Wx Gene in Hordeum chilense: Chromosomal Location and Characterisation of the Allelic Variation in the Two Main Ecotypes of the Species



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
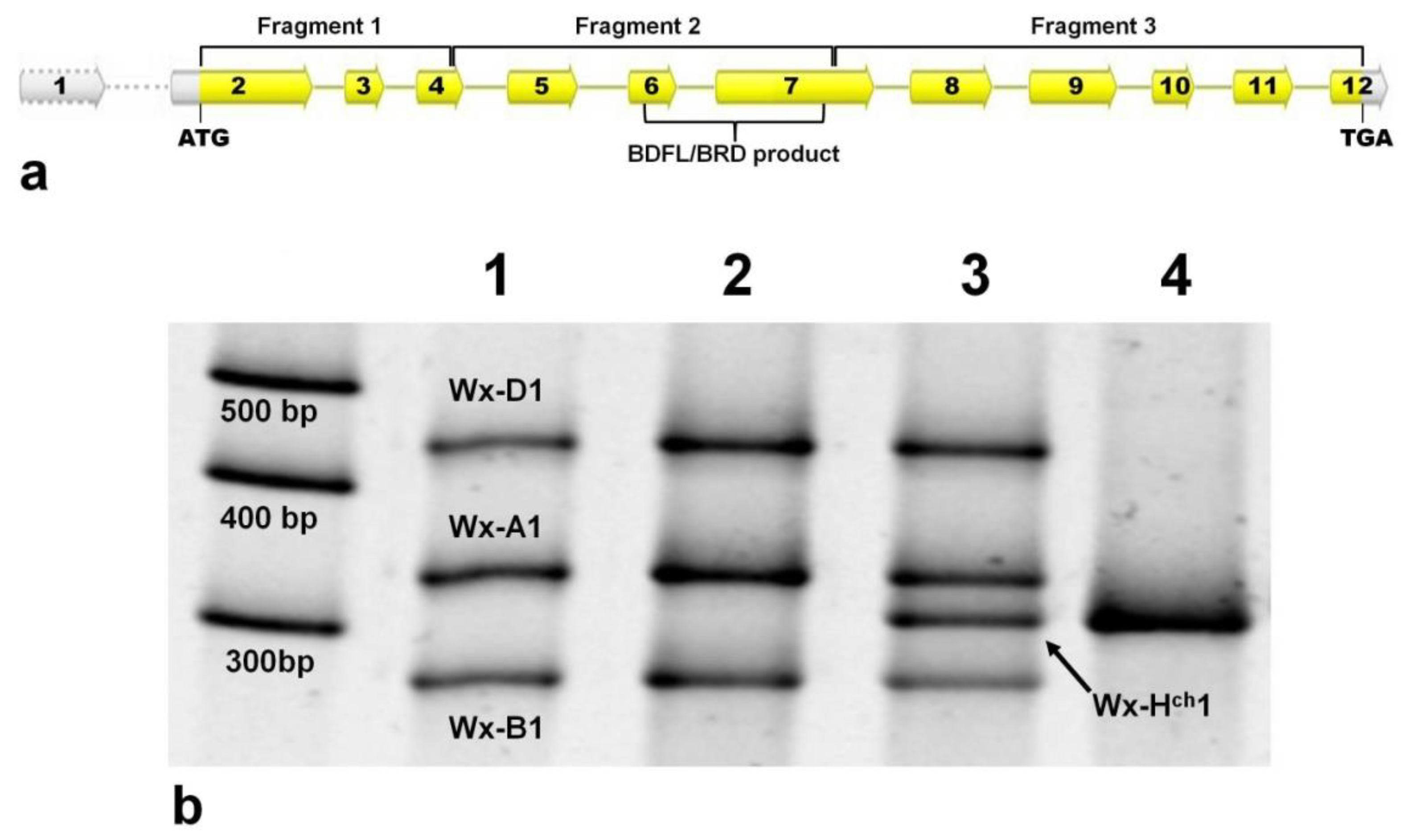
2.2. DNA Extraction and PCR Amplification
2.3. Cloning of PCR Products and Sequencing Analysis
2.4. Data Analysis
3. Results
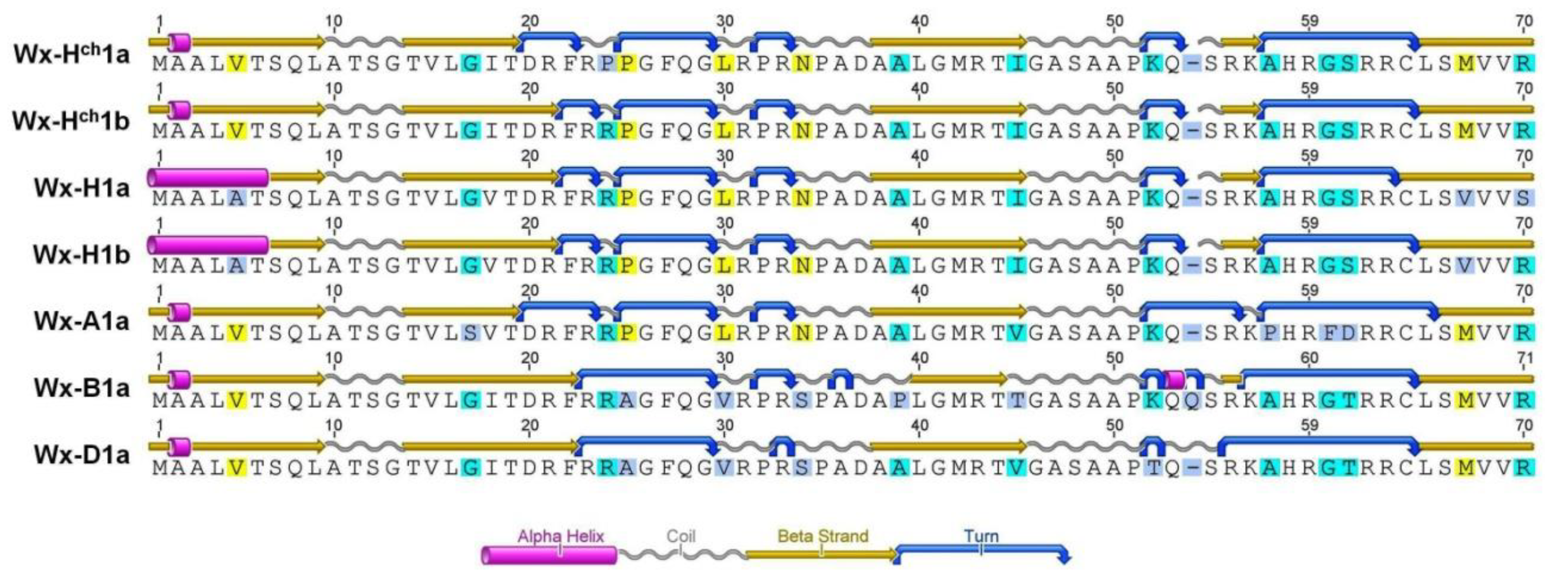
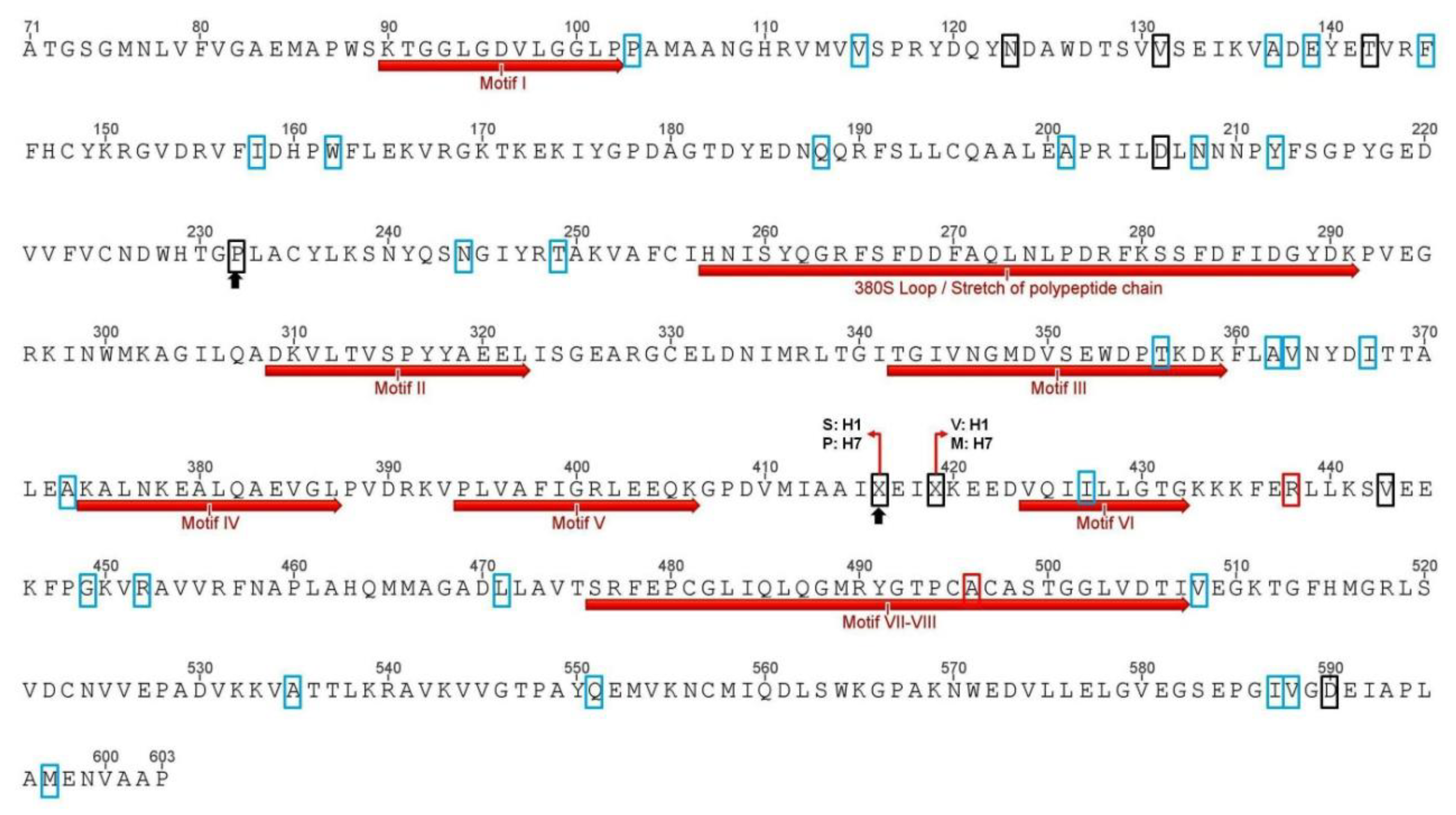
3.1. Amino acid Predicted Sequence Analysis
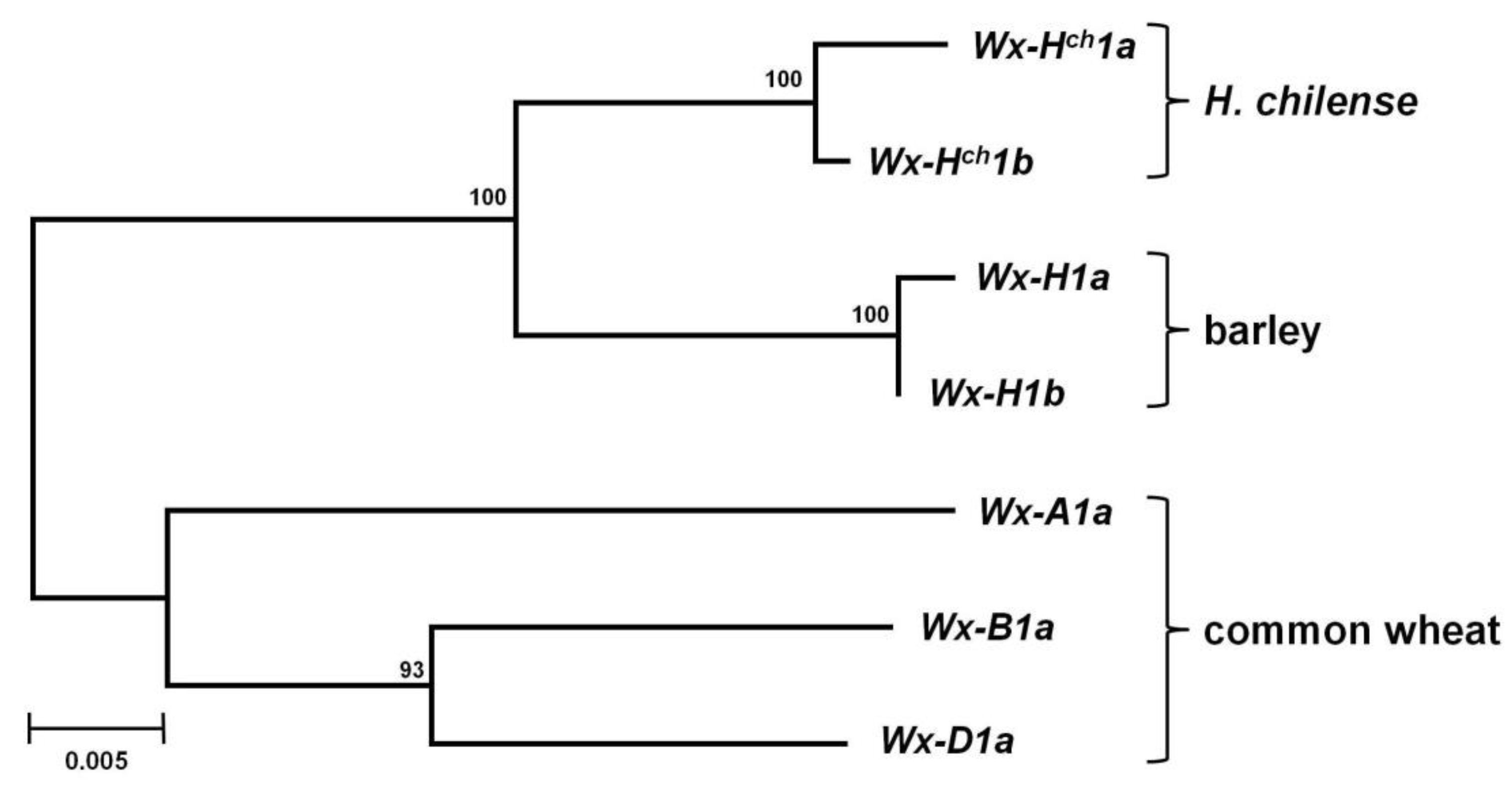
3.2. Phylogenetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- James, M.G.; Denyer, K.; Myers, A.M. Starch synthesis in the cereal endosperm. Curr. Opin. Plant Biol. 2003, 6, 215–222. [Google Scholar] [CrossRef]
- Zeng, M.; Morris, C.F.; Batey, I.L.; Wrigley, C.W. Sources of variation for starch gelatinization, pasting, and gelation properties in wheat. Cereal Chem. 1997, 74, 63–71. [Google Scholar] [CrossRef]
- Martin, J.M.; Sherman, J.D.; Lanning, S.P.; Talbert, L.E.; Giroux, M.J. Effect of variation in amylose content and puroindoline composition on bread quality in a hard spring wheat population. Cereal Chem. 2008, 85, 266–269. [Google Scholar] [CrossRef]
- Miura, H.; Tanii, S. Endosperm starch properties in several wheat cultivars preferred for Japanese noodles. Euphytica 1994, 72, 171–175. [Google Scholar] [CrossRef]
- Park, C.S.; Baik, B.-K. Characteristics of French bread baked from wheat flours of reduced starch amylose content. Cereal Chem. 2007, 84, 437–442. [Google Scholar] [CrossRef]
- Hayakawa, K.; Tanaka, K.; Nakamura, T.; Endo, S.; Hoshino, T. End use quality of waxy wheat flour in various grain-based foods. Cereal Chem. 2004, 81, 666–672. [Google Scholar] [CrossRef]
- Regina, A.; Bird, A.; Topping, D.; Bowden, S.; Freeman, J.; Barsby, T.; Kosar-Hashemi, B.; Li, Z.; Rahman, S.; Morell, M. High-amylose wheat generated by RNA interference improves indices of large-bowel health in rats. Proc. Natl. Acad. Sci. USA 2006, 103, 3546–3551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldwin, P.M. Starch Granule-Associated Proteins and Polypeptides: A Review. Starch-Stärke 2001, 53, 475–503. [Google Scholar] [CrossRef]
- Echt, C.S.; Schwartz, D. Evidence for the inclusion of controlling elements within the structural gene at the Waxy locus in maize. Genetics 1981, 99, 275–284. [Google Scholar]
- Yamamori, M.; Nakamura, T.; Endo, T.R.; Nagamine, T. Waxy protein deficiency and chromosomal location of coding genes in common wheat. Theor. Appl. Genet. 1994, 89, 179–184. [Google Scholar] [CrossRef]
- Mason-Gamer, R.J.; Weil, C.F.; Kellogg, E.A. Granule-bound starch synthase: Structure, function, and phylogenetic utility. Mol. Biol. Evol. 1998, 15, 1658–1673. [Google Scholar] [CrossRef] [PubMed]
- Rohde, W.; Becker, D.; Salamini, F. Structural analysis of the waxy locus from Hordeum vulgare. Nucleic Acids Res. 1988, 16, 7185–7186. [Google Scholar] [CrossRef]
- Guzmán, C.; Alvarez, J.B. Wheat waxy proteins: Polymorphism, molecular characterization and effects on starch properties. Theor. Appl. Genet. 2016, 129, 1–16. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, R.A.; Yamazaki, Y.; Dubcovsky, J.; Rogers, W.J.; Morris, G.; Appels, R.; Xia, X.C. Catalogue of gene symbols for wheat. 2013. Available online: http://www.shigen.nig.ac.jp/wheat/komugi/genes/macgene/2013/GeneSymbol.pdf. (accessed on 23 August 2018).
- Alvarez, J.B.; Guzmán, C. Interspecific and intergeneric hybridization as a source of variation for wheat grain quality improvement. Theor. Appl. Genet. 2018, 131, 225–251. [Google Scholar] [CrossRef] [PubMed]
- Martín, A.; Alvarez, J.B.; Martín, L.M.; Barro, F.; Ballesteros, J. The development of tritordeum: A novel cereal for food processing. J. Cereal Sci. 1999, 30, 85–95. [Google Scholar] [CrossRef]
- Von Bothmer, R.; Jacobsen, N.; Baden, C.; Jorgensen, R.; Linde-Laursen, I. An Ecogeografical Study of the Genus Hordeum, 2nd ed.; International Plant Genetic Resources Institute: Rome, Italy, 1995. [Google Scholar]
- Alvarez, J.B.; Martín, A.; Martín, L.M. Variation in the high-molecular-weight glutenin subunits coded at the Glu-Hch1 locus in Hordeum chilense. Theor. Appl. Genet. 2001, 102, 134–137. [Google Scholar] [CrossRef]
- Alvarez, J.B.; Broccoli, A.; Martín, L.M. Variability and genetic diversity for gliadins in natural populations of Hordeum chilense Roem. et Schult. Genet. Resour. Crop Evol. 2006, 53, 1419–1425. [Google Scholar] [CrossRef]
- Atienza, S.G.; Giménez, M.J.; Martín, A.; Martín, L.M. Variability in monomeric prolamins in Hordeum chilense. Theor. Appl. Genet. 2000, 101, 970–976. [Google Scholar] [CrossRef]
- Atienza, S.G.; Alvarez, J.B.; Villegas, A.M.; Gimenez, M.J.; Ramirez, M.C.; Martín, A.; Martín, L.M. Variation for the low-molecular-weight glutenin subunits in a collection of Hordeum chilense. Euphytica 2002, 128, 269–277. [Google Scholar] [CrossRef]
- Alvarez, J.B.; Campos, L.A.C.; Martín, A.; Martín, L.M. Influence of HMW and LMW glutenin subunits on gluten strength in hexaploid tritordeum. Plant Breed. 1999, 118, 456–458. [Google Scholar] [CrossRef]
- Martinek, P.; Svobodová, I.; Věchet, L. Selection of the wheat genotypes and related species with resistance to Mycosphaerella graminicola. Agriculture 2013, 59, 65–73. [Google Scholar] [CrossRef]
- Rodríguez-Suárez, C.; Mellado-Ortega, E.; Hornero-Méndez, D.; Atienza, S. Increase in transcript accumulation of Psy1 and e-Lcy genes in grain development is associated with differences in seed carotenoid content between durum wheat and tritordeum. Plant Mol. Biol. 2014, 84, 659–673. [Google Scholar] [CrossRef] [PubMed]
- Martín, A.; Martín, L.M.; Cabrera, A.; Ramírez, M.C.; Giménez, M.J.; Rubiales, D.; Hernández, P.; Ballesteros, J. The potential of Hordeum chilense in breeding Triticeae species. In Triticeae III; Jaradat, A.A., Humphreys, M., Eds.; Science Publishers Inc.: Enfield, UK, 1998; pp. 377–386. [Google Scholar]
- Tobes, N.; Ballesteros, J.; Martínez, C.; Lovazzano, G.; Contreras, D.; Cosio, F.; Gastó, J.; Martín, L.M. Collection mission of H. chilense Roem. et Schult. in Chile and Argentina. Genet. Resour. Crop Evol. 1995, 42, 211–216. [Google Scholar] [CrossRef]
- Giménez, M.J.; Cosío, F.; Martínez, C.; Silva, F.; Zuleta, A.; Martín, L.M. Collecting Hordeum chilense Roem. et Schult. germplasm in desert and steppe dominions of Chile. Plant Genet. Resour. Newsl. 1997, 109, 17–19. [Google Scholar]
- Pistón, F.; Shewry, P.; Barro, F. D hordeins of Hordeum chilense: A novel source of variation for improvement of wheat. Theor. Appl. Genet. 2007, 115, 77–86. [Google Scholar] [CrossRef]
- Guzmán, C.; Alvarez, J.B. Molecular characterization of two novel alleles of Hordoindoline genes in Hordeum chilense Roem. et Schult. Genet. Resour. Crop Evol. 2014, 61, 307–312. [Google Scholar] [CrossRef]
- Alvarez, J.B.; Martín, L.M.; Martín, A. Chromosomal localization of genes for carotenoid pigments using addition lines of Hordeum chilense in wheat. Plant Breed. 1998, 117, 287–289. [Google Scholar] [CrossRef]
- Rodríguez-Suárez, C.; Atienza, S.G.; Pistón, F. Allelic variation, alternative splicing and expression analysis of Psy1 gene in Hordeum chilense Roem. et Schult. PLoS ONE 2011, 6, e19885. [Google Scholar] [CrossRef] [PubMed]
- Stacey, J.; Isaac, P.G. Isolation of DNA from Plants. In Protocols for Nucleic Acid Analysis by Nonradioactive Probes; Isaac, P.G., Ed.; Humana Press: Totowa, NJ, USA, 1994; pp. 9–15. [Google Scholar]
- Nakamura, T.; Vrinten, P.; Saito, M.; Konda, M. Rapid classification of partial waxy wheats using PCR-based markers. Genome 2002, 45, 1150–1156. [Google Scholar] [CrossRef]
- Guzman, C.; Alvarez, J.B. Molecular characterization of a novel waxy allele (Wx-Au1a) from Triticum urartu Thum. ex Gandil. Genet. Resour. Crop Evol. 2012, 59, 971–979. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed]
- Dvorak, J.; Akhunov, E.D. Tempos of Gene Locus Deletions and Duplications and Their Relationship to Recombination Rate During Diploid and Polyploid Evolution in the Aegilops-Triticum Alliance. Genetics 2005, 171, 323–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rice, P.; Longden, I.; Bleasby, A. EMBOSS: The European molecular biology open software suite. Trends Genet. 2000, 16, 276–277. [Google Scholar] [CrossRef]
- Choi, Y.; Chan, A.P. PROVEAN web server: A tool to predict the functional effect of amino acid substitutions and indels. Bioinformatics 2015, 31, 2745–2747. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Sims, G.E.; Murphy, S.; Miller, J.R.; Chan, A.P. Predicting the Functional Effect of Amino Acid Substitutions and Indels. PLoS ONE 2012, 7, e46688. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Zuckerkandl, E.; Pauling, L. Evolutionary Divergence and Convergence in Proteins. In Evolving Genes and Proteins; Bryson, V., Vogel, H.J., Eds.; Academic Press: New York, NY, USA, 1965; pp. 97–166. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Murai, J.; Taira, T.; Ohta, D. Isolation and characterization of the three Waxy genes encoding the granule-bound starch synthase in hexaploid wheat. Gene 1999, 234, 71–79. [Google Scholar] [CrossRef]
- Leterrier, M.; Holappa, L.; Broglie, K.; Beckles, D. Cloning, characterisation and comparative analysis of a starch synthase IV gene in wheat: Functional and evolutionary implications. BMC Plant Biol. 2008, 8, 98. [Google Scholar] [CrossRef]
- Guzmán, C.; Caballero, L.; Martín, L.M.; Alvarez, J.B. Waxy genes from spelt wheat: New alleles for modern wheat breeding and new phylogenetic inferences about the origin of this species. Ann. Bot. 2012, 110, 1161–1171. [Google Scholar] [CrossRef]
- Cornejo-Ramírez, Y.I.; Martínez-Cruz, O.; Del Toro-Sánchez, C.L.; Wong-Corral, F.J.; Borboa-Flores, J.; Cinco-Moroyoqui, F.J. The structural characteristics of starches and their functional properties. CyTA-J. Food 2018, 16, 1003–1017. [Google Scholar]
- Yan, L.; Bhave, M.; Fairclough, R.; Konik, C.; Rahman, S.; Appels, R. The genes encoding granule-bound starch synthases at the waxy loci of the A, B, and D progenitors of common wheat. Genome 2000, 43, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Mason-Gamer, R.J. Origin of North American Elymus (Poaceae: Triticeae) allotetraploids based on Granule-bound starch synthase gene sequences. Syst. Bot. 2001, 26, 757–768. [Google Scholar]
- Ingram, A.L.; Doyle, J.J. The origin and evolution of Eragrostis tef (Poaceae) and related polyploids: Evidence from nuclear waxy and plastid rps16. Am. J. Bot. 2003, 90, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Fortune, P.M.; Schierenbeck, K.A.; Ainouche, A.K.; Jacquemin, J.; Wendel, J.F.; Ainouche, M.L. Evolutionary dynamics of Waxy and the origin of hexaploid Spartina species (Poaceae). Mol. Phylogenet. Evol. 2007, 43, 1040–1055. [Google Scholar] [CrossRef] [PubMed]
- Ortega, R.; Alvarez, J.B.; Guzmán, C. Characterization of the Wx gene in diploid Aegilops species and its potential use in wheat breeding. Genet. Resour. Crop Evol. 2014, 61, 369–382. [Google Scholar] [CrossRef]
- Kramer, H.H.; Blander, B.A.S. Orienting linkage maps on the chromosomes of barley. Crop Sci. 1961, 1, 339–342. [Google Scholar] [CrossRef]
- Mattera, M.G.; Ávila, C.M.; Atienza, S.G.; Cabrera, A. Cytological and molecular characterization of wheat-Hordeum chilense chromosome 7Hch introgression lines. Euphytica 2015, 203, 165–176. [Google Scholar] [CrossRef]
- Avila, C.M.; Mattera, M.G.; Rodríguez-Suárez, C.; Palomino, C.; Ramírez, M.C.; Martín, A.; Kilian, A.; Hornero-Méndez, D.; Atienza, S.G. Diversification of seed carotenoid content and profile in wild barley (Hordeum chilense Roem. et Schultz.) and Hordeum vulgare L.-H. chilense synteny as revealed by DArTSeq markers. Euphytica 2019, 215, 45. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wx-A1a1 | Wx-B1a1 | Wx-D1a1 | Wx-H1a2/Wx-H1b3 | Wx-Hch1a/Wx-Hch1b | |
|---|---|---|---|---|---|
| Exon 2 | 321 | 324 | 321 | 318 | 318 |
| Exon 3 | 81 | 81 | 81 | 81 | 81 |
| Exon 4 | 99 | 99 | 99 | 99 | 99 |
| Exon 5 | 154 | 154 | 154 | 154 | 154 |
| Exon 6 | 101 | 101 | 101 | 101 | 101 |
| Exon 7 | 354 | 354 | 354 | 354 | 354 |
| Exon 8 | 180 | 180 | 180 | 180 | 180 |
| Exon 9 | 192 | 192 | 192 | 192 | 192 |
| Exon 10 | 87 | 87 | 87 | 87 | 87 |
| Exon 11 | 129 | 129 | 129 | 129 | 129 |
| Exon 12 | 117 | 117 | 117 | 117 | 117 |
| Intron 2 | 82 | 99 | 90 | 89 | 85 |
| Intron 3 | 84 | 88 | 95 | 84 | 80 |
| Intron 4 | 109 | 113 | 104 | 126 | 109 |
| Intron 5 | 125 | 133 | 152 | 136 | 113 |
| Intron 6 | 99 | 69 | 141 | 106 | 89 |
| Intron 7 | 91 | 92 | 85 | 92 | 89 |
| Intron 8 | 95 | 86 | 82 | 94 | 94 |
| Intron 9 | 90 | 84 | 84 | 82 | 82 |
| Intron 10 | 98 | 97 | 98 | 97 | 97 |
| Intron 11 | 93 | 115 | 116 | 76 | 85/86 |
| Total | 2781 | 2794 | 2862 | 2794 | 2735/2736 |
| Position 1 | Wx-Hch1a/b | Wx-H1a2 | Wx-A1a2 | Wx-B1a2 | Wx-D1a2 |
|---|---|---|---|---|---|
| 103 | Pro | Ala | |||
| 115 | Val | Ile | Ile | Ile | |
| 123 | Asn | Lys | Lys | Lys | Lys |
| 131 | Val | Ile | Ile | ||
| 137 | Ala | Val | Val | ||
| 139 | Glu | Arg | Lys | ||
| 142 | Thr | Arg | Arg | Arg | Arg |
| 145 | Phe | Tyr | Tyr | Tyr | |
| 158 | Ile | Val | Val | Val | |
| 162 | Trp | Cys | Cys | Cys | |
| 189 | Gln | Leu | |||
| 201 | Ala | Val | Val | ||
| 206 | Asp | Asn | Asn | ||
| 208 | Asn | Asp | |||
| 212 | Tyr | His | |||
| 232 | Pro | Leu | Leu | Leu | Leu |
| 244 | Asn | Ser | |||
| 249 | Thr | Ala | |||
| 356 | Thr | Ile | Ala | ||
| 362 | Ala | Thr | |||
| 363 | Val | Ala | |||
| 367 | Ile | Val | Val | ||
| 373 | Ala | Gly | Gly | Gly | |
| 416 | Ser/Pro | Pro | Pro | Pro | Pro |
| 419 | Val/Met | Leu | Leu | Leu | |
| 427 | Ile | Val | Val | Val | |
| 438 | Arg | Lys | |||
| 443 | Val | Met | Ile | ||
| 449 | Gly | Thr | Ser | Ser | |
| 452 | Arg | Trp | |||
| 471 | Leu | Val | Val | Val | |
| 496 | Ala | Val | |||
| 508 | Val | Met | |||
| 535 | Ala | Val | Val | Val | |
| 551 | Gln | His | His | His | |
| 587 | Ile | Val | Val | ||
| 588 | Val | Ile | Ile | ||
| 590 | Asp | Glu | Glu | Glu | Glu |
| 597 | Met | Leu |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alvarez, J.B.; Castellano, L.; Recio, R.; Cabrera, A. Wx Gene in Hordeum chilense: Chromosomal Location and Characterisation of the Allelic Variation in the Two Main Ecotypes of the Species. Agronomy 2019, 9, 261. https://doi.org/10.3390/agronomy9050261
Alvarez JB, Castellano L, Recio R, Cabrera A. Wx Gene in Hordeum chilense: Chromosomal Location and Characterisation of the Allelic Variation in the Two Main Ecotypes of the Species. Agronomy. 2019; 9(5):261. https://doi.org/10.3390/agronomy9050261
Chicago/Turabian StyleAlvarez, Juan B., Laura Castellano, Rocío Recio, and Adoración Cabrera. 2019. "Wx Gene in Hordeum chilense: Chromosomal Location and Characterisation of the Allelic Variation in the Two Main Ecotypes of the Species" Agronomy 9, no. 5: 261. https://doi.org/10.3390/agronomy9050261
APA StyleAlvarez, J. B., Castellano, L., Recio, R., & Cabrera, A. (2019). Wx Gene in Hordeum chilense: Chromosomal Location and Characterisation of the Allelic Variation in the Two Main Ecotypes of the Species. Agronomy, 9(5), 261. https://doi.org/10.3390/agronomy9050261