Solanum aethiopicum gr. gilo and Its Interspecific Hybrid with S. melongena as Alternative Rootstocks for Eggplant: Effects on Vigor, Yield, and Fruit Physicochemical Properties of Cultivar ′Scarlatti′

 ,
, 
 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Seedlings Production and Grafting
2.3. Cultivation Conditions
2.4. Grafting Success, Biometric Parameters, Yield, and Apparent Fruit Quality Evaluation
2.5. Pulp Browning, Soluble Solid Content, and Chemicals
2.6. Statistical Analysis
3. Results
3.1. Grafting Success and Plant Biometric Parameters
3.2. Yield
3.3. Apparent Fruit Quality
3.4. Intrinsic and Extrinsic Fruit Quality
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. Production. Available online: www.fao.org (accessed on 16 January 2019).
- Kaushik, P.; Prohens, J.; Vilanova, S.; Gramazio, P.; Plazas, M. Phenotyping of eggplant wild relatives and interspecific hybrids with conventional and phenomics descriptors provides insight for their potential utilization in breeding. Front. Plant Sci. 2016, 7, 677. [Google Scholar] [CrossRef]
- D’Anna, F.; Sabatino, L. Morphological and agronomical characterization of eggplant genetic resources from the Sicily area. J. Food Agri. Environ. 2013, 11, 401–404. [Google Scholar]
- Bletsos, F.; Thanassoulopoulos, C.; Roupakias, D. Effect of grafting on growth, yield and Verticillium wilt of eggplant. HortScience 2003, 38, 183–186. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y.; Jawad, R.; Kumar, P.; Rea, E.; Cardarelli, M. The effectiveness of grafting to improve NaCl and CaCl2 tolerance in cucumber. Sci. Hortic. 2013, 164, 380–391. [Google Scholar] [CrossRef]
- Kumar, P.; Rouphael, Y.; Cardarelli, M.; Colla, G. Vegetable grafting as a tool to improve drought resistance and water use efficiency. Front. Plant Sci. 2017, 8, 1130. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Lucini, L.; Rouphael, Y.; Cardarelli, M.; Kalunke, R.M.; Colla, G. Insight into the role of grafting and arbuscular mycorrhiza on cadmium stress tolerance in tomato. Front. Plant Sci. 2015, 6, 477. [Google Scholar] [CrossRef]
- Ginoux, G.; Laterrot, H. Greffage de l’aubergine. Proceedings reflexions du portegreffe. PHM Revue Hortic. 1991, 321, 49–54. [Google Scholar]
- Gisbert, C.; Prohens, J.; Raigón, M.D.; Stommel, J.R.; Nuez, F. Eggplant relatives as sources of variation for developing new rootstocks: Effects of grafting on eggplant yield and fruit apparent quality and composition. Sci. Hortic. 2011, 128, 14–22. [Google Scholar] [CrossRef]
- Sabatino, L.; Iapichino, G.; D’Anna, F.; Palazzolo, E.; Mennella, G.; Rotino, G.L. Hybrids and allied species as potential rootstocks for eggplant: Effect of grafting on vigour, yield and overall fruit quality traits. Sci. Hortic. 2018, 228, 81–90. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; Rouphael, Y.; Colla, G.; Zrenner, R.; Schwarz, D. Vegetable grafting: The implications of a growing agronomic imperative for vegetable fruit quality and nutritive value. Front. Plant Sci. 2017, 8, 741. [Google Scholar] [CrossRef]
- Cappelli, C.; Stravato, V.M.; Rotino, G.L.; Buonaurio, R. Sources of resistance among Solanum spp. to an Italian isolate of Fusarium oxysporum f sp. melongenae. In Proceedings of the 9th EUCARPIA Meeting Genetic and Breeding of Capsicum and Eggplant, Budapest, Hungary, 21–25 August 1995; pp. 221–224. [Google Scholar]
- Rizza, F.; Mennella, G.; Collonnier, C.; Shiachakr, D.; Kashyap, V.; Rajam, M.V.; Prestera, M.; Rotino, G.L. Androgenic dihaploids from somatic hybrids between Solanum melongena and S. aethiopicum group gilo as a source of resistance to Fusarium oxysporum f. sp. melongenae. Plant Cell Rep. 2002, 20, 1022–1032. [Google Scholar]
- Hébert, Y. Comparative resistance of nine species of the genes Solanum to bacterial wilt Psedomonas solanacearum and the nematode Meloidogyne incognita. Implications for the breeding of aubergine (S. melongena) in the humid tropical zone. Agronomie 1985, 5, 27–32. [Google Scholar] [CrossRef]
- Prohens, J.; Plazas, M.; Raigón, M.D.; Seguí-Simarro, J.M.; Stommel, J.R.; Vilanova, S. Characterization of interspecific hybrids and backcross generations from crosses between two cultivated eggplants (Solanum melongena and S. aethiopicum Kumba group) and implications for eggplant breeding. Euphytica 2012, 186, 517–538. [Google Scholar] [CrossRef]
- Sawadogo, B.; Bationo-Kando, P.; Sawadogo, N.; Kiebre, Z.; Kiebre, M.; Nanema, K.R.; Traore, R.E.; Sawadogo, M.; Zongo, J.D. Variation, correlation and heritability of interest characters for selection of African eggplant. Afr. Crop Sci. J. 2016, 24, 213–221. [Google Scholar] [CrossRef]
- Toppino, L.; Mennella, G.; Rizza, F.; D’alessandro, A.; Sihachakr, D.; Rotino, G.L. ISSR and isozyme characterization of androgenetic dihaploids reveals tetrasomic inheritance in tetraploid somatic hybrids between Solanum melongena and Solanum aethiopicum group gilo. J. Hered. 2008, 99, 304–315. [Google Scholar] [CrossRef] [PubMed]
- Toppino, L.; Vale, G.; Rotino, G.L. Inheritance of fusarium wilt resistance introgressed from Solanum aethiopicum Gilo and Aculeatum groups into cultivated eggplant (S. melongena) and development of associated PCR-based markers. Mol. Breed. 2008, 22, 237–250. [Google Scholar] [CrossRef]
- Mennella, G.; Rotino, G.L.; Fibiani, M.; D’Alessandro, A.; Francese, G.; Toppino, L.; Cavallanti, F.; Acciarri, N.; Lo Scalzo, R. Characterization of health-related compounds in eggplant (Solanum melongena L.) lines derived from introgression of allied species. J. Agric. Food Chem. 2010, 58, 7597–7603. [Google Scholar] [CrossRef]
- Lee, J.M. Cultivation of grafted vegetables I: Current status, grafting methods and benefits. Hort. Sci. 1994, 29, 235–239. [Google Scholar] [CrossRef]
- Miceli, A.; Sabatino, L.; Moncada, A.; Vetrano, F.; D’Anna, F. Nursery and field evaluation of eggplant grafted onto unrooted cuttings of Solanum torvum Sw. Sci. Hortic. 2014, 178, 203–210. [Google Scholar] [CrossRef]
- Baixauli, C. Berenjena. In La horticultura espãnola; Nuez, F., Llácer, G., Eds.; Ediciones de Horticultura: Reus, Spain, 2001. [Google Scholar]
- Prohens, J.; Blanca, J.M.; Nuez, F. Morphological and molecular variation in a collection of eggplant from a secondary center of diversity: Implications for conservation and breeding. J. Am. Soc. Hortic. Sci. 2005, 130, 54–63. [Google Scholar] [CrossRef]
- Larrigaudiere, C.; Lentheric, I.; Vendrell, M. Relationship between enzymatic browning and internal disorders in controlled atmosphere stored pears. J. Sci. Food Agric. 1998, 78, 232–236. [Google Scholar] [CrossRef]
- Concellòn, A.; Añón, M.; Chaves, A.R. Effect of low temperature storage on physical and physiological characteristics of eggplant fruit (Solanum melongena L.). LWT 2007, 40, 389–396. [Google Scholar] [CrossRef]
- Sabatino, L.; Iapichino, G.; Maggio, A.; D’Anna, E.; Bruno, M.; D’Anna, F. Grafting affects yield and phenolic profile of Solanum melongena L. landraces. J. Integr. Agric. 2016, 15, 1017–1024. [Google Scholar] [CrossRef]
- Fogg, D.N.; Wilkinson, N.T. The colorimetric determination of phosphorus. Analist 1958, 83, 406–414. [Google Scholar] [CrossRef]
- Morand, P.; Gullo, J.L. Mineralisation des tissus vegetaux en vue du dosage de P, Ca, Mg, Na, K. Ann. Agron. 1970, 21, 229–236. [Google Scholar]
- Stommel, J.R.; Whitaker, B.D. Phenolic acid content and composition of eggplant fruit in a germplasm core subset. J. Am. Soc. Hortic. Sci. 2003, 128, 704–710. [Google Scholar] [CrossRef]
- Mennella, G.; Lo Scalzo, R.; Fibiani, M.; D’Alessandro, A.; Francese, G.; Toppino, L.; Acciarri, N.; De Almeida, A.E.; Rotino, G.L. Chemical and bioactive quality traits during fruit ripening in eggplant (S. melongena L.) and allied species. J. Agric. Food Chem. 2012, 60, 11821–11831. [Google Scholar] [CrossRef]
- Lo Scalzo, R.; Fibiani, M.; Mennella, G.; Rotino, G.L.; Dal Sasso, M.; Culici, M.; Spallino, A.; Braga, P.C. Thermal treatments of eggplant (Solanum melongena L.) increases the antioxidant content and the inhibitory effect on human neutrophil burst. J. Agric. Food Chem. 2010, 58, 3371–3379. [Google Scholar] [CrossRef]
- Birner, J. A method for the determination of total steroid bases. J. Pharm. Sci. 1969, 58, 258–259. [Google Scholar] [CrossRef]
- Kuronen, P.; Väänänen, T.; Pehu, E. Reversed-phase liquid chromatographic separation and simultaneous profiling of steroidal glycoalkaloids and their aglycones. J. Chromatogr. A. 1999, 863, 25–35. [Google Scholar] [CrossRef]
- Sabatino, L.; Palazzolo, E.; D’Anna, F. Grafting suitability of Sicilian eggplant ecotypes onto Solanum torvum: Fruit composition, production and phenology. J. Food Agric. Environ. 2013, 11, 1195–1200. [Google Scholar]
- Davis, A.R.; Perkins-Veazie, P.; Hassell, R.; Levi, A.; King, S.R.; Zhang, X.P. Grafting effects on vegetable quality. HortScience 2008, 43, 1670–1672. [Google Scholar] [CrossRef]
- Rouphael, Y.; Kyriacou, M.; Colla, G. Vegetable grafting: A toolbox for securing yield stability under multiple stress conditions. Front. Plant Sci. 2018, 8, 2255. [Google Scholar] [CrossRef]
- Kawaguchi, M.; Taji, A.; Backhouse, D.; Oda, M. Anatomy and physiology of graft incompatibility in solanaceous plants. J. Hortic. Sci. Biotechnol. 2008, 83, 581–588. [Google Scholar] [CrossRef]
- Miguel, A.; de la Torre, F.; Baixauli, C.; Maroto, J.V.; Jordá, M.C.; López, M.M.; García- Jiménez, J. Elinjerto de hortalizas; Ministerio de Agricultura, Pesca y Alimentación: Madrid, Spain, 2007.
- Rahman, M.A.; Rashid, M.A.; Hossain, M.M.; Salam, M.A.; Masum, A.S.M.H. Grafting compatibility of cultivated eggplant varieties with wild Solanum species. Pak. J. Biol. Sci. 2002, 5, 755–757. [Google Scholar]
- Cohen, R.; Horev, C.; Burger, Y.; Shriber, S.; Hershenhorn, J.; Katanand, J.; Edelstein, M. Horticultural and pathological aspects of Fusarium wilt management using grafted melons. Hort. Sci. 2002, 37, 1069–1073. [Google Scholar] [CrossRef]
- Fita, A.; Picó, B.; Roig, C.; Nuez, F. Performance of Cucumis melo spp. agrestis as a rootstock for melon. J. Hortic. Sci. Biotechnol. 2004, 82, 184–190. [Google Scholar]
- Maršič, N.K.; Mikulič-Petkovšek, M.; Štampar, F. Grafting influences phenolic profile and carpometric traits of fruits of greenhouse-grown eggplant (Solanum melongena L.). J. Agric. Food Chem. 2014, 62, 10504–10514. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Falcón, J.E.; Prohens, J.; Rodríguez-Burruezo, A.; Nuez, F. Potential of local varieties and their hybrids for the improvement of eggplant production in the open field and greenhouse cultivation. J. Food Agric. Environ. 2008, 6, 83–88. [Google Scholar]
- Portis, E.; Barchi, L.; Toppino, L.; Lanteri, S.; Acciarri, N.; Felicioni, N.; Fusari, F.; Barbierato, V.; Cericola, F.; Valè, G.; et al. QTL mapping in eggplant reveals clusters of yield-related loci and orthology with the tomato genome. PLoS ONE 2014, 9, e89499. [Google Scholar] [CrossRef] [PubMed]
- Moncada, A.; Miceli, A.; Vetrano, F.; Mineo, V.; Planeta, D.; D’Anna, F. Effect of grafting on yield and quality of eggplant (Solanum melongena L.). Sci. Hortic. 2013, 149, 108–114. [Google Scholar] [CrossRef]
- Mishra, B.B.; Gautam, S.; Sharma, A. Free phenolics and polyphenol oxidase (PPO): The factors affecting post-cut browning in eggplant (Solanum melongena). Food Chem. 2013, 139, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.P.; Luthria, D.; Wilson, T.; Vorsa, N.; Singh, V.; Banuelos, G.S.; Pasakdee, S. Polyphenols content and antioxidant capacity of eggplant pulp. Food Chem. 2009, 114, 955–961. [Google Scholar] [CrossRef]
- King, S.R.; Davis, A.R.; Zhang, X.; Crosby, K. Genetics, breeding and selection of rootstocks for Solanaceae and Cucurbitaceae. Sci. Hortic. 2010, 127, 106–111. [Google Scholar] [CrossRef]
- Prohens, J.; Rodriguez-Burruezo, A.; Raigon, M.D.; Nuez, F. Total phenolic concentration and browning susceptibility in a collection of different varietal types and hybrids of eggplant: Implications for breeding for higher nutritional quality and reduced browning. J. Am. Soc. Hortic. Sci. 2007, 132, 1–9. [Google Scholar] [CrossRef]
- Dixon, R.A.; Paiva, N.L. Stress-induced phenylpropanoid metabolism. Plant Cell 1995, 7, 1085–1097. [Google Scholar] [CrossRef] [PubMed]
- Moglia, A.; Lanteri, S.; Comino, C.; Acquadro, A.; de Vos, R.; Beekwilder, J. Stress induced biosynthesis of dicaffeoylquinic acids in globe artichoke. J. Agric. Food Chem. 2008, 56, 8641–8649. [Google Scholar] [CrossRef]
- Toppino, L.; Barchi, L.; Lo Scalzo, R.; Palazzolo, E.; Francese, G.; Fibiani, M.; D’Alessandro, A.; Papa, V.; Laudicina, V.A.; Sabatino, L.; et al. Mapping quantitative trait loci affectingbiochemical and morphologicalfruitproperties in eggplant (Solanum melongena L.). Front. Plant Sci. 2016, 7, 256. [Google Scholar] [CrossRef]
- Friedman, M.; McDonald, G.M. Potato glycoalkaloids: Chemistry, analyses, safety and plant physiology. Crit. Rev. Plant Sci. 1997, 16, 55–132. [Google Scholar] [CrossRef]
- Friedman, M. Tomato glycoalkaloids: Role in the plant and in the diet. J. Agric. Food Chem. 2002, 50, 5751–5780. [Google Scholar] [CrossRef]
- Jones, P.G.; Fenwick, G.R. The glycoalkaloid content of some edible solanaceous fruits and potato products. J. Sci. Food Agric. 2006, 32, 419–421. [Google Scholar] [CrossRef]
- Krits, P.; Fogelman, E.; Ginzberg, I. Potato steroidal glycoalkaloid levels and the expression of key isoprenoid metabolic genes. Planta 2007, 227, 143–150. [Google Scholar] [CrossRef]
- Khah, E.M.; Kakava, E.M.A.; Chachalis, D.; Goulas, C. Effect of grafting on growth and yield of tomato (Lycopersicon esculentum Mill.) in greenhouse and open field. J. Appl. Hortic. 2006, 8, 3–7. [Google Scholar]
- Gisbert, C.; Sánchez-Torres, P.; Raigón, M.D.; Nuez, F. Phytophthora capsici resistance evaluation in pepper hybrids: Agronomic performance and fruit quality of pepper grafted plants. J. Food Agric. Environ. 2010, 8, 116–121. [Google Scholar]


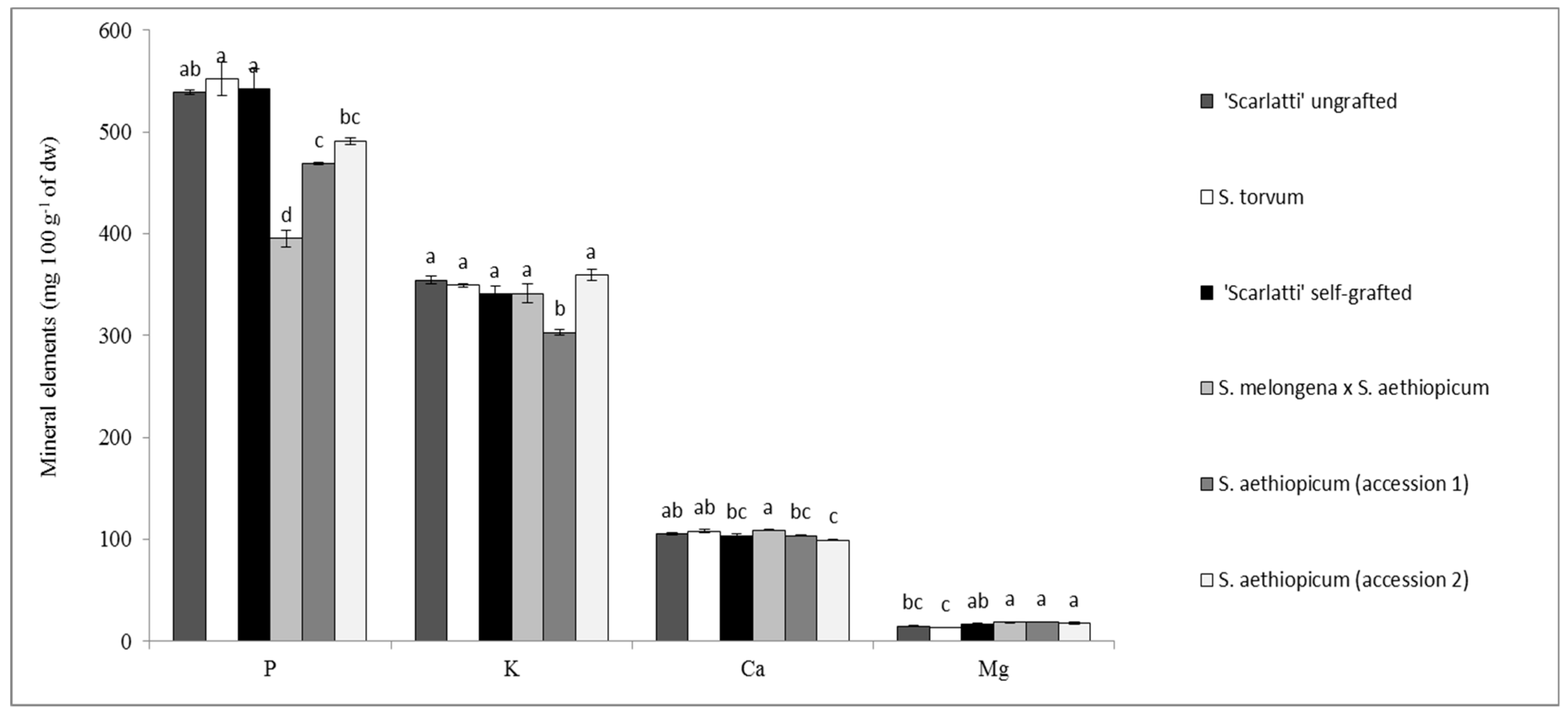{kind=link}
| Treatments | Grafting Success (%) | No. Leaves 50 DAT (No.) | Aboveground Biomass (kg) | First Flower Formation (DAT) |
|---|---|---|---|---|
| Rootstock | ||||
| ′Scarlatti′ ungrafted | - | 17.5 b | 4.5 ab | 53.1 a |
| S. torvum | 99.3 a | 24.3 ab | 5.1 a | 49.0 d |
| ′Scarlatti′ self-grafted | 99.6 a | 25.4 a | 4.9 a | 52.9 ab |
| Hybrid S. melongena × S. aethiopicum | 99.0 a | 21.1 ab | 4.6 ab | 49.3 d |
| S. aethiopicum (accession 1) | 93.6 b | 27.3 a | 3.7 c | 51.9 bc |
| S. aethiopicum (accession 2) | 89.1 b | 17.6 b | 4.0 bc | 51.5 c |
| Year | ||||
| 2014 | 96.5 a | 22.3 a | 4.4 a | 52.0 a |
| 2015 | 95.8 a | 22.1 a | 4.5 a | 50.6 b |
| Significance | ||||
| Rootstock | *** | ** | *** | *** |
| Year | NS | NS | NS | *** |
| Treatments | Total Yield Plant−1 (kg) | Marketable Yield Plant−1 (kg) | No. Marketable Fruits Plant−1 (No.) | Average Fruit Weight (g) |
|---|---|---|---|---|
| Rootstock | ||||
| ‘Scarlatti’ ungrafted | 3.4 ab | 3.3 ab | 14.5 a | 231.3 ns |
| S. torvum | 3.8 a | 3.6 a | 16.6 a | 216.8 ns |
| ‘Scarlatti’ self-grafted | 3.9 a | 3.6 a | 16.2 a | 223.5 ns |
| Hybrid S. melongena × S. aethiopicum | 3.4 ab | 3.2 ab | 15.1 a | 213.9 ns |
| S. aethiopicum (accession 1) | 2.7 c | 2.5 c | 9.1 b | 330.1 ns |
| S. aethiopicum (accession 2) | 3.1 bc | 2.9 bc | 14.1 a | 207.5 ns |
| Significance | ||||
| Rootstock | *** | ** | *** | NS |
| Treatments | H° | Fruit Dry Matter (%) | Firmness (N) | SSC (°Brix) |
|---|---|---|---|---|
| Rootstock | ||||
| Scarlatti’ ungrafted | 360.1 b | 7.3 ns | 60.0 ns | 4.5 b |
| S. torvum | 360.3 a | 7.2 ns | 43.2 ns | 3.9 c |
| Scarlatti’ self-grafted | 360.1 b | 6.8 ns | 44.2 ns | 4.1 c |
| Hybrid S. melongena × S. aethiopicum | 360.0 b | 6.8 ns | 49.0 ns | 4.6 ab |
| S. aethiopicum (accession 1) | 360.2 ab | 6.9 ns | 50.6 ns | 4.6 ab |
| S. aethiopicum (accession 2) | 360.1 b | 6.8 ns | 38.8 ns | 4.9 a |
| Significance | ||||
| Rootstock | *** | NS | NS | *** |
| Treatments | L0 Central Area | L0 Lateral Area | ΔL30 Lateral Area |
|---|---|---|---|
| Rootstock | |||
| ‘Scarlatti’ ungrafted | 82.4 bc | 80.6 b | 2.5 bc |
| S. torvum | 82.9 b | 81.0 b | 2.3 c |
| ‘Scarlatti’ self-grafted | 82.5 b | 80.6 b | 2.5 bc |
| Hybrid S. melongena × S. aethiopicum | 83.5 ab | 81.5 ab | 2.7 ab |
| S. aethiopicum (accession 1) | 84.8 a | 82.9 a | 2.7 ab |
| S. aethiopicum (accession2) | 80.7 c | 78.9 c | 2.9 a |
| Significance | |||
| Rootstock | *** | *** | *** |
| Treatments | Total Anthocyanins (mg∙100 g−1 of fw) | Glycoalkaloids (mg∙100 g−1 of fw) | Chlorogenic Acid (mg∙100 g−1 of fw) | Proteins (g∙100 g−1 of fw) |
|---|---|---|---|---|
| Rootstock | ||||
| ‘Scarlatti’ ungrafted | 60.6 ns | 8.4 a | 21.7 ab | 1.0 ab |
| S. torvum | 43.0 ns | 1.0 b | 40.3 a | 1.0 ab |
| ‘Scarlatti’ self-grafted | 47.8 ns | 2.7 b | 34.1 a | 1.0 ab |
| Hybrid S. melongena × S. aethiopicum | 70.8 ns | 8.6 a | 4.4 b | 0.9 bc |
| S. aethiopicum (accession 1) | 80.0 ns | 3.7 b | 40.0 a | 1.1 ab |
| S. aethiopicum (accession 2) | 106.4 ns | 1.9 b | 17.9 ab | 0.8 d |
| Significance | ||||
| Rootstock | NS | * | ** | *** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sabatino, L.; Iapichino, G.; Rotino, G.L.; Palazzolo, E.; Mennella, G.; D’Anna, F. Solanum aethiopicum gr. gilo and Its Interspecific Hybrid with S. melongena as Alternative Rootstocks for Eggplant: Effects on Vigor, Yield, and Fruit Physicochemical Properties of Cultivar ′Scarlatti′. Agronomy 2019, 9, 223. https://doi.org/10.3390/agronomy9050223
Sabatino L, Iapichino G, Rotino GL, Palazzolo E, Mennella G, D’Anna F. Solanum aethiopicum gr. gilo and Its Interspecific Hybrid with S. melongena as Alternative Rootstocks for Eggplant: Effects on Vigor, Yield, and Fruit Physicochemical Properties of Cultivar ′Scarlatti′. Agronomy. 2019; 9(5):223. https://doi.org/10.3390/agronomy9050223
Chicago/Turabian StyleSabatino, Leo, Giovanni Iapichino, Giuseppe Leonardo Rotino, Eristanna Palazzolo, Giuseppe Mennella, and Fabio D’Anna. 2019. "Solanum aethiopicum gr. gilo and Its Interspecific Hybrid with S. melongena as Alternative Rootstocks for Eggplant: Effects on Vigor, Yield, and Fruit Physicochemical Properties of Cultivar ′Scarlatti′" Agronomy 9, no. 5: 223. https://doi.org/10.3390/agronomy9050223
APA StyleSabatino, L., Iapichino, G., Rotino, G. L., Palazzolo, E., Mennella, G., & D’Anna, F. (2019). Solanum aethiopicum gr. gilo and Its Interspecific Hybrid with S. melongena as Alternative Rootstocks for Eggplant: Effects on Vigor, Yield, and Fruit Physicochemical Properties of Cultivar ′Scarlatti′. Agronomy, 9(5), 223. https://doi.org/10.3390/agronomy9050223