Urea Addition Promotes the Metabolism and Utilization of Nitrogen in Cucumber

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Nitrogen Treatment
2.3. Determination of Plant Yield, Dry Weight, and Nitrogen Content
2.4. Biochemical Analysis
2.5. Total RNA Isolation and cDNA Synthesis
2.6. Gene Expression Analysis by qRT-PCR
2.7. Statistical Analysis
3. Results
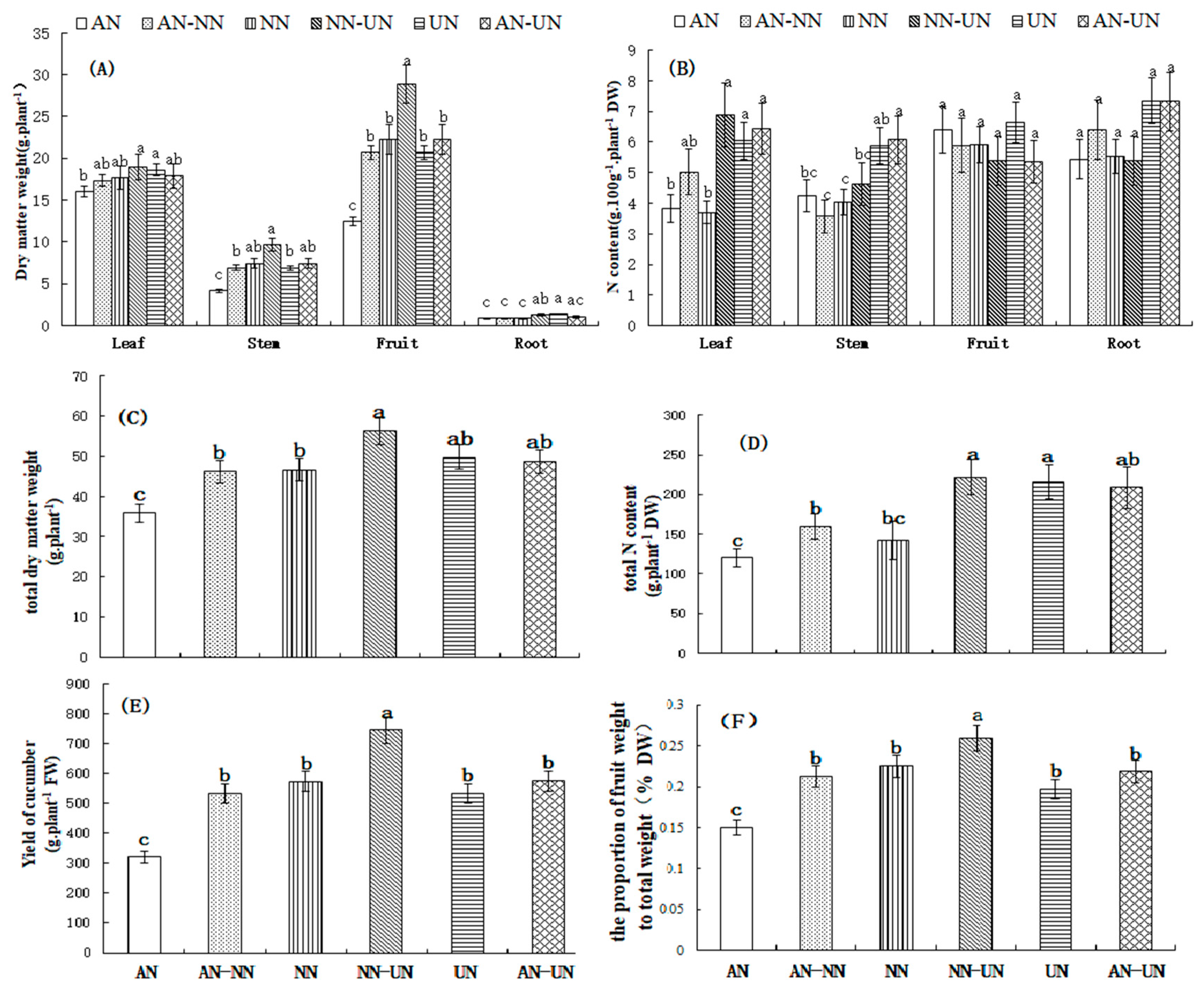
3.1. Effect of Different Nitrogen Forms on Cucumber Growth
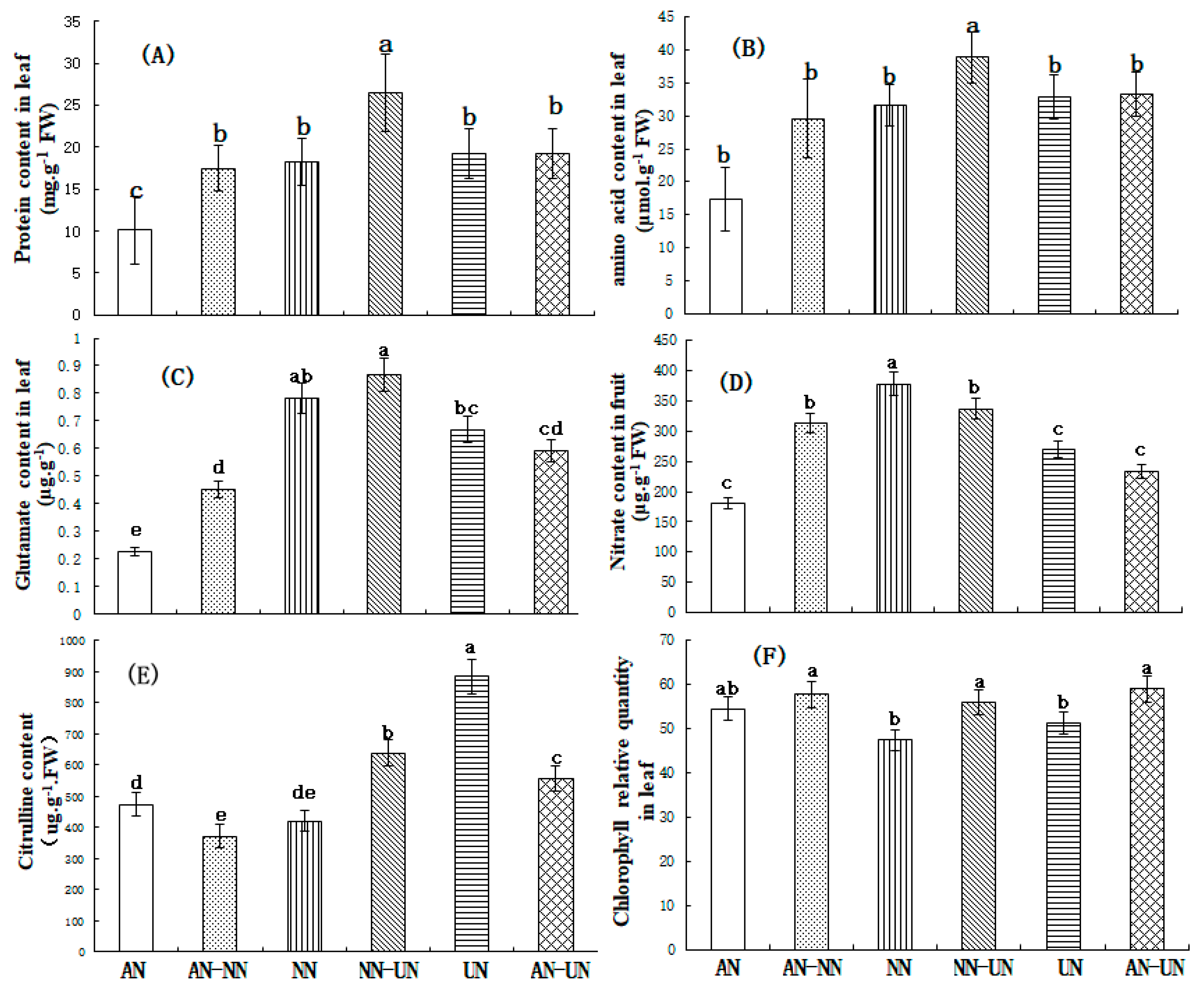
3.2. Effect of Different Nitrogen Forms on the Biochemical Metabolites in Cucumber
3.3. The Correlation between Biochemical Substance and the Growth Index of Cucumber
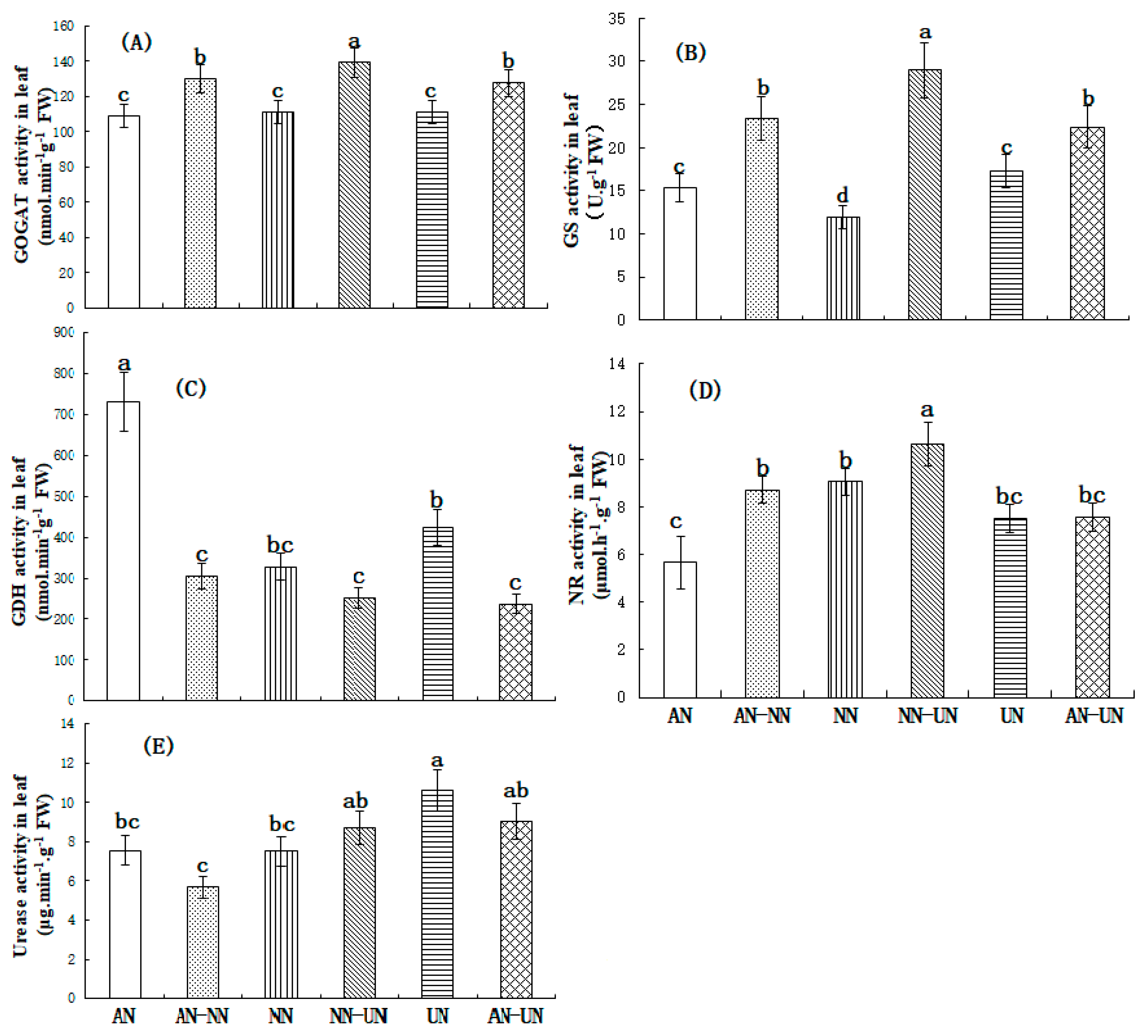
3.4. Effect of Different Nitrogen Forms on Nitrogen-Metabolizing Enzymes in Cucumber
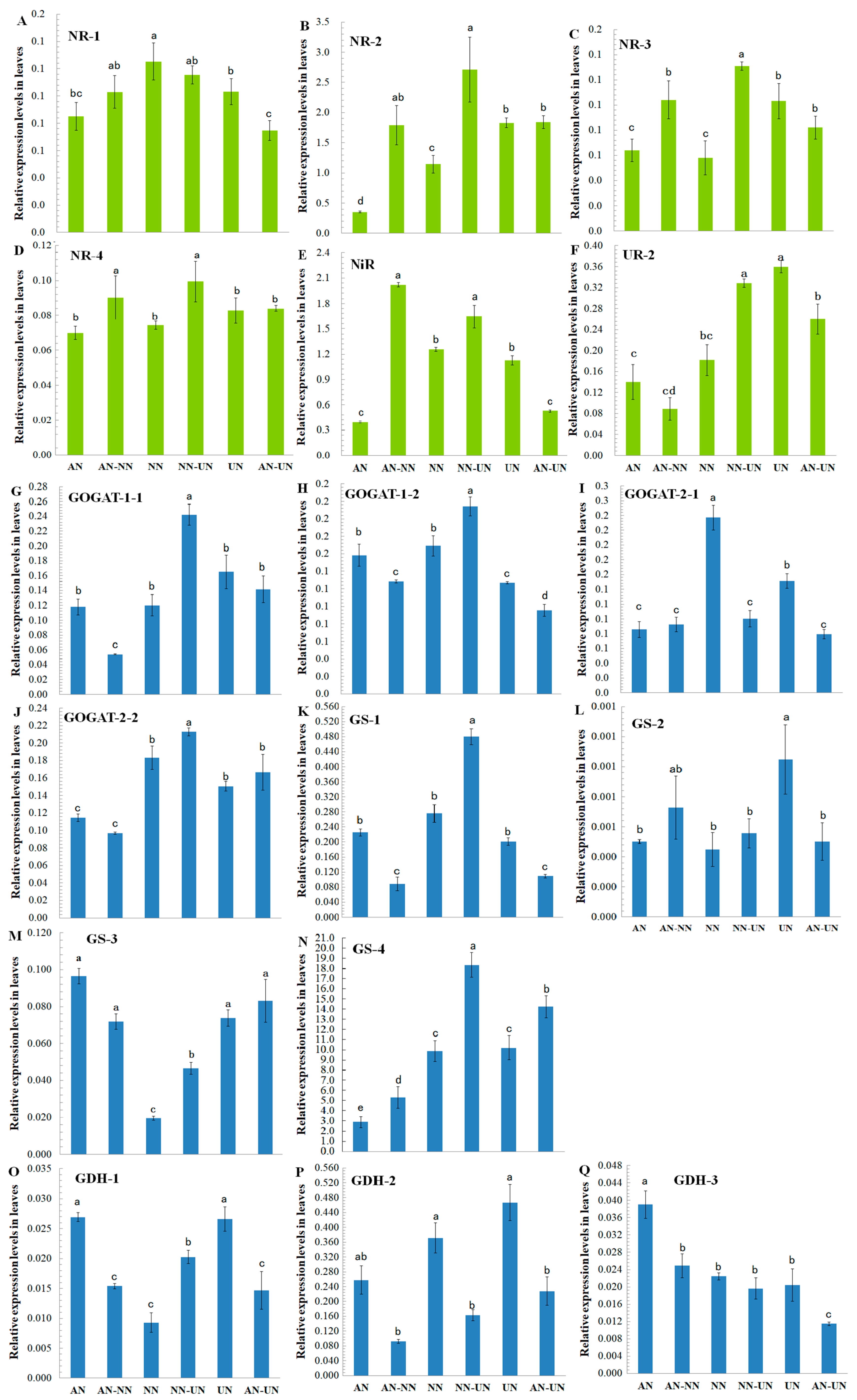
3.5. Expression Profiles of the N-Metabolism-Related Genes of Cucumber Treated with Different Nitrogen Forms
4. Discussion
4.1. Nitrogen Assimilation Response to Nitrogen Forms Prior to the Glutamate Cycle
4.2. Glutamate Cycle Response to Nitrogen Forms
4.3. Utilization of the Nitrogen Response to Nitrogen Forms
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yang, X.; Wang, X.; Wei, M.; Hikosaka, S.; Goto, E. Changes in growth and photosynthetic capacity of cucumber seedings in response to nitrate stress. Braz. J. Plant Phyalol. 2009, 21, 309–317. [Google Scholar] [CrossRef]
- Wu, T.Z.; Qin, L.; Fan, C.; Xue, X.; Zhou, M.; Xin, Y. Involvement of CsNRT1.7 in nitrate recycling during senescence in cucumber. Plant Nutr. Soil Sci. 2014, 177, 714–721. [Google Scholar] [CrossRef]
- Iqbal, N.; Umar, S.; Khan, N.A. Nitrogen availabiity regulates proline and ethylene production and alleviates salinity stress in mustard (Brassica juncea). J. Plant Physlol. 2015, 178, 84–91. [Google Scholar] [CrossRef]
- Muhammad, S.; Muhammad, Y.; Rashid, M.; Lftikhar, A. Effect of Soil applied CaC2 on gas exchange characters, nitrogen metabolism and fruit yield in cucumber (Cucumis sativus). N. Z. J. Crop Hortic. Sci. 2017, 45, 251–262. [Google Scholar] [CrossRef]
- Mi, G.; Chen, F.; Zhang, F. Multiple signaling pathways controls nitrogen-mediated root elongation in maize. Plant Signal Behav. 2008, 3, 1030–1032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohd-Radzman, N.A.; Djordjevic, M.A.; Imin, N. Nitrogen modulation of legume architecture root signaling pathways involves phytohormones and small regulatory molecules. Front. Plant Sci. 2013, 4, 385. [Google Scholar] [CrossRef]
- Zhao, W.; Yang, X.; Yu, H.; Jiang, W.; Sun, N.; Liu, X.; Liu, X.; Zhang, X.; Wang, Y.; Gu, X. RNA-seq-based transcriptome profiling of early nitrogen deficiency response in cucumber seedlings provides new insight into the putative nitrogen regulatory. Plant Cell Physiol. 2015, 56, 455–467. [Google Scholar] [CrossRef]
- Helali, S.M.R.; Nebli, H.; Kaddour, R. Influence of nitrate/ammonium ratio on growth and nutrition of Arabidopsis thaliana. Plant Soil 2010, 336, 65–74. [Google Scholar] [CrossRef]
- Lentherwood, N.R.; Peter, C. Nitrogen: All forms are not equal. Greenh. Manag. 2009, 29, 22–23. [Google Scholar]
- Antonio, L.; Maria, P.P.; Fabrizio, A.; Anthony, J.M.; Francesco, S.; Maria, R. Physiological and molecular responses in tomato under different forms of N nutrition. J. Plant Physiol. 2017, 216, 17–25. [Google Scholar] [CrossRef]
- Chen, L.; Zhu, Y.L.; Yang, L.F. Effects of nitrogen forms and ratios on plant growth, seed antioxidant enzyme activities and reactive oxygen metabolism of vegetable soybean. Plant Nutr. Fertil. Sci. 2010, 16, 768–772, (In Chinese with English Abstract). [Google Scholar]
- Miflin, B.J.; Habash, D.Z. The role of glutamine synthetase and glutamate dehydrogenase in nitrogen assimilation and possibilities for improvement in the nitrogen utilization of crops. J. Exp. Bot. 2002, 53, 979–987. [Google Scholar] [CrossRef] [Green Version]
- Aubert, S.; Bligny, R.; Douce, R.; Gout, E.; Ratcliffe, R.G.; Roberts, J.K.M. Contribution of glutamate dehydrogenase to mitochondrial glutamate metabolism studied by 13C and 31P nuclear magnetic resonance. J. Exp. Bot. 2001, 52, 37–45. [Google Scholar] [CrossRef]
- Zhou, Y.H.; Zhang, Y.L.; Wang, X.M.; Cui, J.X. Effects of nitrogen form on growth, CO2 assimilation, chlorophyll fluorescence, and photosynthetic electron allocation in cucumber and rice plants. J. Zhejiang Univ. Sci. B 2011, 12, 126–134. [Google Scholar] [CrossRef]
- Zanin, L.; Zamboni, A.; Monte, R.; Tomasi, N.; Varanini, Z.; Cesco, S.; Pinton, R. Transcriptomic analysis highlights reciprocal interactions of urea and nitrate for nitrogen acquisition by maize roots. Plant Cell Physiol. 2015, 56, 532–548. [Google Scholar] [CrossRef]
- Baligar, V.C.; Fageriat, N.K. Influence of nitrogen forms and levels on the growth and nutrition of cacao. J. Plant Nutr. 2017, 40, 709–718. [Google Scholar] [CrossRef]
- Marschner, H. Mineral nutrition in higher plants. Acad. Press Univ. Hohenh. Ger. 2011, 8, 147–148. [Google Scholar]
- Li, Q.Y.; Xu, X.J.; Gu, H.L.; Gao, H.Y.; Zhu, Y.Y.; Dong, C.X.; Shen, Q.R. Effects of applying different nitrogen form on cherry tomato nitrogen metabolism during fruit development. Chin. J. Appl. Ecol. 2010, 21, 2335–2341. [Google Scholar] [CrossRef]
- He, X.; Zhang, P.W.; Ding, C.Y. Effects of nitrate/ammonium ratio on nitrate absorption and distribution of carbon and nitrogen in pakchoi growing under low light intensity. Acta. Pedologica Sinica. 2009, 46, 452–458. [Google Scholar]
- Lu, J.M.; Yang, R.T.; Wang, H.C.; Huang, X.M. Stress effects of chlorate on longan (Dimocarpus longanlour) trees:changes in nitrogen and carbon nutrition. Hortic. Plant J. 2017, 3, 237–246. [Google Scholar] [CrossRef]
- Gangwar, S.; Singh, V.P. Indole acetic acid differently changes growth and nitrogen metabolism in Pisum sativum L. seedlings under chromium (VI) phytotoxicity: Implication of oxidative stress. Sci. Hortic. 2011, 129, 321–328. [Google Scholar] [CrossRef]
- Kotsiras, A.; Olympios, C.M.; Drosopoulos, J. Effects of nitrogen form and concentration on the distribution of ions within cucumber fruits. Sci. Hortic. 2002, 95, 175–183. [Google Scholar] [CrossRef]
- Narumol, P.; Xu, Z.; Hans, B. Nitrogen nutrition of Cyperus laevigatus and Phormium tenax: Effects of ammonium versus nitrate on growth, nitrate reductase activity and N uptake kinetics. Aquat. Bot. 2013, 106, 42–51. [Google Scholar] [CrossRef]
- Marschner, H. Mineral Nutrition of Higher Plants, 2nd ed.; California Academic Press: San Diego, CA, USA, 1995; pp. 138–146. [Google Scholar] [CrossRef]
- Yang, Y.; Zheng, Q.L.; Pei, C.G. Effects of NO3−-N/NH4+-N ratios on chardonnay grape seedling growth and nitrogen nutrition. Plant Nutr. Fertil. Sci. 2010, 16, 370–375. [Google Scholar] [CrossRef]
- Lu, Y.L.; Li, Q.Y.; Xu, X.J. Effects of different nitrogen forms on nutrient element contents in tomato seedlings. Chin. Agric. Sci. Bull. 2011, 26, 122–130, (In Chinese with English Abstract). [Google Scholar]
- Sui, L.; Yi, J.N.; Wang, K.C.; Li, Y.Q. Effects of different forms and ratios of nitrogen on physiological characteristic of Perilla frutescens (L.) Britt under salt stress. Chin. J. Ecol. 2018, 37, 3277–3283. [Google Scholar]
- Guan, X.L.; Wang, X.Z.; Liu, B.; Fan, S.S.; Chen, X.P. Yield, fruit quality of pepper and nitrogen loss under different amide/nitrate ratios in three types of soils. J. Plant Nutr. Fertil. 2017, 23, 730–739. [Google Scholar]
- Huang, Y.; Zhao, L.Q.; Kong, Q.S.; Cheng, F.; Niu, M.L.; Xie, J.J.; Muhammad, A.N.; Bie, Z.L. Comprehensive mineral nutrition analysis of watermelon grafted onto two different rootstocks. Hortic. Plant J. 2016, 2, 105–113. [Google Scholar] [CrossRef]
- Witte, C.; Tiller, S.A.; Taylor, M.A.; Davies, H.V. Leaf urea metabolism in potato. Urease activity profiles and patterns of recovery and distribution of 15N after foliar urea application in wild-type and urease-antisense transgenics. Plant Physiol. 2002, 129, 1129–1136. [Google Scholar] [CrossRef]
- Mustapha, A.; Xavier, S.; Laëtitia, J.; Philippe, L.; Philippe, E.; José, M. Hydroponics versus field lysimeter studies of urea, ammonium and nitrate uptake by oilseed rape (Brassica napus L.). J. Exp. Bot. 2012, 63, 5245–5258. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Hageman, R.H.; Hucklesby, D.P. Nitrate reductase from higher plants. Methods Enzymol. 1971, 23, 491–503. [Google Scholar] [CrossRef]
- Li, B.Z.; Xin, W.J.; Sun, S.B. Physiological and molecular responses of nitrogen-starved rice plants to re-supply of different nitrogen sources. Plant Soil 2006, 287, 145–159. [Google Scholar] [CrossRef]
- Cross, J.M.; Korff, M.; Altmann, T.; Bartzetko, L.; Sulpice, R.; Gibon, Y.; Palacios, N.; Stitt, M. Variation of enzyme activities and metabolite levels in 24 Arabidopsis accessions growing in carbon-limited conditions. Plant Physiol. 2006, 142, 1574–1588. [Google Scholar] [CrossRef]
- O’Neal, D.; Joy, K.D. Glutamine synthetase of pea leaves. I. Purification, stabilisation and pH optima. Arch. Biochem. Biophys. 1973, 159, 113–122. [Google Scholar] [CrossRef]
- Esposito, S.; Guerriero, G.; Vona, V.; Martino, V.; Carfagna, S.; Rigano, C. Glutamate synthase activities and protein changes in relation to nitrogen nutrition in barley: the dependence on different plastidic glucose-6P dehydrogenase isoforms. J. Exp. Bot. 2005, 56, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Turano, F.J.; Dashner, R.; Upadhyaya, A.; Caldwell, C.R. Purification of mitochondrial glutamate dehydrogenase from dark-grown soybean seedlings. Plant Physiol. 1996, 112, 1357–1364. [Google Scholar] [CrossRef]
- Bubner, B.; Baldwin, T. Use of real-time PCR for determining copy number and zygosity in transgenic plants. Plant Cell Rep. 2004, 23, 263–271. [Google Scholar] [CrossRef]
- Wang, J.f.; Dong, C.X.; Yue, X.; Shen, Q. Effects of NH4+-N/ NO3−-N ratios on growth, nitrate uptake and organic acid levels of spinach (Spinacia oleracea L.). Afr. J. Biotechnol. 2009, 15, 3597–3602. [Google Scholar] [CrossRef]
- Chen, G.L.; Gao, X.R. Effect of partial replacement of nitrate by amino acid and urea on nitrate content of nonheading Chinese cabbage and lettuce in hydroponics. Sci. Agric. Sin. 2002, 4, 444–449. [Google Scholar] [CrossRef]
- Zhu, Y.; Gang, L.; Liu, H.; Sun, G.; Chen, R.; Song, S. Effects of partial replacement of nitrate with different nitrogen forms on the yield, quality and nitrate content of Chinese kale. Commun. Soil Sci. Plant Anal. 2018, 49, 1384–1393. [Google Scholar] [CrossRef]
- Cao, F.Q.; Liu, G.W.; Wang, W.H.; Wu, X.M.; Liu, L.H. Molecular processes of urea metabolism and transport in higher plants. Chin. Bull. Bot. 2009, 44, 273–282. [Google Scholar] [CrossRef]
- Caroline, M.S.; Collier, J.; Berg, G.; Glibert, P. Role of urea in microbial metabolism in aquatic systems: a biochemical and molecular review. Aquat. Microb. Ecol. 2010, 59, 67–88. [Google Scholar] [CrossRef]
- Zhang, X.C.; Liu, Y.M.; Yu, X.C. Effects of different NO3−-N/NH4+-N ratios on cucumber seedlings growth, nitrogen absorption and metabolism under suboptimal temperature and light intensity. Chin. J. Appl. Ecol. 2016, 27, 2527–2534. [Google Scholar] [CrossRef]
- Zhang, S.Y.; Chu, G.X.; Liang, Y.C. Effects of enhancing ammonium nutrition on the nitrogenous metabolisms of cotton seedlings grown hydroponically under low-temperature stress. J. Plant Nutr. Fertil. 2017, 23, 983–990, (In Chinese with English Abstract). [Google Scholar]
- Céline, M.D.; Valadier, M.H.; Carrayol, E.; Haw, J. Diurnal changes in the expression of glutamate dehydrogenase and nitrate reductase are involved in the C/N balance of tobacco source leaves. Plant Cell Environ. 2002, 25, 1451–1462. [Google Scholar] [CrossRef]
- Villa, M.S.; Gonzalez, G.A.; Torres, J.L. Effect of the NH4+/NO3− ratio on GS and PEPcase activities and on dry matter prodution in wheat. J. Plant Nutr. Fertil. 1992, 15, 2545–2557. [Google Scholar] [CrossRef]
- Botella, M.A.; Martínez, V.; Nieves-Cerdá, M.A. Effect of salinity on the growth and nitrogen uptake by wheat seedlings. J. Plant Nutr. 1997, 20, 793–804. [Google Scholar] [CrossRef]
- Xu, R.Y.; Bie, Z.L.; Huang, D.F. Effects of different nitrogen forms on the dry matter accumulation and leaf nitrogen metabolism of muskmelon. Trans. CSAE. 2005, 21, 147–150, (In Chinese with English Abstract). [Google Scholar]
- Tian, X.H.; Li, S.H. Uptake capacity of several vegetable crops to nitrate and ammonium. Plant Nutr. Fertil. Sci. 2000, 6, 194–201, (In Chinese with English Abstract). [Google Scholar]
- Roosta, H.R.; Schjoerring, J.K. Effects of ammonium toxicity on nitrogen metabolism and elemental profile of cucumber plants. J. Plant. Nutr. 2007, 30, 1933–1951. [Google Scholar] [CrossRef]
- Baev, M.V.; Kiriukhin, M.Y. Regulation of ammonia assimilation in an obligate methylotroph Methylobacillus flagellatum under steady-state and transient growth conditions. Antonie Leeuwenhoek 1997, 71, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Lopes, M.S.; Araus, J.L. Comparative genomic and physiological analysis of nutrient response to NH4+, NH4+:NO3− and NO3− in barley seedlings. Physiol. Plant. 2008, 134, 134–150. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Single Nitrogen Forms | Treatments | Mixed Nitrogen Forms |
|---|---|---|---|
| AN(Control 1) | 100% NH4+-N | AN-NN | 50% NO3−-N + 50% NH4+-N |
| NN(Control 2) | 100% NO3−-N | NN-UN | 50% NO3−-N + 50% CO(NH2)2 |
| UN | 100% CO(NH2)2 | AN-UN | 50% NH4+-N + 50% CO(NH2)2 |
| Gene Abbreviation | Accession No. | Biological Process and Molecular Function | Function |
|---|---|---|---|
| NR-1 | XM004139616.2 | Cyclic pyranopterin monophosphate synthase accessory protein Mitochondrial transcript variant X1, | nitrate reduction |
| NR-2 | NM001280767.1 | nitrate reductase [NADH]-like (NR2) | nitrate reduction |
| NR-3 | XM004135853.2 | Cyclic pyranopterin monophosphate synthase, mitochondrial | nitrate reduction |
| NR-4 | XM011661764.1 | Molybdopterin biosynthesis protein CNX1 | nitrate reduction |
| NiR | XM004140647.2 | Ferredoxin-nitrite reductase, chloroplastic (NIR) | nitrite reduction |
| GS-1 | NM001280715.1 | Glutamine synthetase cytosolic isozyme-like (GS1) | glutamine synthesis |
| GS-2 | XM011661119.1 | Glutamine synthetase nodule isozyme-like | glutamine synthesis |
| GS-3 | XM011656924.1 | Glutamine synthetase nodule isozyme | glutamine synthesis |
| GS-4 | XM004134113.2 | Glutamine synthetase leaf isozyme, chloroplastic transcript variant X1 | glutamine synthesis |
| GOGAT-1-1 | XM004136730.2 | Ferredoxin-dependent glutamate synthase, chloroplastic, transcript variant X1 | glutamate synthesis |
| GOGAT-1-2 | XM011653889.1 | Ferredoxin-dependent glutamate synthase, chloroplastic, transcript variant X2 | glutamate synthesis |
| GOGAT-2-1 | XM011653296.1 | Glutamate synthase [NADH], amyloplastic, transcript variant X1 | glutamate synthesis |
| GOGAT-2-2 | XM011653298.1 | Glutamate synthase [NADH], amyloplastic, transcript variant X3 | glutamate synthesis |
| GDH-1 | XM004147487.2 | Glutamate dehydrogenase 1, transcript variant X2 | glutamate dehydrogenation |
| GDH-2 | XM004146845.2 | Glutamate dehydrogenase 2 | glutamate dehydrogenation |
| GDH-3 | XM004143618.2 | Glutamate dehydrogenase 2-like | glutamate dehydrogenation |
| Urease-1 | XM011657264.1 | Urease, transcript variant X2 | urea decomposition |
| Urease-2 | XM011659065.1 | Urease accessory protein G | urea decomposition |
| Gene | Forward Primer (5′) | Reverse Primer (3′) | Length |
|---|---|---|---|
| NR-1 | TACTTCGGCTTTGACTCATGTTG | GTATGTTTTGCTCCGCTTATTCC | 207 bp |
| NR-2 | TACTGGTGCTGGTGTTTCTGGTC | GATTTCTCCCTTGTGAGGTTTGC | 195 bp |
| NR-3 | GAACTGTGTTATAATGCGTGGTT | CAAGTCGTGTAAGGTTTGTGAAG | 197 bp |
| NR-4 | CAGAGAACACAGAAAAGAAGGAA | CACAATGAAACAGTGGTCGAATA | 207 bp |
| NiR | GTCCCTCTCTGTGGAGCCATCTT | CCCTTCTTTCCCATTGCTTATTT | 199 bp |
| GS-1 | TTCTTTCTTTTGATCCAAAACCA | ATGTCGCCCTGTGAGACGACGCT | 197 bp |
| GS-2 | CAAGTCGGTCCTACCGTTGGTATTG | TCGAAGTAGACCTGTAATTGGTG | 188 bp |
| GS-3 | CTTTTGACCCCAAACCAATTCAG | GTGTCGACCAGTTAGACGACGCT | 191 bp |
| GS-4 | GTGCCCATCCCTACAAACAAACG | ACACCACAGTAATAAGGCCCCTG | 185 bp |
| GOGAT-1-1 | GAACGAGAACTTTACATTTGTAG | CTATATCTTCGATGATAAATAGC | 206 bp |
| GOGAT-1-2 | GAAATTGATTGAAAGAGAAGCAA | CTATATCTTCGATGATAAATAGC | 183 bp |
| GOGAT-2-1 | AGTTGGGATCGTGCTCAGCCT | CTAATTAAAAGCTCAAGAACACC | 216 bp |
| GOGAT-2-2 | ATGCGTGTTTTGGGCCACAATG | CTAATTAAAAGCTCAAGAACACC | 194 bp |
| GDH-1 | GCAATCCTGGAGAATTAAGTATA | AGAGATCCACCTAGATCAATAGG | 217 bp |
| GDH-2 | TAAAGAAAGTACTGGAAGCCTTG | TCATCTGCCTCTGGATCTGTGGG | 195 bp |
| GDH-3 | ATTGATGTACCTGAGCTGATTATTCA | ATCTGCTTCTGGATCAGTAGGATG | 219 bp |
| Urease-1 | GATGGCTTCATTATGACCCTTGG | TGTGATGCCGCTTGATATTGCTT | 195 bp |
| Urease-2 | GAAAGGATTAGAGCAGTGGAAAC | AGACACATCGATGATATAGATGA | 201 bp |
| Correlation Coefficient (r) | Leaf Dry Matter (g·plant−1) | Total Dry Matte (g·plant−1) | Leaf N Content (g·plant−1) | Leaf Protein Content (g·plant−1) | Total N Content (g·plant−1) | Nitrate (μg·g−1) |
|---|---|---|---|---|---|---|
| Leaf dry matter (g·plant−1) | -- | 0.987 ** | -- | -- | -- | -- |
| Leaf N content (g·plant−1) | 0.895 * | 0.861 * | -- | -- | -- | -- |
| Leaf protein content (mg·g−1) | 0.930 ** | 0.968 ** | 0.768 | -- | -- | -- |
| Leaf amino acid content (μmol·g−1) | 0.954 ** | 0.985 ** | 0.741 | 0.967 ** | 0.827 * | -- |
| Total N content (g·plant−1) | 0.895 * | 0.888 * | 0.962 ** | 0.813 * | -- | |
| Yield (g·plant−1) | 0.885 * | 0.939 ** | 0.684 | 0.986 ** | 0.731 | -- |
| NR (μmol·h−1·g−1) | -- | -- | -- | -- | -- | 0.871 * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, C.; Ban, T.; Yu, H.; Li, Q.; Li, X.; Jiang, W.; Xie, J. Urea Addition Promotes the Metabolism and Utilization of Nitrogen in Cucumber. Agronomy 2019, 9, 262. https://doi.org/10.3390/agronomy9050262
Ma C, Ban T, Yu H, Li Q, Li X, Jiang W, Xie J. Urea Addition Promotes the Metabolism and Utilization of Nitrogen in Cucumber. Agronomy. 2019; 9(5):262. https://doi.org/10.3390/agronomy9050262
Chicago/Turabian StyleMa, Chao, Tiantian Ban, Hongjun Yu, Qiang Li, Xiaohui Li, Weijie Jiang, and Jianming Xie. 2019. "Urea Addition Promotes the Metabolism and Utilization of Nitrogen in Cucumber" Agronomy 9, no. 5: 262. https://doi.org/10.3390/agronomy9050262
APA StyleMa, C., Ban, T., Yu, H., Li, Q., Li, X., Jiang, W., & Xie, J. (2019). Urea Addition Promotes the Metabolism and Utilization of Nitrogen in Cucumber. Agronomy, 9(5), 262. https://doi.org/10.3390/agronomy9050262