Modelling Crop Transpiration in Greenhouses: Different Models for Different Applications



Abstract
:1. Introduction
2. The Most Common Approaches
2.1. The Penman–Monteith Model
2.2. Simplified Models
2.3. Single and Multilayer Models
2.4. The Sub-Models
2.4.1. Aerodynamic Conductance
2.4.2. Stomatal Conductance
2.4.3. Net Radiation of a Greenhouse Crop
3. Transpiration from External Climate
4. Different Models for Different Applications
Are All Sub-Models Equally Necessary?
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Abdessalam, O.A. Preface. In Good Agricultural Practices for Greenhouse Vegetable Crops: Principles for Mediterranean Climate Areas; FAO Plant Production and Protection Paper 217; Baudoin, W., Nono-Womdim, R., Lutaladio, N., Hodder, A., Castilla, N., Leonardi, C., De Pascale, S., Quaryouti, M., Eds.; FAO: Rome, Italy, 2013. [Google Scholar]
- Fernández, M.D.; Gallardo, M.; Bonachela, S.; Orgaz, F.; Thompson, R.B.; Fereres, E. Water use and production of a greenhouse pepper crop under optimum and limited water supply. J. Hortic. Sci. Biotechnol. 2005, 80, 87–96. [Google Scholar] [CrossRef]
- Stanghellini, C. Horticultural production in greenhouses: Efficient use of water. Acta Hortic. 2014, 1034, 25–32. [Google Scholar] [CrossRef]
- Stanghellini, C. Transpiration of Greenhouse Crops an Aid to Climate Management. Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, 1987. [Google Scholar]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop evapotranspiration. In Guidelines for Computing Crop Water Requirements; FAO Irrigation and Drainage Paper 56; FAO: Rome, Italy, 1998. [Google Scholar]
- Kittas, C.; Katsoulas, N.; Baille, A. Transpiration and canopy resistance of greenhouse soilless roses in Greece. Measurements and modeling. Acta Hortic. 1999, 507, 61–68. [Google Scholar] [CrossRef]
- Yang, X.; Short, T.H.; Robert, D.F.; Bauerle, W.L. Transpiration, leaf temperature and stomatal resistance of a cucumber crop. Agric. For. Meteorol. 1990, 51, 197–209. [Google Scholar] [CrossRef]
- Katsoulas, N.; Kittas, C.; Baille, A. Estimating transpiration rate and canopy resistance of a rose crop in a fan-ventilated greenhouse. Acta Hortic. 1999, 548, 303–309. [Google Scholar] [CrossRef]
- Tarara, J.M. Microclimate modification with plastic mulch. HortScience 2000, 35, 169–180. [Google Scholar] [CrossRef]
- Seginer, I.; Kantz, D. In-situ determination of transfer coefficients for heat and water vapour in a small greenhouse. J. Agric. Eng. Res. 1986, 35, 39–54. [Google Scholar] [CrossRef]
- Yang, X. Comments on ‘Thermal and aerodynamic conditions in greenhouses in relation to estimation of heat flux and evapotranspiration’. Agric. For. Meteorol. 1995, 77, 131–136. [Google Scholar] [CrossRef]
- Leonardi, C.; Baille, A.; Guichard, S. Predicting transpiration of shaded and non-shaded tomato fruits under greenhouse environments. Sci. Hortic. 2000, 84, 297–307. [Google Scholar] [CrossRef]
- Boulard, T.; Fatnassi, H.; Roy, J.C.; Lagier, J.; Fargues, J.; Smits, N.; Rougier, M.; Jeannequin, B. Effect of greenhouse ventilation on humidity of inside air and in leaf boundary-layer. Agric. For. Meteorol. 2004, 125, 225–239. [Google Scholar] [CrossRef]
- Penman, H.L. Natural evaporation from open water, bare soil and grass. Proc. R. Soc. Lond. A 1948, 193, 120–145. [Google Scholar] [CrossRef] [Green Version]
- Monteith, J.L. Evaporation and environment. In Symposia of the Society for Experimental Biology; Cambridge University Press (CUP): Cambridge, UK, 1965; Volume 19, pp. 205–234. [Google Scholar]
- Morris, L.G.; Neale, F.E.; Postlethwaite, J.D. The transpiration of glasshouse crops and its relationship to the incoming solar radiation. J. Agric. Eng. Res. 1957, 2, 111–122. [Google Scholar]
- De Villèle, O. Besoins en eau des cultures sous serres—Essai de conduite des arrosages en fonction de l’ensoleillement. Acta Hortic. 1972, 35, 123–135. [Google Scholar]
- Stanhill, G.; Scholte Albers, J. Solar radiation and water loss from glasshouse roses. J. Amer. Soc. Hortic. Sci. 1974, 99, 107–110. [Google Scholar]
- Van der Post, C.J.; van Schie, J.J.; de Graaf, R. Energy balance and water supply in glasshouses in the West-Netherlands. Acta Hortic. 1974, 35, 13–22. [Google Scholar] [CrossRef]
- De Graaf, R. Influence of moisture deficit leaf-air and cultural practices on transpiration of glasshouse roses. Acta Hortic. 1995, 401, 545–552. [Google Scholar] [CrossRef]
- Jones, H.G.; Tardieu, F. Modelling water relations of horticultural crops: A review. Sci. Hortic. 1998, 74, 21–46. [Google Scholar] [CrossRef]
- Bakker, J.C. Leaf conductance of four glasshouse vegetable crops as affected by air humidity. Agric. For. Meteorol. 1991, 55, 23–36. [Google Scholar] [CrossRef]
- Baille, M.; Baille, A.; Delmon, D. Microclimate and transpiration of greenhouse rose crops. Agric. For. Meteorol. 1994, 71, 83–97. [Google Scholar] [CrossRef]
- Katsoulas, N.; Baille, A.; Kittas, C. Effect of air misting on transpiration and bulk conductances of a greenhouse rose canopy. Agric. For. Meteorol. 2001, 106, 233–247. [Google Scholar] [CrossRef]
- Kittas, C.; Katsoulas, N.; Tchamitchian, M. Greenhouse Soilless Rose Crop Transpiration. Application for Accurate Irrigation Control. In Proceedings of the VII Conference on Protection and Restoration of the Environment, Mykonos, Greece, 28 June–1 July 2004; pp. 998–1005. Available online: http://pre14.civil.auth.gr/index.php/previous-conferences/previous-conferences (accessed on 7 June 2019).
- Kittas, C.; Baille, A.; Katsoulas, N. Influence of misting on the diurnal hysteresis of canopy transpiration rate and conductance in a rose greenhouse. Acta Hortic. 1999, 534, 185–193. [Google Scholar] [CrossRef]
- Seginer, I. Transpirational cooling of a greenhouse crop with partial ground cover. Agric. For. Meteorol. 1994, 71, 265–281. [Google Scholar] [CrossRef]
- Seginer, I.; Tarnopolsky, M. Response of plants to ventilation regimes. In Transpirational Cooling of Greenhouse Crops; Chapter T3, Final Report, BARD Project IS-2538-95R; Seginer, I., Willits, D.H., Raviv, M., Peet, M.M., Eds.; Agricultural Research Organization of Israel: Bet Dagan, Israel, 2000; pp. 165–171. [Google Scholar]
- Seginer, I. Alternative design formulae for the ventilation rate of greenhouses. J. Agric. Eng. Res. 1997, 68, 355–365. [Google Scholar] [CrossRef]
- Carmassi, G.; Bacci, L.; Bronzini, M.; Incrocci, L.; Maggini, R.; Bellocchi, G.; Massa, D.; Pardossi, A. Modelling transpiration of greenhouse gerbera (Gerbera jamesonii H. Bolus) grown in substrate with saline water in a Mediterranean climate. Sci. Hortic. 2013, 156, 9–18. [Google Scholar] [CrossRef]
- Seginer, I. The Penman–Monteith Evapotranspiration Equation as an Element in Greenhouse Ventilation Design. Biosyst. Eng. 2002, 82, 423–439. [Google Scholar] [CrossRef]
- Katsoulas, N.; Sapounas, A.; de Zwart, F.; Dieleman, J.A.; Stanghellini, C. Reducing ventilation requirements in semi-closed greenhouses increases water use efficiency. Agric. Water. Man. 2015, 156, 90–99. [Google Scholar] [CrossRef]
- Jemaa, R. Mise AU Point ET Validation de Modèles DE Transpiration de Cultures de Tomate Hors SOL Sous Serre. Application à La Conduite DE La Fert-Irrigation. Ph.D. Thesis, Brittany National College of Architecture, Rennes, France, 1995; p. 111. [Google Scholar]
- Boulard, T.; Baille, A.; Le Gall, F. Etude de différentes méthodes de refroidissement sur le climat et la transpiration de tomates de serre. Agronomie 1991, 11, 543–554. [Google Scholar] [CrossRef]
- Papadakis, G.; Frangoudakis, A.; Kyritsis, S. Experimental investigation and modelling of heat mass transfer between a tomato crop and greenhouse environment. J. Agric. Eng. Res. 1994, 57, 217–227. [Google Scholar] [CrossRef]
- González-Real, M.M. Estudio y Modelizacion de Intercambios Gaseosos en Cultivos de Rosas Bajo Invernadero. Ph.D. Τhesis, Universidad Politecnica de Valencia, Valencia, Spain, 1995; p. 192. [Google Scholar]
- Stanghellini, C. Response to comments on ‘Thermal and aerodynamic conditions in greenhouses in relation to estimation of heat flux and evapotranspiration’. Agric. For. Meteorol. 1995, 77, 137–138. [Google Scholar] [CrossRef]
- Raupach, M.R.; Finnigan, J.J. Single-layer models of evaporation from plant canopies are incorrect but useful, whereas mutlilayer models are correct but useless: Discuss. Aust. J. Plant Physiol. 1988, 15, 705–716. [Google Scholar]
- Yang, X.; Short, T.H.; Fox, R.D.; Bauerle, W.L. The microclimate and transpiration of a greenhouse cucumber crop. Am. Soc. Agric. Eng. 1989, 32, 2143–2150. [Google Scholar] [CrossRef]
- Kittas, C.; Katsoulas, N.; Baille, A. Influence of greenhouse ventilation regime on microclimate and energy partitioning of a rose canopy during summer conditions. J. Agric. Eng. Res. 2001, 79, 349–360. [Google Scholar] [CrossRef]
- Kittas, C.; Katsoulas, N.; Baille, A. Influence of an aluminised thermal screen on greenhouse microclimate and canopy energy balance. Trans. ASAE 2001, 46, 1653–1663. [Google Scholar] [CrossRef]
- Bojaca, C.R.; Gil, R.; Cooman, A. Use of geostatistical and crop growth modelling to assess the variability of greenhouse tomato yield caused by spatial temperature variations. Comp. Electr. Agric. 2009, 65, 219–227. [Google Scholar] [CrossRef]
- Morille, B.; Migeon, C.; Bournet, P. Is the Penman–Monteith model adapted to predict crop transpiration under greenhouse conditions? Application to a New Guinea Impatiens crop. Sci. Hortic. 2013, 152, 80–91. [Google Scholar] [CrossRef]
- Stanghellini, C.; Dieleman, J.A.; Driever, S.M.; Marcelis, L.F.M. Modelling the effect of the position of cooling elements on the vertical profile of transpiration in a greenhouse tomato crop. Acta Hortic. 2012, 952, 763–769. [Google Scholar] [CrossRef]
- Kreith, F. Principles of Heat Transfer, 3rd ed.; Intext Educational Publishers: New York, NY, USA, 1973; p. 656. [Google Scholar]
- Monteith, J.L.; Unsworth, M.H. Principles of Environmental Physics, 4th ed.; Academic Press: Cambridge, MA, USA, 2014; p. 401. [Google Scholar]
- Schuepp, P.H. Leaf boundary layers. New Phytol. 1993, 125, 477–507. [Google Scholar] [CrossRef]
- Montero, J.I.; Antón, A.; Muñoz, P.; Lorenzo, P. Transpiration from geranium grown under high temperatures and low humidities in greenhouses. Agric. For. Meteorol. 2001, 107, 323–332. [Google Scholar] [CrossRef]
- Katsoulas, N.; Baille, A.; Kittas, C. Leaf boundary layer conductance in ventilated greenhouses. An experimental approach. Agric. For. Meteorol. 2007, 144, 180–192. [Google Scholar] [CrossRef]
- Bailey, B.J.; Montero, J.I.; Biel, C.; Wilkinson, D.J.; Anton, A.; Jolliet, O. Transpiration of Ficus benjamina: Comparison of measurements with predictions of the Penman-Monteith model and a simplified version. Agric. For. Meteorol. 1993, 65, 229–243. [Google Scholar] [CrossRef]
- Kitano, M.; Eguchi, H. Buoyancy effect on forced convection in the leaf boundary layer. Plant Cell Env. 1990, 13, 965–970. [Google Scholar] [CrossRef]
- Zhang, L.; Lemeur, R. Effect of aerodynamic resistance on energy balance and Penman-Monteith estimates of evapotranspiration in greenhouse conditions. Agric. For. Meteorol. 1992, 58, 209–228. [Google Scholar] [CrossRef]
- Seginer, I. On the night transpiration of greenhouse roses under glass or plastic cover. Agric. For. Meteorol. 1984, 30, 257–268. [Google Scholar] [CrossRef]
- Stanghellini, C. Mixed convection above greenhouse crop canopies. Agric. For. Meteorol. 1993, 66, 111–117. [Google Scholar] [CrossRef]
- Stanghellini, C.; de Jong, T. A model of humidity and its applications in a greenhouse. Agric. For. Meteorol. 1995, 76, 129–148. [Google Scholar] [CrossRef]
- Cowan, I.R. Regulation of water use in relation to carbon gain in higher plants. In Physiological Plant Ecology II; Lange, O.L., Novel, P.S., Osmond, C.B., Zeigler, H., Eds.; Springer: Berlin, Germany, 1982; pp. 589–614. [Google Scholar]
- Jarvis, P.G. The interpretation of the variations in leaf water potential and stomatal conductance found in canopies in the field. Philos. Trans. R. Soc. Lond. B 1976, 273, 593–610. [Google Scholar] [CrossRef]
- Jolliet, O.; Bailey, B.J. The effect of climate on tomato transpiration in greenhouses: Measurements and models comparison. Agric. For. Meteorol. 1992, 58, 43–62. [Google Scholar] [CrossRef]
- Fuchs, M.; Stanghellini, C. The functional dependence of canopy conductance on water vapour pressure deficit revisited. Int. J. Biometeorol. 2018, 62, 1211–1220. [Google Scholar] [CrossRef]
- Monsi, M.; Saeki, T. Uber den Lichtfaktor in den Pflanzengesellschaften und seine Bedeutung für die Stoffproduktion. Jpn. J. Bot. 1953, 14, 22–25. [Google Scholar]
- Ross, J. Radiative transfer in plant communities. In Vegetation and the Atmosphere; Monteith, J.L., Ed.; Academic Press: London, UK, 1975; pp. 13–52. [Google Scholar]
- Goudriaan, J. Crop Micrometeorology: A Simulation Study, Simulation Monographs; Pudoc: Wageningen, The Netherlands, 1977; p. 249. [Google Scholar]
- Bontsema, J.; Hemming, J.; Stanghellini, C.; De Visser, P.; Van Henten, E.J.; Budding, J.; Rieswijk, T.; Nieboer, S. On-line estimation of the transpiration in greenhouse horticulture. In Proceedings of the Agricontrol 2007, 2nd IFAC International Conference on Modeling and Design of Control Systems in Agriculture, Osijek, Croatia, 3–5 September 2007; pp. 29–34. [Google Scholar]
- Aubinet, M.; Deltour, J.; de Halleux, D.; Nijskens, J. Stomatal regulation in greenhouse crops: Analysis and simulation. Agric. For. Meteorol. 1989, 48, 21–44. [Google Scholar] [CrossRef]
- Boulard, T.; Wang, S. Greenhouse crop transpiration simulation from external climate conditions. Agric. For. Meteorol. 2000, 100, 25–34. [Google Scholar] [CrossRef]
- Jolliet, O. HORTITRANS, a model for predicting and optimizing humidity and transpiration in greenhouses. J. Agric. Eng. Res. 1994, 57, 23–37. [Google Scholar] [CrossRef]
- Kirnak, H.; Hansen, R.; Keener, H.; Short, T. An evaluation of physically based and empirically determined evapotranspiration models for nursery plants. Turk. J. Agric. For. 2002, 26, 355–362. [Google Scholar]
- Lopez-Cruz, I.; Olivera-Lopez, M.; Herrera-Ruiz, G. Simulation of greenhouse tomato crop transpiration by two theoretical models. Acta Hortic. 2008, 797, 145–150. [Google Scholar] [CrossRef]
- Prenger, J.J.; Fynn, R.P.; Hansen, R.C. A comparison of four evapotranspiration models in a greenhouse environment. Trans. ASAE 2002, 45, 1779–1788. [Google Scholar] [CrossRef]
- Villarreal-Guerrero, F.; Kacira, M.; Fitz-Rodríguez, E.; Linker, R.; Kubota, C.; Giacomelli, G.A.; Arbel, A. Simulated performance of a greenhouse cooling control strategy with natural ventilation and fog cooling. Biosyst. Eng. 2012, 111, 217–228. [Google Scholar] [CrossRef]
- Acquah, S.J.; Yan, H.; Zhang, C.; Wang, G.; Zhao, B.; Wu, H.; Zhang, H. Application and evaluation of Stanghellini model in the determination of crop evapotranspiration in a naturally ventilated greenhouse. Int. J. Agric. Biol. Eng. 2018, 11, 95–103. [Google Scholar]



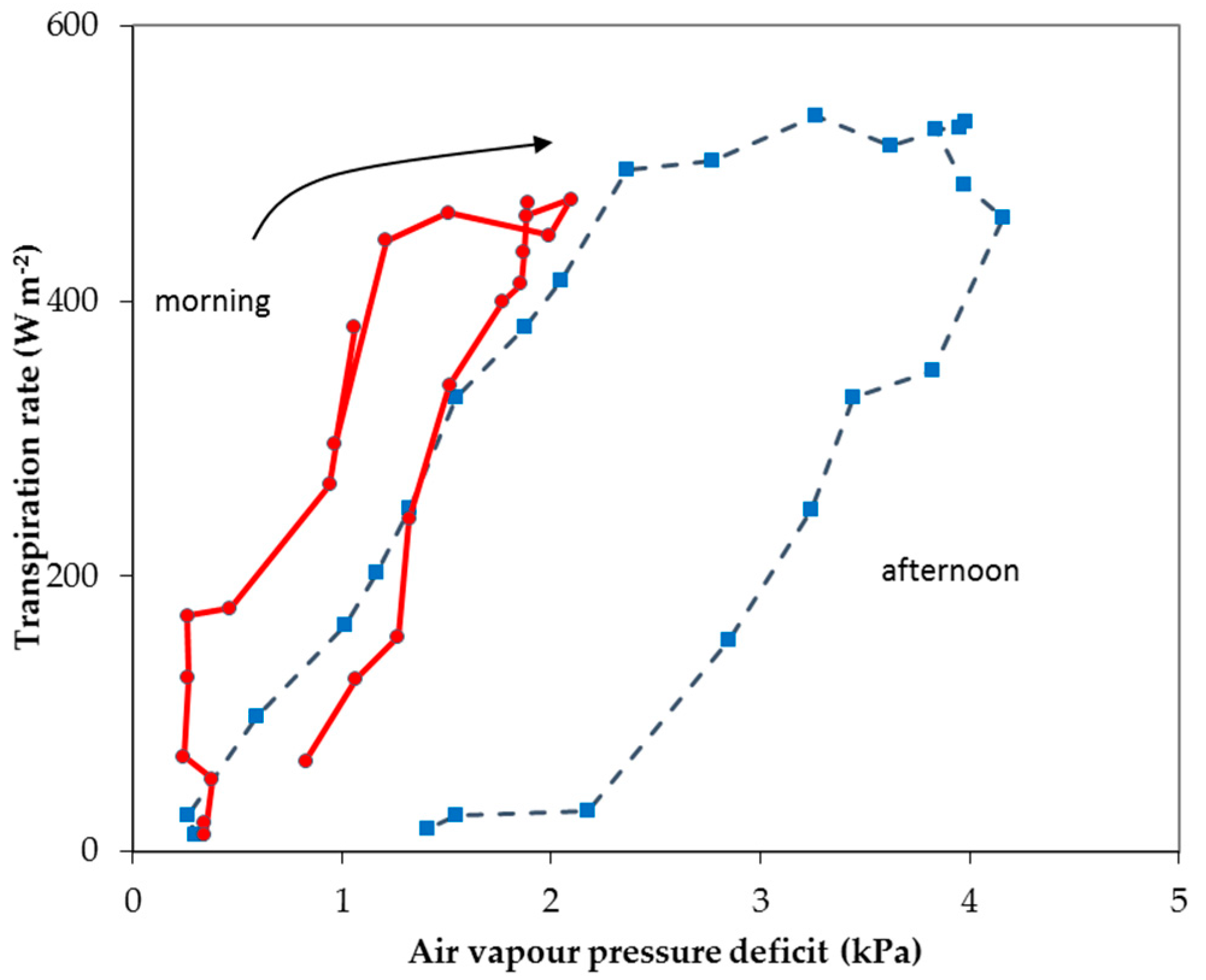{kind=link}
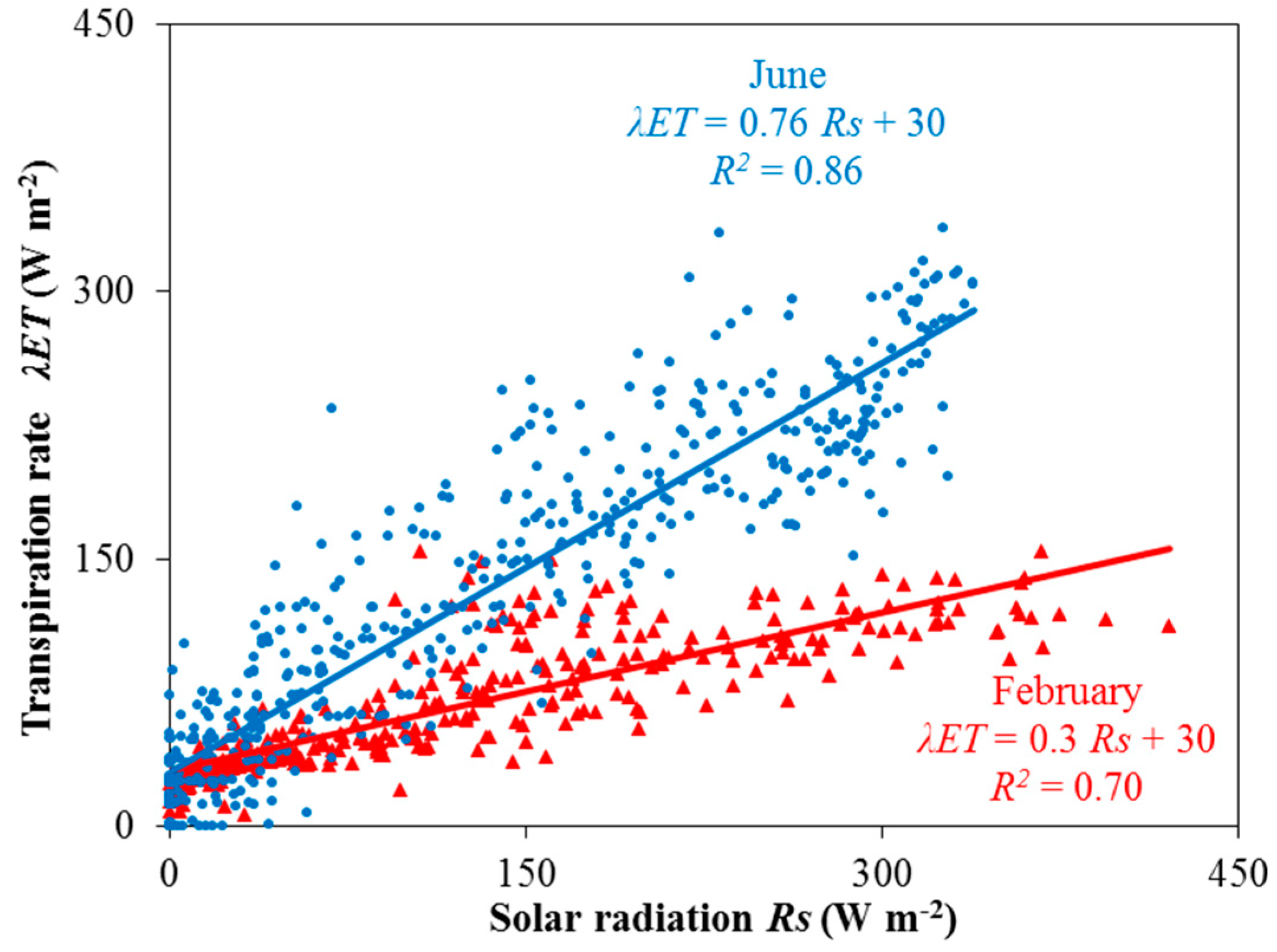{kind=link}
| f1(Rs) | f2(Di) | f3(Ti) | f4(CO2) | Reference |
|---|---|---|---|---|
| (0.54 + Rs)/(4.3 + Rs)] | 1/[1 + 4.3 Di 2] | 1/[1 + 0.023(Ti − 24.5)2] | 1/[1 + 6.110−7 (CO2 − 200)2] | [4] |
| 1/(0.6 + Rs) | 1/Di 55.1 | 1/(2.1 + Tc)40.8 | - | [35] |
| 1/{[1 + 1/[exp(0.005(Rs − 50))]} | 1/{1 + 0.11exp[0.34(Di/100) − 10]} | - | - | [34] |
| Conductance | Yearly Mean (mm/s) | Min (mm/s) | When |
|---|---|---|---|
| Max (mm/s) | |||
| gt | 2–4 (crop-dependent) | 0.02 | Small crops, night-time |
| 5–10 (crop-dependent) | Large crops, sunny afternoon | ||
| gcd | 2–3 (single cover) | 0 | Well-insulated cover |
| 5 | Very cold cover | ||
| gv | 3–4 | 0.03 | Closed windows and well tight |
| 50 | Fully open windows, windy |
| Author | Crop | Model | Slope | R2 |
|---|---|---|---|---|
| [67] | Red Sunset Maple (Summer) | Penman | 0.96 | 0.58 |
| Penman–Monteith | 1.03 | 0.70 | ||
| Stanghellini | 0.80 | 0.65 | ||
| Fynn | 0.73 | 0.64 | ||
| [68] | Tomato (Spring) | Penman–Monteith | Unknown | 0.62 |
| Stanghellini | Unknown | 0.72 | ||
| [69] | Red Sunset Maple (Summer) | Penman | Unknown | 0.21 |
| Penman–Monteith | Unknown | 0.48 | ||
| Stanghellini | Unknown | 0.87 | ||
| Fynn | Unknown | −0.85 | ||
| [70] | Bell pepper (Summer + natural ventilation + fog) | Stanghellini | 0.92 | 0.88 |
| Energy balanced equation | 0.94 | 0.90 | ||
| Penman–Monteith | 1.28 | 0.95 | ||
| Bell pepper (Summer + pad and fan) | Stanghellini | 0.84 | 0.96 | |
| Energy balanced equation | 1.01 | 0.89 | ||
| Penman–Monteith | 1.25 | 0.96 | ||
| Tomato (Fall + pad and fan) | Stanghellini | 0.98 | 0.72 | |
| Energy balanced equation | 0.85 | 0.66 | ||
| Penman–Monteith | 0.94 | 0.51 | ||
| Tomato (Spring + pad and fan) | Stanghellini | 0.96 | 0.93 | |
| Energy balanced equation | 1.04 | 0.86 | ||
| Penman–Monteith | 1.04 | 0.90 | ||
| Tomato (Spring + natural ventilation) | Stanghellini | 1.02 | 0.95 | |
| Energy balanced equation | 0.96 | 0.88 | ||
| Penman–Monteith | 1.14 | 0.94 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Katsoulas, N.; Stanghellini, C. Modelling Crop Transpiration in Greenhouses: Different Models for Different Applications. Agronomy 2019, 9, 392. https://doi.org/10.3390/agronomy9070392
Katsoulas N, Stanghellini C. Modelling Crop Transpiration in Greenhouses: Different Models for Different Applications. Agronomy. 2019; 9(7):392. https://doi.org/10.3390/agronomy9070392
Chicago/Turabian StyleKatsoulas, Nikolaos, and Cecilia Stanghellini. 2019. "Modelling Crop Transpiration in Greenhouses: Different Models for Different Applications" Agronomy 9, no. 7: 392. https://doi.org/10.3390/agronomy9070392
APA StyleKatsoulas, N., & Stanghellini, C. (2019). Modelling Crop Transpiration in Greenhouses: Different Models for Different Applications. Agronomy, 9(7), 392. https://doi.org/10.3390/agronomy9070392