Funct. Plant Biol. 2014, 41, 634–647. [Google Scholar] [CrossRef]
- Pou, A.; Medrano, H.; Tomàs, M.; Martorell, S.; Ribas-Carbó, M.; Flexas, J. Anisohydric behaviour in grapevines results in better performance under moderate water stress and recovery than isohydric behaviour. Plant Soil 2012, 359, 335–349. [Google Scholar] [CrossRef]
- Maroco, J.P.; Rodrigues, M.L.; Lopes, C.; Chaves, M.M. Limitations to leaf photosynthesis in field-grown grapevine under drought—Metabolic and modelling approaches. Funct. Plant Biol. 2002, 29, 451–459. [Google Scholar] [CrossRef]
- Gibberd, M.R.; Walker, R.R.; Blackmore, D.H.; Condon, A.G. Transpiration efficiency and carbon-isotope discrimination of grapevines grown under well-watered conditions in either glasshouse or vineyard. Aust. J. Grape Wine Res. 2001, 7, 110–117. [Google Scholar] [CrossRef]
- Tardieu, F.; Granier, C.; Muller, B. Modelling Leaf Expansion in a Fluctuating Environment: Are Changes in Specific Leaf Area a Consequence of Changes in Expansion Rate? New Phytol. 1999, 143, 33–43. [Google Scholar] [CrossRef]
- De Pinheiro Henriques, A.R.; Marcelis, L.F.M. Regulation of growth at steady-state nitrogen nutrition in lettuce (Lactuca sativa L.): Interactive effects of nitrogen and irradiance. Ann. Bot. 2000, 86, 1073–1080. [Google Scholar] [CrossRef]
- Koundouras, S.; Tsialtas, I.T.; Zioziou, E.; Nikolaou, N. Rootstock effects on the adaptive strategies of grapevine (Vitis vinifera L. cv. Cabernet-Sauvignon) under contrasting water status: Leaf physiological and structural responses. Agric. Ecosyst. Environ. 2008, 128, 86–96. [Google Scholar] [CrossRef]
- Peterson, R.B.; Zelitch, I. Relationship between Net CO2 Assimilation and Dry Weight Accumulation in Field-Grown Tobacco. Plant Physiol. 1982, 70, 677–685. [Google Scholar] [CrossRef]
- Gómez-del-Campo, M.; Ruiz, C.; Sotés, V.; Lissarrague, J.R. Consequences of Water Consumption in the Leaf Area and Dry Matter Partitioning in Four Grapevine Varieties. Acta Hortic. 1996, 331–338. [Google Scholar] [CrossRef]
- Gómez-del-Campo, M.; Baeza, P.; Ruiz, C.; Lissarrague, J.R. Effects of water stress on dry matter content and partitioning in four grapevine cultivars (Vitis vinifera L.). J. Int. Sci. Vigne Vin 2005, 39, 1–10. [Google Scholar] [CrossRef]
- Mullins, M.G.; Bouquet, A.; Williams, L.E. Biology of the Grapevine; Cambridge University Press: Cambridge, UK, 1992. [Google Scholar]
- Williams, L.E. Grape. In Photoassimilate Distribution in Plants and Crops Source-Sink Relationship, 1st ed.; Zamski, E., Ed.; CRC Press: New York, NY, USA, 1996; pp. 851–881. ISBN 9780824794408. [Google Scholar]



{kind=link}
{kind=link}
| Water Deficit Treatment | Daily Irrigation to | ||||
|---|---|---|---|---|---|
| Control | 75% AWC | ||||
| Moderate | 50% AWC | ||||
| Severe | 25% AWC | ||||
| Year | Timeframe | Weeks of Water Deficit | BBCH (at the Beginning of Water Deficit) | ||
| 2014 | 22.9. | – | 29.10. | 5.5 | 19 |
| 2015 | 12.5. | – | 21.7. | 10 | 16–18 |
| 2016 | 15.6. | – | 16.9. | 12 | 71 |
| Cultivar | Treatment | An | E | gs | Ci | WUEinst | WUEi | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (µmol m−2s−1) | (mmol m−2s−1) | (mmol m−2s−1) | (µmol m−2s−1) | A/E | A/gs | |||||||||||||||
| Muscat Bleu | Control | 4.65 | a | C | 0.96 | a | C | 39.60 | a | C | 218.12 | a | ABC | 3.98 | b | ABC | 0.10 | b | ABC | |
| Moderate | 5.57 | a | 1.07 | b | 43.84 | b | 203.69 | a | 4.76 | b | 0.12 | b | ||||||||
| Severe | 5.32 | b | 0.69 | c | 27.19 | c | 120.74 | b | 6.57 | a | 0.17 | a | ||||||||
| Fanny | Control | 12.30 | a | A | 3.28 | a | A | 148.08 | a | A | 247.20 | a | ABC | 3.74 | b | ABC | 0.08 | b | ABC | |
| Moderate | 11.86 | a | 2.45 | b | 104.30 | b | 213.00 | a | 4.23 | b | 0.10 | b | ||||||||
| Severe | 8.71 | b | 1.32 | c | 53.11 | c | 149.88 | b | 6.84 | a | 0.17 | a | ||||||||
| Nero | Control | 10.67 | a | B | 3.14 | a | A | 135.63 | a | A | 258.41 | a | A | 3.25 | b | C | 0.08 | b | C | |
| Moderate | 7.54 | a | 1.93 | b | 81.64 | b | 241.87 | a | 3.67 | b | 0.09 | b | ||||||||
| Severe | 7.14 | b | 1.77 | c | 75.26 | c | 236.15 | b | 3.69 | a | 0.09 | a | ||||||||
| Palatina | Control | 11.03 | a | AB | 1.80 | a | B | 74.22 | a | B | 155.00 | a | BC | 5.83 | b | AB | 0.14 | b | AB | |
| Moderate | 9.89 | a | 1.44 | b | 59.32 | b | 149.40 | a | 6.72 | b | 0.17 | b | ||||||||
| Severe | 7.80 | b | 1.03 | c | 41.17 | c | 127.33 | b | 6.75 | a | 0.17 | a | ||||||||
| Thompson Seedless | Control | 6.64 | a | C | 1.29 | a | C | 51.29 | a | C | 175.67 | a | C | 5.15 | b | A | 0.13 | b | A | |
| Moderate | 5.88 | a | 1.14 | b | 46.38 | b | 191.45 | a | 4.87 | b | 0.12 | b | ||||||||
| Severe | 4.81 | b | 0.71 | c | 27.05 | c | 117.62 | b | 7.11 | a | 0.19 | a | ||||||||
| Crimson Seedless | Control | 5.17 | a | C | 1.24 | a | C | 49.66 | a | C | 225.53 | a | AB | 4.00 | b | BC | 0.10 | b | BC | |
| Moderate | 5.19 | a | 1.25 | b | 48.36 | b | 220.30 | a | 4.05 | b | 0.10 | b | ||||||||
| Severe | 2.12 | b | 0.44 | c | 15.98 | c | 192.44 | b | 4.51 | a | 0.12 | a | ||||||||
| ANOVA | ||||||||||||||||||||
| Cultivar (C) | <0.0001 | <0.0001 | <0.0001 | 0.0361 | 0.0355 | 0.0324 | ||||||||||||||
| Treatment (T) | 0.0006 | <0.0001 | <0.0001 | <0.0001 | 0.0001 | <0.0001 | ||||||||||||||
| C*T | 0.5774 | 0.4113 | 0.3597 | 0.5151 | 0.4336 | 0.3549 | ||||||||||||||
| Cultivar | Treatment | LA | SLA | Total DM | Number of Leaves | WUEDM | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (cm2) | (cm2 g−1) | (g) | (g L−1) | ||||||||||||
| Muscat Bleu | Control | 1074.27 | C | 171.72 | a | AB | 32.54 | a | CD | 9.88 | D | 0.04 | c | B | |
| Moderate | 1151.48 | 152.17 | b | C | 35.95 | a | 10.00 | 0.05 | b | ||||||
| Severe | 1122.71 | 167.16 | a | AB | 36.08 | b | 9.38 | 0.06 | a | ||||||
| Fanny | Control | 1191.18 | C | 168.62 | a | B | 60.33 | a | A | 12.75 | C | 0.07 | c | A | |
| Moderate | 1228.93 | 171.35 | a | A | 70.98 | a | 11.63 | 0.09 | b | ||||||
| Severe | 1087.03 | 175.67 | a | A | 53.89 | b | 11.63 | 0.09 | a | ||||||
| Nero | Control | 1282.80 | C | 152.30 | ab | C | 43.45 | a | B | 16.38 | B | 0.05 | c | B | |
| Moderate | 1229.79 | 144.03 | b | C | 41.43 | a | 16.25 | 0.05 | b | ||||||
| Severe | 1130.14 | 159.73 | a | BC | 35.79 | b | 16.25 | 0.06 | a | ||||||
| Palatina | Control | 1311.60 | C | 152.09 | a | C | 34.06 | a | BC | 16.25 | B | 0.04 | c | B | |
| Moderate | 1206.16 | 153.33 | a | BC | 37.52 | a | 15.25 | 0.05 | b | ||||||
| Severe | 1121.81 | 149.95 | a | C | 33.65 | b | 15.25 | 0.05 | a | ||||||
| Thompson Seedless | Control | 1766.40 | B | 169.15 | a | B | 32.31 | a | E | 14.71 | B | 0.03 | c | C | |
| Moderate | 1560.87 | 164.10 | a | AB | 29.46 | a | 14.88 | 0.03 | b | ||||||
| Severe | 1658.88 | 161.83 | a | B | 29.37 | b | 15.13 | 0.05 | a | ||||||
| Crimson Seedless | Control | 2185.06 | A | 180.90 | a | A | 34.90 | a | DE | 21.13 | A | 0.03 | c | C | |
| Moderate | 1836.10 | 174.25 | ab | A | 31.77 | a | 19.25 | 0.03 | b | ||||||
| Severe | 1767.82 | 165.01 | b | AB | 25.47 | b | 21.25 | 0.04 | a | ||||||
| ANOVA | |||||||||||||||
| Cultivar (C) | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | ||||||||||
| Treatment (T) | 0.0633 | 0.0448 | 0.0324 | 0.8144 | <0.0001 | ||||||||||
| C*T | 0.7734 | 0.0069 | 0.4589 | 0.9975 | 0.7317 | ||||||||||
| Cultivar | Treatment | Leaf DM (g) | Stem DM (g) | Petioles DM (g) | Fruit DM (g) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Muscat Bleu | Control | 6.33 | D | 17.99 | a | AB | 1.05 | a | D | 21.98 | B | ||
| Moderate | 7.66 | 18.06 | a | AB | 1.04 | a | 14.81 | ||||||
| Severe | 6.76 | 16.06 | a | A | 1.03 | b | 11.58 | ||||||
| Fanny | Control | 7.14 | D | 13.46 | a | C | 1.14 | a | D | 35.48 | A | ||
| Moderate | 7.27 | 12.51 | a | C | 1.15 | a | 57.62 | ||||||
| Severe | 6.27 | 11.36 | a | C | 0.93 | b | 37.45 | ||||||
| Nero | Control | 8.55 | C | 16.83 | a | B | 1.33 | a | C | 27.08 | B | ||
| Moderate | 9.12 | 16.31 | a | B | 1.38 | a | 17.80 | ||||||
| Severe | 7.27 | 14.98 | a | AB | 1.09 | b | 14.62 | ||||||
| Palatina | Control | 8.72 | C | 17.49 | a | B | 1.63 | a | B | 17.55 | B | ||
| Moderate | 7.98 | 17.80 | a | AB | 1.47 | a | 23.13 | ||||||
| Severe | 7.69 | 15.20 | b | AB | 1.33 | b | 16.74 | ||||||
| Thompson Seedless | Control | 10.47 | B | 17.47 | a | B | 1.61 | a | B | 18.51 | B | ||
| Moderate | 10.01 | 17.27 | a | AB | 1.49 | a | 10.11 | ||||||
| Severe | 10.39 | 16.54 | a | A | 1.46 | b | 14.03 | ||||||
| CrimsonSeedless | Control | 12.11 | A | 20.03 | a | A | 2.07 | a | A | n.a. | n.a. | ||
| Moderate | 11.07 | 18.96 | a | A | 1.85 | a | n.a. | ||||||
| Severe | 10.90 | 13.06 | b | BC | 1.62 | b | n.a. | ||||||
| ANOVA | |||||||||||||
| Cultivar (C) | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |||||||||
| Treatment (T) | 0.1221 | <0.0001 | 0.0003 | 0.5901 | |||||||||
| C*T | 0.6549 | 0.0191 | 0.6271 | 0.4693 | |||||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weiler, C.S.; Merkt, N.; Hartung, J.; Graeff-Hönninger, S. Variability among Young Table Grape Cultivars in Response to Water Deficit and Water Use Efficiency. Agronomy 2019, 9, 135. https://doi.org/10.3390/agronomy9030135
Weiler CS, Merkt N, Hartung J, Graeff-Hönninger S. Variability among Young Table Grape Cultivars in Response to Water Deficit and Water Use Efficiency. Agronomy. 2019; 9(3):135. https://doi.org/10.3390/agronomy9030135
Chicago/Turabian StyleWeiler, Carolin Susanne, Nikolaus Merkt, Jens Hartung, and Simone Graeff-Hönninger. 2019. "Variability among Young Table Grape Cultivars in Response to Water Deficit and Water Use Efficiency" Agronomy 9, no. 3: 135. https://doi.org/10.3390/agronomy9030135
APA StyleWeiler, C. S., Merkt, N., Hartung, J., & Graeff-Hönninger, S. (2019). Variability among Young Table Grape Cultivars in Response to Water Deficit and Water Use Efficiency. Agronomy, 9(3), 135. https://doi.org/10.3390/agronomy9030135