Genetic Diversity and Geographic Differentiation of Tung Tree, Vernicia Fordii (Euphorbiaceae), A Potential Biodiesel Plant Species with Low Invasion Risk



Abstract
:1. Introduction
2. Materials and Methods
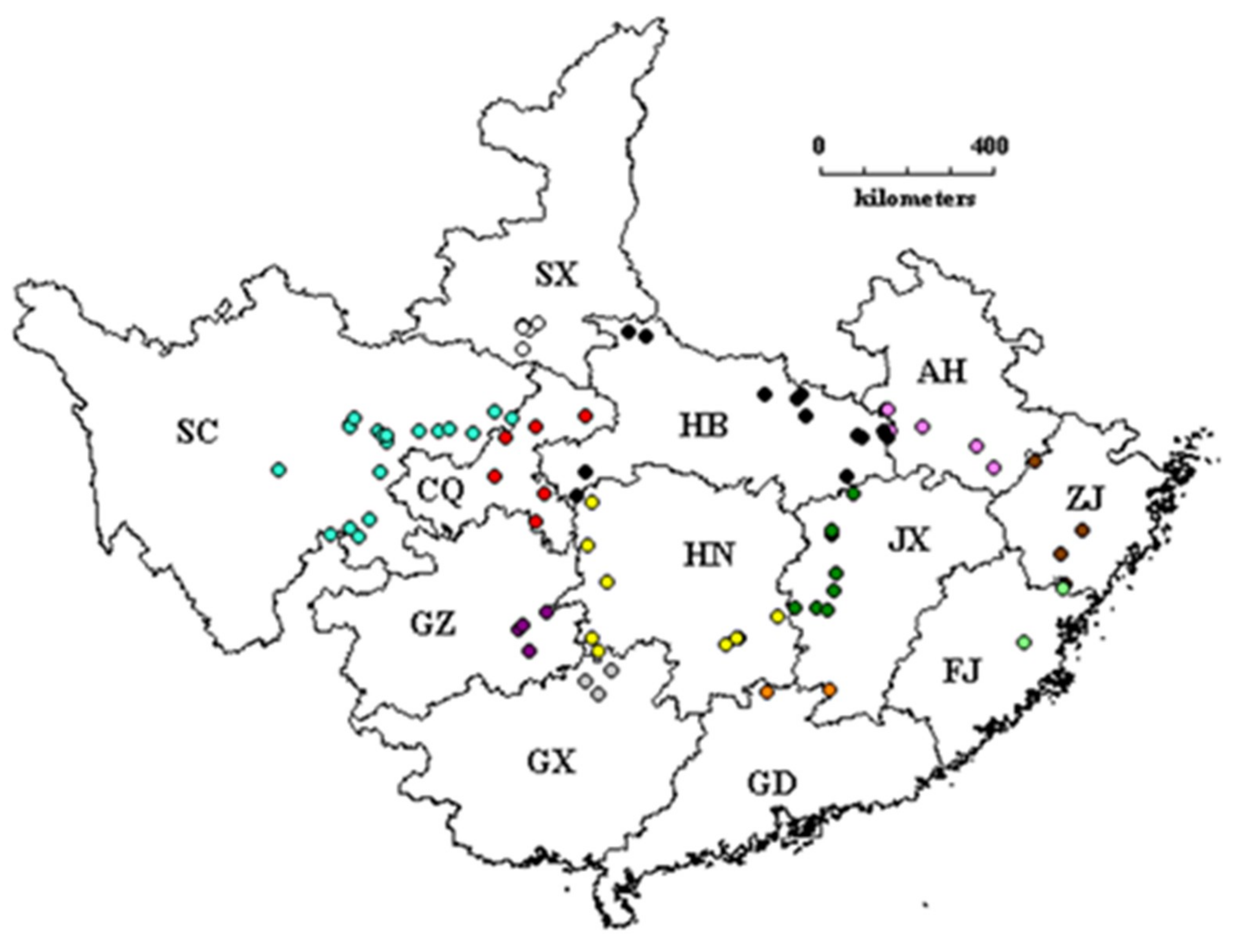
2.1. Collection of Tung Tree Germplasm
2.2. SSR Genotyping and Data Analysis
3. Results
3.1. Genetic Diversity Revealed by SSR Markers in Tung Tree Germplasm
3.2. Group-Wise Genetic Diversity and Geographical Differentiation
3.3. Genetic Diversity and Geographical Differentiation among the Administration Regions
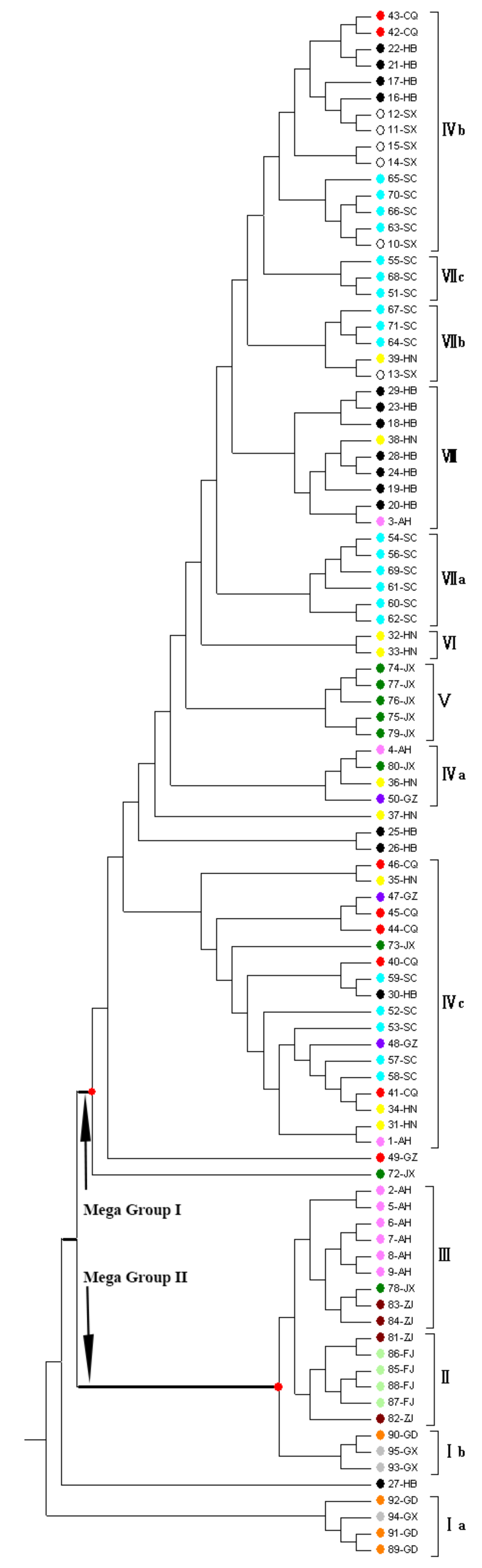
3.4. Genetic Distance and Phylogenetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chen, Y.H.; Chen, J.H.; Luo, Y.M. Complementary biodiesel combination from tung and medium-chain fatty acid oils. Renew. Energy 2012, 44, 305–310. [Google Scholar] [CrossRef]
- Manh, D.V.; Chen, Y.H.; Chang, C.C.; Chang, C.Y.; Hanh, H.D.; Chau, N.H.; Tuyen, T.V.; Long, P.Q.; Minh, C.V. Effects of blending composition of tung oil and ultrasonic irradiation intensity on the biodiesel production. Energy 2012, 48, 519–524. [Google Scholar] [CrossRef]
- Fang, J.X.; He, F. China Tung Oil Tree; Forestry Publishing Press: Beijing, China, 1998. [Google Scholar]
- Chen, J.Z.; Zhang, S.S.; Zhang, X.; Wang, W.J.; Deng, M.Z. Current status of Vernicia fordii development at home and abroad, and its development strategy in Jianyang. Subtrop. Agric. Res. 2009, 5, 69–72. [Google Scholar]
- Langeland, K.A.; Ferrell, J.A.; Sellers, B.; MacDonald, G.E.; Stocker, R.K. Integrated management of nonnative plants in natural areas of Florida. University of Florida-IFAS Publication # SP 242. 2011. [Google Scholar]
- Potter, G.F. The domestic tung industry I Production and improvement of the tung tree. Econ. Bot. 1959, 13, 328–342. [Google Scholar] [CrossRef]
- Cai, J.B.; Ding, Z.J.; Chen, B.Y. Classification of the varieties and forma of China tung tree. Nonwood For. Res. 1997, 15, 47–50. [Google Scholar]
- Long, Z.X. Genetic analysis on the traits related to overwintering of tung tree seedling introduce to the north. J. Northwest For. Coll. 1996, 11, 31–35. [Google Scholar]
- Tan, X.F. Status and suggestion on development of Vernicia fordii. Nonwood For. Res. 2006, 24, 62–64. [Google Scholar]
- Cai, Y.; Sun, D.; Wu, G.; Peng, J.H. ISSR-based genetic diversity of Jatropha curcas germplasm in China. Biomass Bioenerg. 2010, 34, 1739–1750. [Google Scholar] [CrossRef]
- Derero, A.; Gailing, O.; Finkeldey, R. Maintenance of genetic diversity in Cordia africana Lam, a declining forest tree species in Ethiopia. Tree Genet. Genomes 2011, 7, 1–9. [Google Scholar] [CrossRef]
- Urrestarazu, J.; Miranda, C.; Santesteban, L.G.; Royo, J.B. Genetic diversity and structure of local apple cultivars from Northeastern Spain assessed by microsatellite markers. Tree Genet. Genomes 2012, 8, 1163–1180. [Google Scholar] [CrossRef]
- Li, P.; Zhang, X.P.; Chen, Y.C.; Liu, G.Q.; Zhou, G.; Wang, Y.D. Genetic diversity and germplasm resource research on tung tree (Vernicia fordii) cultivars, investigated by inter-simple sequence repeats. Afr. J. Bioechnol. 2008, 7, 1054–1059. [Google Scholar]
- Zhang, L.L.; Luo, M.C.; You, F.M.; Nevo, E.; Lu, S.Y.; Sun, D.F.; Peng, J.H. Development of microsatellite markers in tung tree (Vernicia fordii) using cassava genomic sequences. Plant Mol. Biol. Reporter. 2015, 33, 893–904. [Google Scholar] [CrossRef]
- Zhang, L.L.; Lu, S.Y.; Sun, D.F.; Peng, J.H. Genetic variation and geographic differentiation revealed using ISSR markers in tung tree, Vernicia fordii. J. Genet. 2015, 94, e5–e9. [Google Scholar] [CrossRef] [PubMed]
- Karan, M.; Evans, D.S.; Reilly, D.; Schulte, K.; Wright, C.; Innes, D.; Holton, T.A.; Nikles, D.G.; Dickinson, G.R. Rapid microsatellite marker development for African mahogany Khaya senegalensis, Meliaceae using next-generation sequencing and assessment of its intra-specific genetic diversity. Mol. Ecol. Resour. 2012, 12, 344–353. [Google Scholar] [CrossRef] [PubMed]
- Vieira, M.L.; Santini, L.; Diniz, A.L.; Munhoz, C.F. Microsatellite markers: What they mean and why they are so useful. Genet. Mol. Biol. 2016, 39, 312–328. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.H.; Ye, Q.G.; Kang, M.; Zhou, J.F.; Xu, Y.Q.; Wang, Y.; Huang, H.W. Characterization of microsatellite markers in the endangered Sinojackia xylocarpa, Styracaceae and cross-species amplification in closely related taxa. Mol. Ecol. Notes 2006, 6, 133–136. [Google Scholar] [CrossRef]
- Yang, A.H.; Zhang, J.J.; Tian, H.; Yao, X.H. Characerization of 39 novel EST-SSR markers for Liriodendron tulipofera and cross-species amplification in L chinense, Magnoliaceae. Am. J. Bot. 2012, 99, e460–e464. [Google Scholar] [CrossRef]
- Zhao, H.; Yu, J.Y.; You, F.M.; Luo, M.C.; Peng, J.H. Transferability of microsatellite markers from Brachypodium distachyon to Miscanthus sinensis, a potential biomass crop. J. Integr. Plant Biol. 2011, 53, 232–245. [Google Scholar] [CrossRef]
- Zhang, L.L.; Dai, L.J.; Gou, J.B.; Peng, J.H. An effective protocol to solve the problem in genomic DNA isolation of tung tree. J. Plant Biochem. Biot. 2013, 22, 492–497. [Google Scholar] [CrossRef]
- Pan, Y.; Pan, L.; Chen, L.; Zhang, L.L.; Nevo, E.; Peng, J.H. Development of microsatellite markers in the oil-producing species Vernicia fordii (Euphorbiaceae), a potential biodiesel feedstock. Appl. Plant Sci. 2013, 1, 1–4. [Google Scholar] [CrossRef]
- Yeh, F.C.; Yang, R.C.; Boyle, T.B.J.; Ye, Z.H.; Mao, J.X. POPGENE, The User-Friendly Shareware for Population Genetic Analysis; Molecular Biology and Biotechnology Centre: University of Alberta, Edmonton, AB, Canada, 1997. [Google Scholar]
- Nei, M. Analysis of gene diversity in subdivided populations. Proc. Natl. Acad. Sci. USA 1973, 70, 3321–3323. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GENALEX 6, genetic analysis in Excel Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Wang, Y.; Gu, Y.; Gao, H.; Qiu, L.; Chang, R.; Chen, S.; He, C. Molecular and geographic evolutionary support for the essential role of giganteaa in soybean domestication of flowering time. BMC Evolut. Biol. 2016, 16, 79. [Google Scholar] [CrossRef]
- Liu, Z.X.; Li, H.H.; Wen, Z.X.; Fan, X.H.; Li, Y.; Guan, R.; Gao, Y.; Wang, S.; Wang, D.; Qiu, L. Comparison of genetic diversity between Chinese and American soybean (Glycine max (L.)) accessions revealed by high-density SNPs. Front. Plant Sci. 2017, 8, 2014. [Google Scholar] [CrossRef]
- Chung, W.H.; Jeong, N.; Kim, J.; Lee, W.K.; Lee, Y.G.; Lee, S.H.; Yoon, W.; Kim, J.H.; Choi, I.Y.; Choi, H.K.; et al. Population structure and domestication revealed by high-depth re-sequencing of Korean cultivated and wild Soybean genomes. DNA Res. 2014, 21, 153–167. [Google Scholar] [CrossRef]
- Arora, S.; Steuernagel, B.; Gaurav, K.; Chandramohan, S. Resistance gene cloning from a wild crop relative by sequence capture and association genetics. Nat. Biotechnol. 2019, 37, 139–143. [Google Scholar] [CrossRef]
- Potter, G.F.; Sitton, B.G.; Merrill, S.; Wright, R.E.; Johns, D.M. Cold injury to young tung trees in Northern Louisiana. J. Am. Soc. Hortic. Sci. 1954, 63, 179–181. [Google Scholar]
- Neff, M.S.; Painter, J.H. Factors affecting branching and growth of newly transplanted tung trees trained to vase form. J. Am Soc. Hortic. Sci. 1948, 52, 75–80. [Google Scholar]
- Drosdoff, M.; Kilby, W.W.; Potter, G.F. Response of tung trees to phosphorus and other elements on Savannah very fine sandy loam. Soil Sci. 1954, 78, 361–366. [Google Scholar] [CrossRef]
- Peng, J.H.; Sun, D.F.; Nevo, E. Domestication evolution, genetics and genomics in wheat. Mol. Breed. 2011, 28, 281–301. [Google Scholar] [CrossRef]
- Liu, P.H.; Li, L.S.; Xie, F.M.; Gao, S.Q.; Zhen, D.S. Screening for good cultivars of tung tree. Nonwood For. Res. 1993, 11, 88–89. [Google Scholar]



{kind=link}
{kind=link}
| Primer Code | Forward Primer | Reverse Primer | Primer Code | Forward Primer | Reverse Primer |
|---|---|---|---|---|---|
| Xwtm4 | GGAAACTGCTTGCACAAAGA | CAGCAAGACCATCACCAGTTT | vfSSR9 | GATCGAGTGCTTCATGTGCT | TGACTAGGAAATCTCACTTTAG |
| Xwtm27 | GCCAGCAAGGTTTGCTACAT | AACTGTCAAACCATTCTACTTGC | vfSSR10 | TGAAAGTAGGGGCACAGCTT | TTCACACTCATGGCACTGCT |
| Xwtm50 | TTCTATAGACCAGGGGCCAA | TTTATCCCCAAAGTTGCAGG | vfSSR12 | CATCCCATGTCCTTTTCTGG | CTTCATAGGCATGGCCACAT |
| Xwtm51 | TGGCATCTCAAGACTGGTTG | ATTGCGCCTCTGTCATCTCT | vfSSR15 | TGGGTATACAAGAGGCTAGGTT | CTTGACCTCTTGCTCTGTGCT |
| Xwtm56 | TACCCAATTGACTTGAGGGG | AATGGGTTTTAACTTGGGGC | vfSSR16 | GAAGATCACCCTTCCGACAA | CTTCTATCAAGGTTTTCATGCT |
| Xwtm59 | ATCACATCCCATGGGTCACT | CCGCCAACATGGTGTAAGTT | vfSSR20 | CCATCATCTTTTCTCATTTCAC | CCATATTGGCCAAACATCAA |
| Xwtm63 | GTACCAGCAGGTCCCACAAT | ACTTCCTTTTCGCCTGTTGT | vfSSR22 | TTCCTAGAAAAGGGGCGTCT | GCATCATTTGGAGGTCTGGT |
| Xwtm64 | ACTGGCTGTATGGGGTCACT | TGTCAGAGGTGATTTGGGTTC | vfSSR25 | GCCTACAGTCTACAGTTCCAAAAA | CAAAAATTGAGACAACACATGACA |
| Xwtm76 | CAGACAGGAAGTCCAGGAGC | CGTGTCACCACATAGTCCCA | vfSSR26 | AATGAAAGAGCACTGCATGG | TCCAAACACCAAAGCCCTAC |
| Xwtm89 | CGTCAAATTAATGGCGAGGT | ATTGGCAGTGTCTTTGACCC | vfSSR35 | AATGTATGATTGCATGAGAA | CTGGCCATCCATTGATATT |
| Xwtm99 | CCCAACGGCCATATTTAAGA | TTCTTTGCTCAATGCCACAC | vfSSR36 | GACCCACTAACACAAATTGC | TGGATCTAGCATGTGCTCACT |
| Xwtm102 | GAGCTTCAGCCAGAGGTGAG | GTAGAATGGGCGTCACTGGT | vfSSR40 | CGGAGTTAGTGGCATGT | CCTTCAAAAACAAAACAGAAGC |
| Xwtm104 | CGGGTCACATGCTAATGGAG | TCCACTGCCCTTATGTCTCA | vfSSR41 | AAGACCGGCGAAAGCTAAC | CAAGCCCAACATTTCTACC |
| Xwtm113 | TTCTGCATTCAGATTCTCCAAA | TTCTTCTTCTTCTTCTTCTTCCTCC | vfSSR44 | GGGGAGCTCAAAGAAAAGA | CTTTATATGCACAATCATTGAC |
| Xwtm116 | CTGCCATTTGCAAGGAAGAT | ATCCAAGAAGCACATCAGGC | vfSSR45 | GTTGGAAACGGAGGTAGAA | AAGCAGAAAAGGAGAGACAAAA |
| Xwtm124 | TTTGATTTGGTTTAGTTGATTTTGA | CAAGCCTTTCTCTTGGTTCG | vfSSR49 | ATTACATGAATGTTCGGGATCT | AAGCTGTAGGCGTCGGATA |
| Xwtm125 | AAATAAGCGCCGAGTTTTGA | TAAGGAAAAGTGGATGGGCA | vfSSR50 | TGAACCAGAGAAACAAACG | AACCAGAACTCTTCTTCTTTTT |
| Xwtm152 | GTGAGGATTCGCACTTAGCC | TCCAAAGTCACCTCCAAACA | vfSSR56 | CAAACTGTAATACCCTAAGGA | CAGTGGCAGCATCTCTTTT |
| Xwtm160 | TGCGATTAAGTGTGAAGGCA | TACGCAAGCCCTTAATCTGG | vfSSR57 | GTAATTTTACATGCTGGTG | AGAATGCATGTGCTGTTGC |
| Xwtm168 | CCTACGCTCCTCCATGAAAA | TAAACAACACCGTTCCGTCA | vfSSR58 | AAAATAACCGTATAAGACA | TCCCAAGTTTCTTTGGACATT |
| Xwtm181 | AACCGCAACAATGACCTTTC | CGGTAAACTCGAAGCTGAGG | vfSSR61 | GGTGAATACTTCGTTGGTCTT | CTCAACACTATGCACATAACCA |
| Xwtm183 | TATTGGTGGCCATGTCTTCA | ACCGGATGCACCATAAACAT | vfSSR63 | TGTTTGTTTCTATCTTCCCTCTTTT | GCGTAACGTTTCACTCTCC |
| Xwtm207 | CAGCTAGAGTTGGTCCGAGG | GGTTCGATTCGGTTTCTGAC | vfSSR65 | TTGGGAGATAGCCAAAGCA | AGAGAGGTGGGTACTGAAGTG |
| Xwtm222 | TTTAAACGTGTTATGGGGGC | CGGTTCTACTTCACACCCAAA | vfSSR73 | ACAACAAAACTAGAGAAAC | CTTCGGAGCGTCACTTCTT |
| Xwtm227 | TTAGGTGAATAGGGCGATGG | TAACTTGGGATGGACCTTGC | vfSSR76 | TGCGGAACAGAGAACTAAGAGA | CCCCTAATATGGTTGCCTACTTT |
| SSR Type | Group | GS | NPL | PR | Na | Ne | He | I |
|---|---|---|---|---|---|---|---|---|
| All SSR | Anhui | 9 | 65 | 34.57 | 1.3457 | 1.2287 | 0.1314 | 0.1941 |
| Shaanxi | 6 | 46 | 24.47 | 1.2447 | 1.1525 | 0.0910 | 0.1359 | |
| Hubei | 15 | 88 | 46.81 | 1.4681 | 1.2367 | 0.1453 | 0.2237 | |
| Hunan | 9 | 77 | 40.96 | 1.4096 | 1.2572 | 0.1516 | 0.2259 | |
| Chongqing | 6 | 65 | 34.57 | 1.3457 | 1.2292 | 0.1334 | 0.1972 | |
| Guizhou | 5 | 72 | 38.30 | 1.3830 | 1.2700 | 0.1543 | 0.2259 | |
| Sichuan | 21 | 83 | 44.15 | 1.4415 | 1.2289 | 0.1383 | 0.2117 | |
| Jiangxi | 9 | 76 | 40.43 | 1.4043 | 1.2433 | 0.1435 | 0.2154 | |
| Zhejiang | 4 | 47 | 25.00 | 1.2500 | 1.1830 | 0.1042 | 0.1516 | |
| Fujian | 4 | 41 | 21.81 | 1.2181 | 1.1596 | 0.0909 | 0.1322 | |
| Guangdong | 4 | 57 | 30.32 | 1.3032 | 1.2170 | 0.1247 | 0.1820 | |
| Guangxi | 3 | 47 | 25.00 | 0.0125 | 1.2021 | 0.1117 | 0.1597 | |
| Cassava SSR | Marginal region | 39 | 60 | 56.07 | 1.5607 | 1.2595 | 0.157 | 0.243 |
| Tung tree SSR | 39 | 40 | 56.34 | 1.5634 | 1.2481 | 0.1554 | 0.2435 | |
| All SSR | 39 | 108 | 57.45 | 1.5745 | 1.2586 | 0.1590 | 0.2476 | |
| Cassava SSR | Main production region | 56 | 72 | 67.29 | 1.6729 | 1.3414 | 0.2024 | 0.3092 |
| Tung tree SSR | 56 | 48 | 67.61 | 1.6761 | 1.297 | 0.1807 | 0.2806 | |
| All SSR | 56 | 128 | 68.09 | 1.6809 | 1.3279 | 0.1959 | 0.3007 | |
| Cassava SSR | Whole region | 95 | 74 | 69.16 | 1.6916 | 1.3177 | 0.1904 | 0.2943 |
| Tung tree SSR | 95 | 49 | 69.01 | 1.6901 | 1.2961 | 0.1806 | 0.2809 | |
| All SSR | 95 | 131 | 69.68 | 1.6968 | 1.3134 | 0.1891 | 0.2927 |
| Source of Variation | d.f. | Sum of Squares | Variance Component | Percentage of Variation |
|---|---|---|---|---|
| Between-region | 1 | 73.209 | 1.206 | 6.35% |
| Within-region | 93 | 1653.191 | 17.776 | 93.64% |
| Total | 94 | 1726.400 | 18.982 | 100% |
| Fixation index, Fst = 0.064, p = 0.010; Genetic distance, GD = 0.0185 | ||||
| Source of Variation | d.f. | Sum of Squares | Variance Components | Percentage of Variation |
|---|---|---|---|---|
| Among groups | 11 | 497.534 | 3.993 | 21.24% |
| Within groups | 83 | 1228.866 | 14.806 | 78.76% |
| Total | 94 | 1726.400 | 18.799 | 100% |
| Fixation index, Fst = 0.212, p = 0.001 | ||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Liu, X.; Peng, J. Genetic Diversity and Geographic Differentiation of Tung Tree, Vernicia Fordii (Euphorbiaceae), A Potential Biodiesel Plant Species with Low Invasion Risk. Agronomy 2019, 9, 402. https://doi.org/10.3390/agronomy9070402
Zhang L, Liu X, Peng J. Genetic Diversity and Geographic Differentiation of Tung Tree, Vernicia Fordii (Euphorbiaceae), A Potential Biodiesel Plant Species with Low Invasion Risk. Agronomy. 2019; 9(7):402. https://doi.org/10.3390/agronomy9070402
Chicago/Turabian StyleZhang, Lingling, Xionglun Liu, and Junhua Peng. 2019. "Genetic Diversity and Geographic Differentiation of Tung Tree, Vernicia Fordii (Euphorbiaceae), A Potential Biodiesel Plant Species with Low Invasion Risk" Agronomy 9, no. 7: 402. https://doi.org/10.3390/agronomy9070402
APA StyleZhang, L., Liu, X., & Peng, J. (2019). Genetic Diversity and Geographic Differentiation of Tung Tree, Vernicia Fordii (Euphorbiaceae), A Potential Biodiesel Plant Species with Low Invasion Risk. Agronomy, 9(7), 402. https://doi.org/10.3390/agronomy9070402