Tolerance of Three Quinoa Cultivars (Chenopodium quinoa Willd.) to Salinity and Alkalinity Stress During Germination Stage

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Germination Assay
2.3. Biometric Measurements
2.4. Proline Determination
2.5. Statistical Analysis
3. Results and Discussion
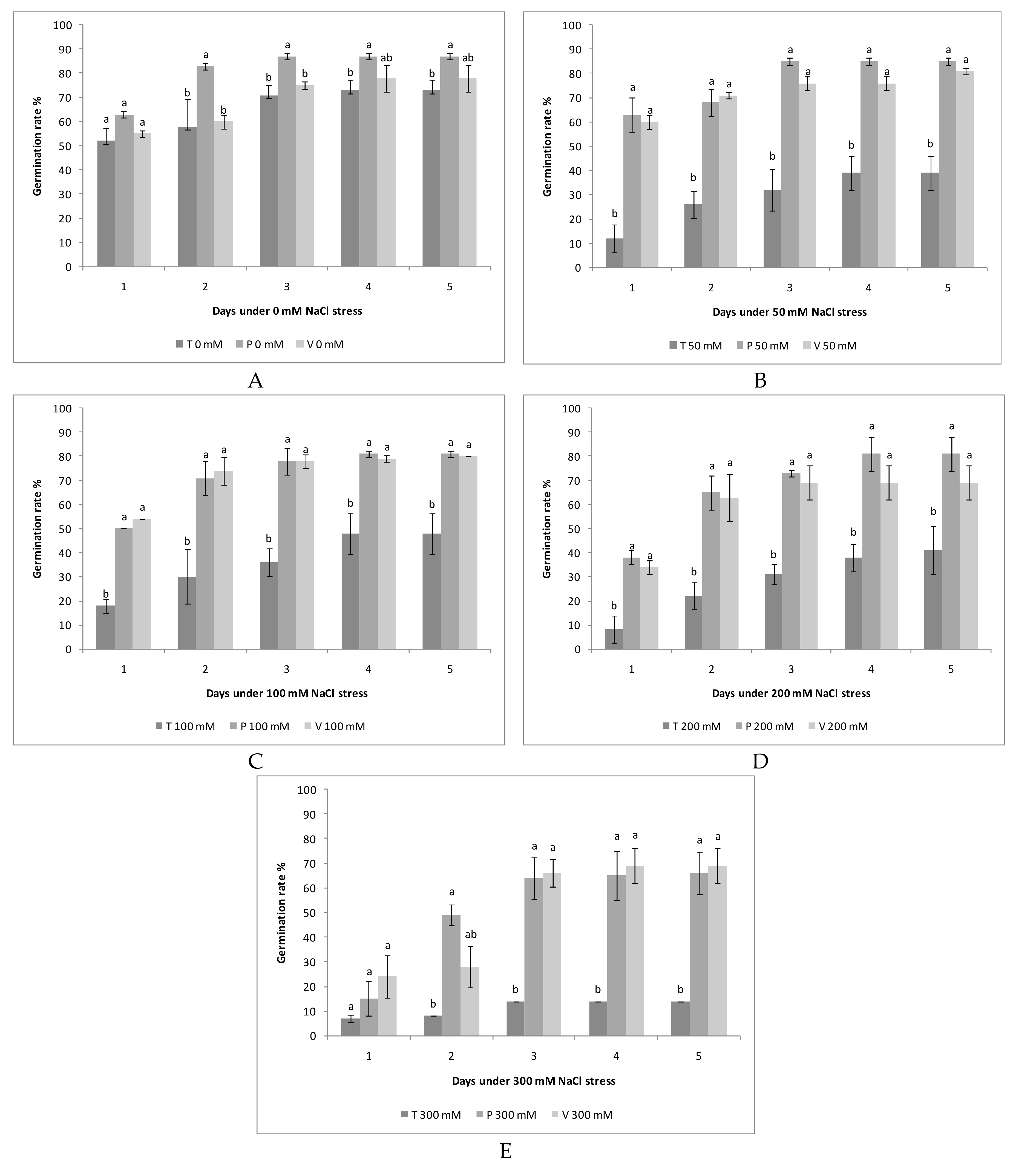
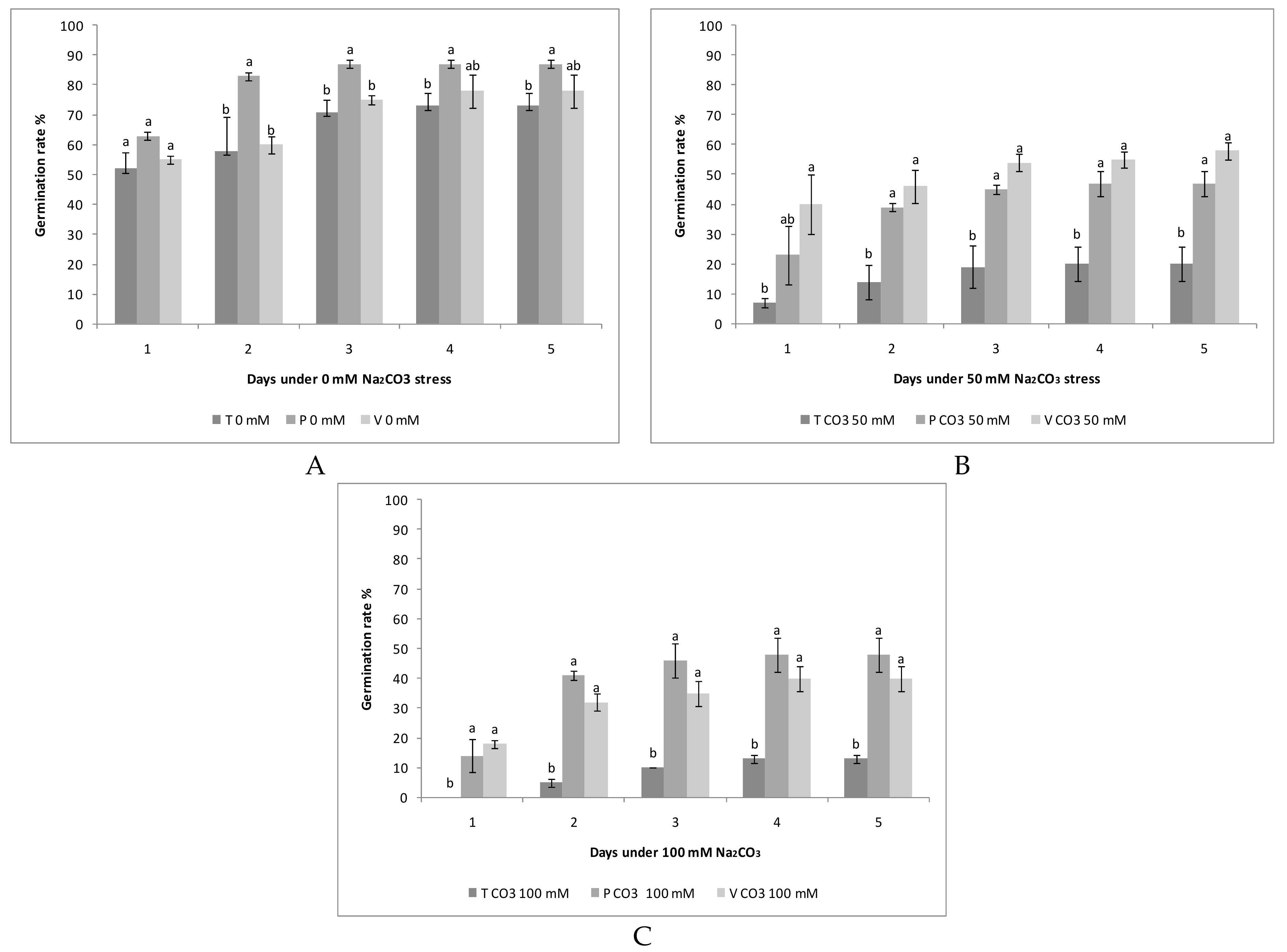
3.1. Salinity and Alkalinity Effect on the Germination of the Seeds
3.2. Tolerance of Quinoa Cultivars to the Salt and Alkali Stress
3.3. Effects of the Salinity and Alkalinity on the Growth of Radicles and Hypocotyls
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jones, A.; Panagos, P.; Barcelo, S.; Bouraoui, F.; Bosco, C.; Dewitte, O.; Gardi, C.; Erhard, M.; Hervás, J.; Hiederer, R.; et al. The State of Soil in Europe: A Contribution of the JRC to the European Environment Agency’s Environment State and Outlook Report—SOER 2010; European Commission, Joint Research Centre, Institute for Environment and Sustainability, Publication Office of the European Union: Luxembourg, 2012. [Google Scholar]
- Gómez-Pando, L.R.; Álvarez-Castro, R.; Eguiluz-de la Barra, A. Short communication: Effect of salt stress on peruvian germplasm of chenopodium quinoa willd.: A promising crop. J. Agron. Crop Sci. 2010, 196, 391–396. [Google Scholar] [CrossRef]
- Grigore, M.; Toma, C.; Zamfirache, M.; Ivanescu, L. A survey of anatomical adaptations in romanian halophytes. Towards an ecological interpretation. Fresenius Environ. Bull. 2012, 21, 3370–3375. [Google Scholar]
- Stolte, J.; Tesfai, M.; Øygarden, L.; Kværnø, S.; Keizer, J.; Verheijen, F.; Panagos, P.; Ballabio, C.; Hessel, R. Soil in Europe—Threats, Functions and Ecosystem Services; JRC Technical Report; Publication Office of the European Union: Luxembourg, 2016. [Google Scholar]
- Koyro, H.-W.; Eisa, S.S. Effect of salinity on composition, viability and germination of seeds of Chenopodium quinoa Willd. Plant Soil 2008, 302, 79–90. [Google Scholar] [CrossRef]
- Ruiz-Carrasco, K.; Antognoni, F.; Coulibaly, A.K.; Lizardi, S.; Covarrubias, A.; Martínez, E.A.; Molina-Montenegro, M.A.; Biondi, S.; Zurita-Silva, A. Variation in salinity tolerance of four lowland genotypes of quinoa (Chenopodium quinoa Willd.) as assessed by growth, physiological traits, and sodium transporter gene expression. Plant Physiol. Biochem. 2011, 49, 1333–1341. [Google Scholar] [CrossRef] [PubMed]
- Moreno, C.; Seal, C.E.; Papenbrock, J. Seed priming improves germination in saline conditions for Chenopodium quinoa and Amaranthus caudatus. J. Agron. Crop Sci. 2018, 204, 40–48. [Google Scholar] [CrossRef]
- Wang, H.; Takano, T.; Liu, S. Screening and Evaluation of Saline–Alkaline Tolerant Germplasm of Rice (Oryza sativa L.) in Soda Saline–Alkali Soil. Agronomy 2018, 8, 205. [Google Scholar] [CrossRef]
- Hu, H.; Liu, H.; Liu, F. Seed germination of hemp (Cannabis sativa L.) cultivars responds differently to the stress of salt type and concentration. Ind. Crops Prod. 2018, 123, 254–261. [Google Scholar] [CrossRef]
- Orsini, F.; Accorsi, M.; Gianquinto, G.; Dinelli, G.; Antognoni, F.; Carrasco, K.B.R.; Martinez, E.A.; Alnayef, M.; Marotti, I.; Bosi, S.; et al. Beyond the ionic and osmotic response to salinity in Chenopodium quinoa: Functional elements of successful halophytism. Funct. Plant Biol. 2011, 38, 818–831. [Google Scholar] [CrossRef]
- Ruiz, K.B.; Biondi, S.; Martínez, E.A.; Orsini, F.; Antognoni, F.; Jacobsen, S.E. Quinoa—A model crop for understanding salt-tolerance mechanisms in halophytes. Plant Biosyst. 2016, 150, 357–371. [Google Scholar] [CrossRef]
- Panuccio, M.R.; Jacobsen, S.E.; Akhtar, S.S.; Muscolo, A. Effect of saline water on seed germination and early seedling growth of the halophyte quinoa. AoB PLANTS 2014, 6, plu047. [Google Scholar] [CrossRef]
- Wu, G.; Peterson, A.J.; Morris, C.F.; Murphy, K.M. Quinoa seed quality response to sodium chloride and sodium sulfate salinity. Front. Plant Sci. 2016, 7, 790. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, S.-E. The scope for adaptation of quinoa in Northern Latitudes of Europe. J. Agron. Crop Sci. 2017, 203, 603–613. [Google Scholar] [CrossRef]
- Hinojosa, L.; González, J.A.; Barrios-Masias, F.H.; Fuentes, F.; Murphy, K.M. Quinoa abiotic stress responses: A review. Plants 2018, 7, 106. [Google Scholar] [CrossRef]
- Adolf, V.I.; Shabala, S.; Andersen, M.N.; Razzaghi, F.; Jacobsen, S.-E. Varietal differences of quinoa’s tolerance to saline conditions. Plant Soil 2012, 357, 117–129. [Google Scholar] [CrossRef]
- Talebnejad, R.; Sepaskhah, A.R. Physiological characteristics, gas exchange, and plant ion relations of quinoa to different saline groundwater depths and water salinity. Arch. Agron. Soil Sci. 2016, 62, 1347–1367. [Google Scholar] [CrossRef]
- Cocozza, C.; Pulvento, C.; Lavini, A.; Riccardi, M.; d’Andria, R.; Tognetti, R. Effects of increasing salinity stress and decreasing water availability on ecophysiological traits of quinoa (Chenopodium quinoa Willd.) grown in a mediterranean-type agroecosystem. J. Agron. Crop Sci. 2013, 199, 229–240. [Google Scholar] [CrossRef]
- Razzaghi, F.; Ahmadi, S.H.; Jacobsen, S.-E.; Jensen, C.R.; Andersen, M.N. Effects of salinity and soil–drying on radiation use efficiency, water productivity and yield of quinoa (Chenopodium quinoa Willd.). J. Agron. Crop Sci. 2012, 198, 173–184. [Google Scholar] [CrossRef]
- Pulvento, C.; Riccardi, M.; Lavini, A.; Iafelice, G.; Marconi, E.; d’Andria, R. Yield and quality characteristics of quinoa grown in open field under different saline and non-saline irrigation regimes. J. Agron. Crop Sci. 2012, 198, 254–263. [Google Scholar] [CrossRef]
- Shabala, L.; Mackay, A.; Tian, Y.; Jacobsen, S.-E.; Zhou, D.; Shabala, S. Oxidative stress protection and stomatal patterning as components of salinity tolerance mechanism in quinoa (Chenopodium quinoa). Physiol. Plant. 2012, 146, 26–38. [Google Scholar] [CrossRef]
- Gómez-Caravaca, A.M.; Iafelice, G.; Lavini, A.; Pulvento, C.; Caboni, M.F.; Marconi, E. Phenolic compounds and saponins in quinoa samples (Chenopodium quinoa Willd.) grown under different saline and nonsaline irrigation regimens. J. Agric. Food Chem. 2012, 60, 4620–4627. [Google Scholar] [CrossRef]
- Hariadi, Y.; Marandon, K.; Tian, Y.; Jacobsen, S.-E.; Shabala, S. Ionic and osmotic relations in quinoa (Chenopodium quinoa Willd.) plants grown at various salinity levels. J. Exp. Bot. 2011, 62, 185–193. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Carillo, P.; Gibbon, Y. PROTOCOL: Extraction and determination of proline. PrometheusWiki. 2011. Available online: http://www.researchgate.net/publication/211353600_PROTOCOL_Extraction_and_determination_of_proline (accessed on 3 June 2019).
- Brakez, M.; Harrouni, M.C.; Tachbibi, N.; Daoud, S. Comparative effect of NaCl and seawater on germination of quinoa seed (Chenopodium quinoa Willd). Emir. J. Food Agric. 2014, 26, 1091–1096. [Google Scholar] [CrossRef]
- Shrivastava, P.; Kumar, R. Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi J. Biol. Sci. 2015, 22, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Peterson, A.; Murphy, K. Tolerance of lowland quinoa cultivars to sodium chloride and sodium sulfate salinity. Crop Sci. 2015, 55, 331–338. [Google Scholar] [CrossRef]
- Shahid, S.A.; Rahman, K. Soil salinity development, classification, assessment and management in irrigated agriculture. In Handbook of Plant and Crop Stress, 3rd ed.; Pessarakli, M., Ed.; CRC Press: Boca Raton, FL, USA, 2010; pp. 23–41. [Google Scholar]
- Grattan, S.R.; Grieve, C.M. Salinity–mineral nutrient relations in horticultural crops. Sci. Hortic. 1998, 78, 127–157. [Google Scholar] [CrossRef]
- Shabala, S.; Hariadi, Y.; Jacobsen, S.-E. Genotypic difference in salinity tolerance in quinoa is determined by differential control of xylem Na+ loading and stomatal density. J. Plant Physiol. 2013, 170, 906–914. [Google Scholar] [CrossRef]
- González, J.A.; Prado, F.E. Germination in relation to salinity and temperature in Chenopodium quinoa (Willd.). Agrochimica 1992, 36, 101–107. [Google Scholar]
- Ruffino, A.M.C.; Rosa, M.; Hilal, M.; González, J.A.; Prado, F.E. The role of cotyledon metabolism in the establishment of quinoa (Chenopodium quinoa) seedlings growing under salinity. Plant Soil 2010, 326, 213–224. [Google Scholar] [CrossRef]
- Prado, F.E.; Boero, C.; Gallardo, M.; González, J.A. Effect of NaCl on germination, growth, and soluble sugar content in Chenopodium quinoa Willd. seeds. Bot. Bull. Acad. Sin. 2000, 41, 27–34. [Google Scholar]
- Gupta, B.; Huang, B. Mechanism of salinity tolerance in plants: Physiological, biochemical, and molecular characterization. Int. J. Genom. 2014, 2014, 18. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, M.; Li, J.; Yahya, M.; Wang, M.; Ali, A.; Cheng, A.; Wang, X.; Ma, C. Grain Legumes and Fear of Salt Stress: Focus on Mechanisms and Management Strategies. Int. J. Mol. Sci. 2019, 20, 799. [Google Scholar] [CrossRef] [PubMed]
- Adolf, V.I.; Jacobsen, S.-E.; Shabala, S. Salt tolerance mechanisms in quinoa (Chenopodium quinoa Willd.). Environ. Exp. Bot. 2013, 92, 43–54. [Google Scholar] [CrossRef]
- Hooks, T.; Niu, G. Relative Salt Tolerance of Four Herbaceous Perennial Ornamentals. Horticulturae 2019, 5, 36. [Google Scholar] [CrossRef]
- Orlovsky, N.; Japakova, U.; Zhang, H.; Volis, S. Effect of salinity on seed germination, growth and ion content in dimorphic seeds of Salicornia europaea L. (Chenopodiaceae). Plant Divers. 2016, 38, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Delatorre-Herrera, J.; Pinto, M. Importance of ionic and osmotic components of salt stress on the germination of four quinua (Chenopodium quinoa Willd.) selections. Chil. J. Agric. Res. 2009, 69, 477–485. [Google Scholar] [CrossRef]
- Li, Y. Effect of salt stress on seed germination and seedling growth of three salinity plants. Pak. J. Biol. Sci. 2008, 11, 1268–1272. [Google Scholar] [CrossRef]
- Demir, I.; Mavi, K.M. Effect of salt and osmotic stresses on the germination of pepper seeds of different maturation stages. Braz. Arch. Biol. Technol. 2008, 51, 897–902. [Google Scholar] [CrossRef] [Green Version]
- Wierzbicka, M.; Obidzińska, J. The effect of lead on seed imbibition and germination in different plant species. Plant Sci. 1998, 137, 155–171. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Salt | Concentration (mM) | Proline Content (nmol mg−1 FW) | ||
|---|---|---|---|---|
| Titicaca | Puno | Vikinga | ||
| NaCl | 0 | 0.59 a | 0.81 a | 0.47 c |
| 50 | 0.38 a | 0.8 a | 0.56 bc | |
| 100 | 0.96 a | 1.14 a | 0.77 abc | |
| 200 | 0.97 a | 1.14 a | 1.02 ab | |
| 300 | 0.97 a | 1.06 a | 1.19 a | |
| Na2SO4 | 0 | 0.59 a | 0.81 a | 0.47 a |
| 50 | 0.61 a | 0.46 a | 0.99 a | |
| 100 | 1.11 a | 0.43 a | 0.91 a | |
| 200 | 0.63 a | 0.55 a | 0.54 a | |
| 300 | 0.91 a | 1.01 a | 0.81 a | |
| Na2CO3 | 0 | 0.59 a | 0.81 a | 0.47 a |
| 50 | 0.81 a | 0.49 a | 0.38 a | |
| 100 | 0.47 a | 0.57 a | 0.55 a | |
| 200 | 0.48 a | 0.74 a | 0.56 a | |
| 300 | 0.51 a | 0.98 a | 0.50 a | |
| Treatment | Titicaca | Puno | Vikinga | ||||
|---|---|---|---|---|---|---|---|
| Salt | Concentration (mM) | Radicle Length (mm) | Hypocotyls Length (mm) | Radicle Length (mm) | Hypocotyls Length (mm) | Radicle Length (mm) | Hypocotyls Length (mm) |
| NaCl | 0 50 100 200 300 | 8.1 ab 7.2 ab 10.4 a 6.1 ab 4.9 b | 23.3 a 18.1 b 9.5 c 2.7 d 0 d | 12.8 a 7.4 b 7.4 b 5.5 bc 4.8 c | 27.2 a 20.2 b 16.8 bc 10.4 c 0 d | 20.5 a 12.6 bc 18.5 ab 11.8 c 6.8 c | 31.8 a 22.8 ab 17.1 b 10.7 bc 5.8 c |
| Na2SO4 | 0 50 100 200 300 | 8.1 a 6.1 a 5.2 ab 2.9 bc 1.6 c | 23.3 a 11.6 b 8.5 bc 0 c 0 c | 12.8 a 7.8 b 5.9 bc 4.6 bc 3.5 c | 27.2 a 15.8 b 9.45 b 0 c 0 c | 20.5 a 15 ab 8.2 bc 5.1 c 2.6 c | 31.75 a 9.9 bc 11.8 b 2.4 cd 0 d |
| Na2CO3 | 0 50 100 200 300 | 8.1 a 2.2 b 1.5 b 0 c 0 c | 23.3 a 0 b 0 b 0 b 0 b | 12.8 a 3.1 b 2 b 0 c 0 c | 27.2 a 0 b 0 b 0 b 0 b | 20.5 a 8.4 b 2.7 c 0 c 0 c | 31.8 a 8.9 b 3.9 c 0 d 0 d |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stoleru, V.; Slabu, C.; Vitanescu, M.; Peres, C.; Cojocaru, A.; Covasa, M.; Mihalache, G. Tolerance of Three Quinoa Cultivars (Chenopodium quinoa Willd.) to Salinity and Alkalinity Stress During Germination Stage. Agronomy 2019, 9, 287. https://doi.org/10.3390/agronomy9060287
Stoleru V, Slabu C, Vitanescu M, Peres C, Cojocaru A, Covasa M, Mihalache G. Tolerance of Three Quinoa Cultivars (Chenopodium quinoa Willd.) to Salinity and Alkalinity Stress During Germination Stage. Agronomy. 2019; 9(6):287. https://doi.org/10.3390/agronomy9060287
Chicago/Turabian StyleStoleru, Vasile, Cristina Slabu, Maricel Vitanescu, Catalina Peres, Alexandru Cojocaru, Mihaela Covasa, and Gabriela Mihalache. 2019. "Tolerance of Three Quinoa Cultivars (Chenopodium quinoa Willd.) to Salinity and Alkalinity Stress During Germination Stage" Agronomy 9, no. 6: 287. https://doi.org/10.3390/agronomy9060287
APA StyleStoleru, V., Slabu, C., Vitanescu, M., Peres, C., Cojocaru, A., Covasa, M., & Mihalache, G. (2019). Tolerance of Three Quinoa Cultivars (Chenopodium quinoa Willd.) to Salinity and Alkalinity Stress During Germination Stage. Agronomy, 9(6), 287. https://doi.org/10.3390/agronomy9060287