
Responses of Polyamine-Metabolic Genes to Polyamines and Plant Stress Hormones in Arabidopsis Seedlings




Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Plant Growth Conditions
2.3. RNA Extraction and Expression Analyses
2.4. T-DNA Construction and Plant Transformation
2.5. GUS Assays
2.6. Statistics
3. Results
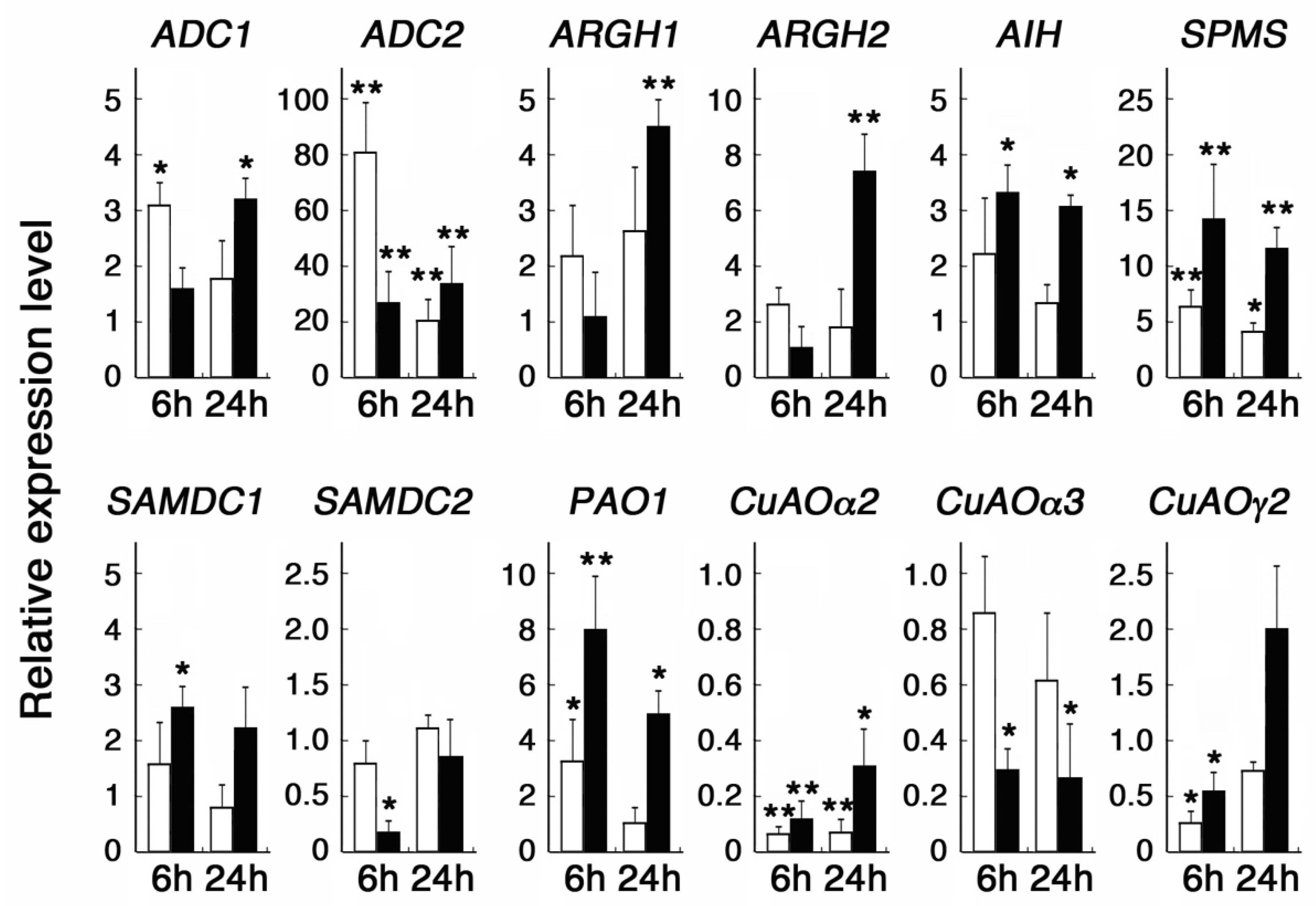
3.1. Responses of Polyamine Metabolic Genes to Polyamines
3.2. Responses of Polyamine Metabolic Genes to Plant Stress Hormones
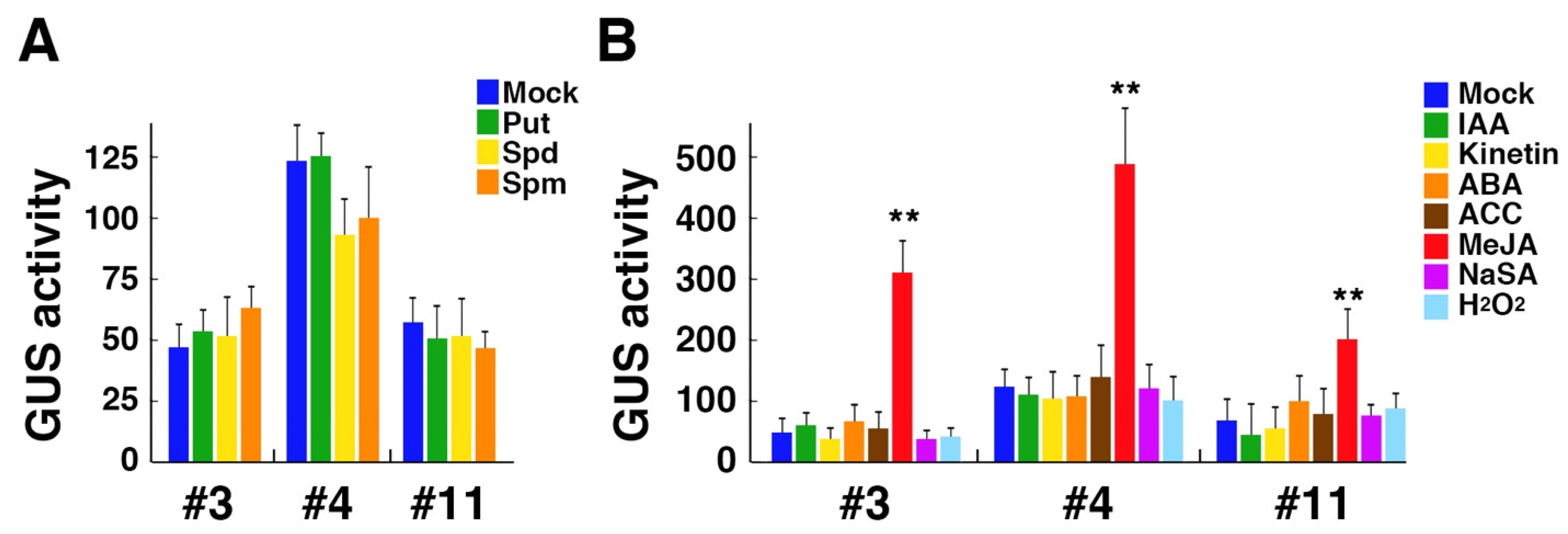
3.3. OsPAO6 Promoter Is Responsive to MeJA in Arabidopsis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Cuevas, J.C.; López-Cobollo, R.; Alcázar, R.; Zarza, X.; Koncz, C.; Altabella, T.; Salinas, J.; Tiburcio, A.F.; Ferrando, A. Putrescine is involved in Arabidopsis freezing tolerance and cold acclimation by regulating ABA levels in response to low temperature. Plant Physiol. 2008, 148, 1094–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, F.R.; Marina, M.; Pieckenstain, F.L. 2014 Role of Arginine decarboxylase (ADC) in Arabidopsis thaliana defense against the pathogenic bacterium Pseudomonas viridiflava. Plant Biol. 2015, 17, 831–839. [Google Scholar] [CrossRef]
- Soyka, S.; Heyer, A.G. Arabidopsis knockout mutation of ADC2 gene reveals inducibility by osmotic stress. FEBS Lett. 1999, 458, 219–223. [Google Scholar] [CrossRef] [Green Version]
- Perez-Amador, M.A.; Leon, J.; Green, P.J.; Carbonell, J. 2002 Induction of the arginine decarboxylase ADC2 gene provides evidence for the involvement of polyamines in the wound response in Arabidopsis. Plant Physiol. 2002, 130, 1454–1463. [Google Scholar] [CrossRef] [Green Version]
- Armengaud, P.; Breitling, R.; Amtmann, A. The potassium-dependent transcriptome of Arabidopsis reveals a prominent role of jasmonic acid in nutrient signaling. Plant Physiol. 2004, 136, 2556–2576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hummel, I.; Bourdais, G.; Gouesbet, G.; Couée, I.; Malmberg, R.L.; El Amrani, A. Differential gene expression of ARGININE DECARBOXYLASE ADC1 and ADC2 in Arabidopsis thaliana: Characterization of transcriptional regulation during seed germination and seedling development. New Phytol. 2004, 163, 519–531. [Google Scholar] [CrossRef]
- Urano, K.; Yoshiba, Y.; Nanjo, T.; Ito, T.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Arabidopsis stress-inducible gene for arginine decarboxylase AtADC2 is required for accumulation of putrescine in salt tolerance. Biochem. Biophys. Res. Commun. 2004, 313, 369–375. [Google Scholar] [CrossRef]
- Janowitz, T.; Kneifel, H.; Piotrowski, M. Identification and characterization of plant agmatine iminohydrolase, the last missing link in polyamine biosynthesis of plants. FEBS Lett. 2003, 544, 258–261. [Google Scholar] [CrossRef] [Green Version]
- Piotrowski, M.; Janowitz, T.; Kneifel, H. 2002 Plant C-N-hydrolases: Identification of a plant N-carbamoylputrescine amidohydrolase involved in polyamine biosynthesis. J. Biol. Chem. 2003, 278, 1708–1712. [Google Scholar] [CrossRef] [Green Version]
- Patel, J.; Ariyaratne, M.; Ahmed, S.; Ge, L.; Phuntumart, V.; Kalinoski, A.; Morris, P.F. Dual functioning of plant arginases provides a third route for putrescine synthesis. Plant Sci. 2017, 262, 62–73. [Google Scholar] [CrossRef]
- Hanzawa, Y.; Imai, A.; Michael, A.J.; Komeda, Y.; Takahashi, T. Characterization of the spermidine synthase-related gene family in Arabidopsis thaliana. FEBS Lett. 2002, 527, 176–180. [Google Scholar] [CrossRef] [Green Version]
- Kakehi, J.-I.; Kuwashiro, Y.; Niitsu, M.; Takahashi, T. Thermospermine is required for stem elongation in Arabidopsis thaliana. Plant Cell Physiol. 2008, 49, 1342–1349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanfrey, C.; Elliott, K.A.; Franceschetti, M.; Mayer, M.J.; Illingworth, C.; Michael, A.J. A dual upstream open reading frame-based autoregulatory circuit controlling polyamine-responsive translation. J. Biol. Chem. 2005, 280, 39229–39237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaszler, N.; Benkő, P.; Bernula, D.; Szepesi, Á.; Fehér, A.; Gémes, K. Polyamine metabolism is involved in the direct regeneration of shoots from Arabidopsis lateral root primordia. Plants 2021, 10, 305. [Google Scholar] [CrossRef] [PubMed]
- Cona, A.; Rea, G.; Angelini, R.; Federico, R.; Tavladoraki, P. Functions of amine oxidases in plant development and defense. Trends Plant Sci. 2006, 11, 80–88. [Google Scholar] [CrossRef]
- Angelini, R.; Tisi, A.; Rea, G.; Chen, M.M.; Botta, M.; Federico, R.; Cona, A. Involvement of polyamine oxidase in wound healing. Plant Physiol. 2008, 146, 162–177. [Google Scholar] [CrossRef] [Green Version]
- Tavladoraki, P.; Cona, A.; Angelini, R. Copper-containing amine oxidases and FAD-dependent polyamine oxidases are key players in plant tissue differentiation and organ development. Front. Plant Sci. 2016, 7, 824–925. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Paschalidis, K.; Feng, J.-C.; Song, J.; Liu, J.-H. Polyamine catabolism in plants: A universal process with diverse functions. Front. Plant Sci. 2019, 10, 561. [Google Scholar] [CrossRef] [Green Version]
- Planas-Portell, J.; Gallart, M.; Tiburcio, A.F.; Altabella, T. Copper-containing amine oxidases contribute to terminal polyamine oxidation in peroxisomes and apoplast of Arabidopsis thaliana. BMC Plant Biol. 2013, 13, 109. [Google Scholar] [CrossRef] [Green Version]
- Fraudentali, I.; Ghuge, S.A.; Carucci, A.; Tavladoraki, P.; Angelini, R.; Rodrigues-Pousada, R.A.; Cona, A. Developmental, hormone- and stress-modulated expression profiles of four members of the Arabidopsis copper-amine oxidase gene family. Plant Physiol. Biochem. 2020, 147, 141–160. [Google Scholar] [CrossRef]
- Fraudentali, I.; Ghuge, S.A.; Carucci, A.; Tavladoraki, P.; Angelini, R.; Cona, A.; Rodrigues-Pousada, R.A. The copper amine oxidase AtCuAOδ participates in abcisic acid-induced stomatal closure in Arabidopsis. Plants 2019, 8, 183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moschou, P.N.; Sanmartin, M.; Andriopoulou, A.H.; Rojo, E.; Sanchez-Serrano, J.J.; Roubelakis-Angelakis, K.A. Bridging the gap between plant and mammalian polyamine catabolism: A novel peroxisomal polyamine oxidase responsible for a full back-conversion pathway in Arabidopsis. Plant Physiol. 2008, 147, 1845–1857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, F.R.; Gárriz, A.; Marina, M.; Pieckenstain, F.L. Modulation of polyamine metabolism in Arabidopsis thaliana by salicylic acid. Physiol. Plant. 2021, in press. [Google Scholar] [CrossRef] [PubMed]
- Marina, M.; Sirera, F.V.; Rambla, J.L.; Gonzalez, M.E.; Blázquez, M.A.; Carbonell, J.; Pieckenstain, F.L.; Ruiz, O.A. Thermospermine catabolism increases Arabidopsis thaliana resistance to Pseudomonas viridiflava. J. Exp. Bot. 2013, 64, 1393–1402. [Google Scholar] [CrossRef] [Green Version]
- Bordenave, C.D.; Granados-Mendoza, C.; Jiménez-Bremont, J.F.; Gárriz, A.; Rodríguez, A.A. Defining novel plant polyamine oxidase subfamilies through molecular modeling and sequence analysis. BMC Evol. Biol. 2019, 19, 28. [Google Scholar] [CrossRef]
- Miyamoto, M.; Shimao, S.; Tong, W.; Motose, H.; Takahashi, T. Effect of thermospermine on the growth and expression of polyamine-related genes in rice seedlings. Plants 2019, 8, 269. [Google Scholar] [CrossRef] [Green Version]
- Sagor, G.H.M.; Inoue, M.; Kusano, T.; Berberich, T. Expression profile of seven polyamine oxidase genes in rice (Oryza sativa) in response to abiotic stresses, phytohormones and polyamines. Physiol. Mol. Biol. Plants 2021, 27, 1353–1359. [Google Scholar] [CrossRef]
- Upadhyay, R.K.; Fatima, T.; Handa, A.K.; Mattoo, A.K. Polyamines and their biosynthesis/catabolism genes are differentially modulated in response to heat versus cold stress in tomato leaves (Solanum lycopersicum L.). Cells 2020, 9, 1749. [Google Scholar] [CrossRef]
- Tong, W.; Imai, A.; Tabata, R.; Shigenobu, S.; Yamaguchi, K.; Yamada, M.; Hasebe, M.; Sawa, S.; Motose, H.; Takahashi, T. Polyamine resistance is increased by mutations in a nitrate transporter gene NRT1.3 (AtNPF6.4) in Arabidopsis thaliana. Front. Plant Sci. 2016, 7, 834. [Google Scholar] [CrossRef]
- Hanzawa, Y.; Takahashi, T.; Komeda, Y. ACL5: An Arabidopsis gene required for internodal elongation after flowering. Plant J. 1997, 12, 863–874. [Google Scholar] [CrossRef] [Green Version]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [Green Version]
- Jefferson, R.A.; Kavanagh, T.A.; Bevan, M.W. GUS fusions: Beta-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 1987, 6, 3901–3907. [Google Scholar] [CrossRef]
- Tong, W.; Yoshimoto, K.; Kakehi, J.-I.; Motose, H.; Niitsu, M.; Takahashi, T. Thermospermine modulates expression of auxin-related genes in Arabidopsis. Front. Plant Sci. 2014, 5, 94. [Google Scholar] [CrossRef] [Green Version]
- Brownfield, D.L.; Todd, C.D.; Deyholos, M.K. Analysis of Arabidopsis arginase gene transcription patterns indicates specific biological functions for recently diverged paralogs. Plant Mol. Biol. 2008, 67, 429–440. [Google Scholar] [CrossRef]
- Park, S.W.; Kaimoyo, E.; Kumar, D.; Mosher, S.; Klessig, D.F. Methyl salicylate is a critical mobile signal for plant systemic acquired resistance. Science 2007, 318, 113–116. [Google Scholar] [CrossRef] [PubMed]
- Katayama, H.; Iwamoto, K.; Kariya, Y.; Asakawa, T.; Kan, T.; Fukuda, H.; Ohashi-Ito, K. A negative feedback loop controlling bHLH complexes is involved in vascular cell division and differentiation in the root apical meristem. Curr. Biol. 2015, 25, 3144–3150. [Google Scholar] [CrossRef] [Green Version]
- Vera-Sirera, F.; De Rybel, B.; Úrbez, C.; Kouklas, E.; Pesquera, M.; Álvarez-Mahecha, J.C.; Minguet, E.G.; Tuominen, H.; Carbonell, J.; Borst, J.W.; et al. A bHLH-based feedback loop restricts vascular cell proliferation in plants. Dev. Cell 2015, 35, 432–443. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Atanasov, K.E.; Arafaty, N.; Murillo, E.; Tiburcio, A.F.; Zeier, J.; Alcázar, R. Putrescine elicits ROS-dependent activation of the salicylic acid pathway in Arabidopsis thaliana. Plant Cell Environ. 2020, 43, 2755–2768. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.S.; Lee, S.H.; Hwang, S.B.; Park, K.Y. Characterization and translational regulation of the arginine decarboxylase gene in carnation (Dianthus caryophyllus L.). Plant J. 2000, 24, 45–56. [Google Scholar] [CrossRef]
- Hiragori, Y.; Takahashi, H.; Hayashi, N.; Sasaki, S.; Nakao, K.; Motomura, T.; Yamashita, Y.; Naito, S.; Onouchi, H. Genome-wide identification of Arabidopsis non-AUG-initiated upstream ORFs with evolutionarily conserved regulatory sequences that control protein expression levels. bioRxiv 2021. [Google Scholar] [CrossRef]
- Hao, Y.; Huang, B.; Jia, D.; Mann, T.; Jiang, X.; Qiu, Y.; Niitsu, M.; Berberich, T.; Kusano, T.; Liu, T. Identification of seven polyamine oxidase genes in tomato (Solanum lycopersicum L.) and their expression profiles under physiological and various stress conditions. J. Plant Physiol. 2018, 228, 1–11. [Google Scholar] [CrossRef]
- Guerrero-González, M.L.; Rodríguez-Kessler, M.; Jiménez-Bremont, J.F. uORF, a regulatory mechanism of the Arabidopsis polyamine oxidase 2. Mol. Biol. Rep. 2014, 41, 2427–2443. [Google Scholar] [CrossRef]
- Guerrero-González, M.L.; Ortega-Amaro, M.A.; Juárez-Montiel, M.; Jiménez-Bremont, J.F. Arabidopsis Polyamine oxidase-2 uORF is required for downstream translational regulation. Plant Physiol. Biochem. 2016, 108, 381–390. [Google Scholar] [CrossRef]
- Sagor, G.H.M.; Kusano, T.; Berberich, T. Identification of the actual coding region for polyamine oxidase 6 from rice (OsPAO6) and its partial characterization. Plant Signal. Behav. 2017, 12, e1359456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wimalasekera, R.; Villar, C.; Begum, T.; Scherer, G.F. COPPER AMINE OXIDASE1 (CuAO1) of Arabidopsis thaliana contributes to abscisic acid- and polyamine-induced nitric oxide biosynthesis and abscisic acid signal transduction. Mol. Plant 2011, 4, 663–678. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Pottosin, I.; Lamade, E.; Tcherkez, G. What is the role of putrescine accumulated under potassium deficiency? Plant Cell Environ. 2020, 43, 1331–1347. [Google Scholar] [CrossRef] [PubMed]
- Schraut, D.; Heilmeier, H.; Hartung, W. Radial transport of water and abscisic acid (ABA) in roots of Zea mays under conditions of nutrient deficiency. J. Exp. Bot. 2005, 56, 879–886. [Google Scholar] [CrossRef] [Green Version]
- Réthoré, E.; Jing, L.; Ali, N.; Yvin, J.C.; Pluchon, S.; Hosseini, S.A. K deprivation modulates the primary metabolites and increases putrescine concentration in Brassica napus. Front. Plant. Sci. 2021, 12, 681895. [Google Scholar] [CrossRef]
- Németh, M.; Janda, T.; Horvat, E.; Paldi, E.; Szalai, G. Exogenous salicylic acid increases polyamine content but may decrease drought tolerance in maize. Plant. Sci. 2002, 162, 569–574. [Google Scholar] [CrossRef]
- Jang, E.-K.; Min, K.-H.; Kim, S.-H.; Nam, S.-H.; Zhang, S.; Kim, Y.C.; Cho, B.H.; Yang, K.-Y. Mitogen-activated protein kinase cascade in the signaling for polyamine biosynthesis in tobacco. Plant. Cell Physiol. 2009, 50, 658–664. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Shen, L.; Li, F.; Meng, D.; Sheng, J. Methyl salicylate-induced arginine catabolism is associated with up-regulation of polyamine and nitric oxide levels and improves chilling tolerance in cherry tomato fruit. J. Agric. Food Chem. 2011, 59, 9351–9357. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.M.; Chu, C. Ethylene-induced polyamine accumulation in rice (Oryza sativa L.) coleoptiles. Plant. Physiol. 1992, 100, 238–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alcázar, R.; García-Martínez, J.L.; Cuevas, J.C.; Tiburcio, A.F.; Altabella, T. Overexpression of ADC2 in Arabidopsis induces dwarfism and late-flowering through GA deficiency. Plant. J. 2005, 43, 425–436. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Gene Name | Gene ID | Treatment (6 h) | |||
|---|---|---|---|---|---|
| Put 5 mM | Spd 1 mM | Spm 0.5 mM | Tspm 0.1 mM | ||
| ADC1 | AT2G16500 | 1.25 | 0.50 | 1.17 | 1.30 |
| ADC2 | AT4G34710 | 5.90 * | 0.14 * | 1.07 | 0.60 |
| ARGAH1 | AT4G08900 | 1.46 | 1.04 | 1.36 | 0.74 |
| ARGAH2 | AT4G08870 | 1.51 | 1.10 | 1.04 | 1.18 |
| AIH | AT5G08170 | 1.89 | 1.13 | 1.21 | 1.27 |
| CPA | AT2G27450 | 1.33 | 1.20 | 1.21 | 0.89 |
| SPDS1 | AT1G23820 | 1.77 | 1.04 | 1.01 | 1.61 |
| SPDS2 | AT1G70310 | 1.17 | 1.85 | 1.20 | 0.91 |
| SPMS | AT5G53120 | 1.49 | 1.90 | 1.49 | 0.85 |
| ACL5 | AT5G19530 | 1.10 | 2.33 | 1.34 | 0.28 ** |
| SAMDC1 | AT3G02470 | 1.14 | 0.96 | 1.17 | 0.94 |
| SAMDC2 | AT5G15950 | 1.63 | 1.42 | 1.30 | 0.87 |
| SAMDC3 | AT3G25570 | 1.06 | 1.43 | 1.26 | 0.74 |
| SAMDC4/BUD2 | AT5G18930 | 0.64 | 1.63 | 0.65 | 0.19 ** |
| PAO1 | AT5G13700 | 1.54 | 2.13 | 1.22 | 0.69 |
| PAO2 | AT2G43020 | 2.30 | 3.88 * | 1.38 | 0.90 |
| PAO3 | AT3G59050 | 1.49 | 2.35 | 1.28 | 1.04 |
| PAO4 | AT1G65840 | 0.58 | 2.54 | 1.34 | 1.38 |
| PAO5 | AT4G29720 | 1.34 | 2.25 | 1.05 | 1.03 |
| CuAOα1 | AT1G31670 | N.D. | N.D. | N.D. | N.D. |
| CuAOα2 | AT1G31690 | 0.25 | 1.88 | 1.15 | 0.91 |
| CuAOα3/CuAO2 | AT1G31710 | 1.13 | 1.36 | 1.26 | 0.78 |
| CuAOβ/ATAO1 | AT4G14940 | 0.48 | 1.39 | 1.45 | 0.61 |
| CuAOγ1/CuAO1 | AT1G62810 | 1.40 | 1.10 | 0.91 | 1.09 |
| CuAOγ2 | AT3G43670 | 0.69 | 0.95 | 1.36 | 0.87 |
| CuAOδ | AT4G12270 | 0.90 | 1.34 | 2.40 | 0.35 |
| CuAOζ/CuAO3 | AT2G42490 | 1.13 | 1.49 | 1.79 | 0.79 |
| UBQ10 | AT4G05320 | 0.98 | 0.95 | 0.94 | 0.92 |
| Gene Name | Gene ID | Treatment (6 h) | |||||
|---|---|---|---|---|---|---|---|
| IAA 10 μM | Kinetin 5 μM | ABA 1 μM | ACC 10 μM | MeJA 2 μM | NaSA 1 mM | ||
| ADC1 | AT2G16500 | 1.85 | 1.34 | 0.62 | 2.45 | 0.51 | 1.64 |
| ADC2 | AT4G34710 | 4.06 * | 1.19 | 8.01 ** | 27.28 ** | 0.96 | 26.91 ** |
| ARGAH1 | AT4G08900 | 1.13 | 1.08 | 3.95 * | 2.09 | 5.94 * | 1.13 |
| ARGAH2 | AT4G08870 | 2.16 | 1.11 | 9.78 ** | 2.31 | 31.78 ** | 0.97 |
| AIH | AT5G08170 | 1.66 | 0.59 | 5.24 * | 1.34 | 1.65 | 3.23 * |
| CPA | AT2G27450 | 1.74 | 0.74 | 3.84 * | 1.82 | 1.23 | 1.49 |
| SPDS1 | AT1G23820 | 1.14 | 1.72 | 1.55 | 1.36 | 1.07 | 1.40 |
| SPDS2 | AT1G70310 | 1.85 | 1.43 | 2.81 * | 1.07 | 0.89 | 1.15 |
| SPMS | AT5G53120 | 1.70 | 0.90 | 20.25 ** | 1.21 | 1.11 | 13.83 ** |
| ACL5 | AT5G19530 | 3.46 * | 2.34 | 1.10 | 1.01 | 1.49 | 0.72 |
| SAMDC1 | AT3G02470 | 0.88 | 0.58 | 2.47 * | 1.37 | 0.81 | 2.66 |
| SAMDC2 | AT5G15950 | 1.11 | 0.71 | 9.00 ** | 0.83 | 0.77 | 0.14 |
| SAMDC3 | AT3G25570 | 2.08 | 0.77 | 1.39 | 1.47 | 1.20 | 0.85 |
| SAMDC4/BUD2 | AT5G18930 | 2.95 * | 0.92 | 2.15 | 1.41 | 0.91 | 0.40 |
| PAO1 | AT5G13700 | 3.51 * | 1.09 | 5.28 * | 2.73 | 0.73 | 8.00 ** |
| PAO2 | AT2G43020 | 0.72 | 1.05 | 3.16 * | 1.71 | 1.16 | 1.25 |
| PAO3 | AT3G59050 | 2.38 * | 0.83 | 2.39 * | 2.10 | 2.46 * | 1.11 |
| PAO4 | AT1G65840 | 1.04 | 0.56 | 3.25 * | 1.14 | 0.66 | 0.86 |
| PAO5 | AT4G29720 | 2.87 * | 1.52 | 2.03 | 1.73 | 1.88 | 0.33 |
| CuAOα1 | AT1G31670 | N.D. | N.D. | N.D. | N.D. | N.D. | N.D. |
| CuAOα2 | AT1G31690 | 0.19 * | 1.58 | 1.45 | 0.03 ** | 1.17 | 0.11 ** |
| CuAOα3/CuAO2 | AT1G31710 | 0.32 * | 1.61 | 0.89 | 0.88 | 3.84 * | 0.30 * |
| CuAOβ/ATAO1 | AT4G14940 | 0.71 | 1.52 | 1.46 | 1.60 | 0.08 | 0.71 |
| CuAOγ1/CuAO1 | AT1G62810 | 1.38 | 0.70 | 10.10 ** | 1.57 | 0.79 | 0.74 |
| CuAOγ2 | AT3G43670 | 1.01 | 0.54 | 1.51 | 1.00 | 0.88 | 0.55 * |
| CuAOδ | AT4G12270 | 1.26 | 0.16 | 4.72 * | 0.6 | 0.77 | 1.74 |
| CuAOζ/CuAO3 | AT2G42490 | 0.84 | 0.76 | 1.91 | 1.91 | 1.22 | 2.45 |
| UBQ10 | AT4G05320 | 1.02 | 0.94 | 1.11 | 0.96 | 0.94 | 0.92 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yariuchi, Y.; Okamoto, T.; Noutoshi, Y.; Takahashi, T. Responses of Polyamine-Metabolic Genes to Polyamines and Plant Stress Hormones in Arabidopsis Seedlings. Cells 2021, 10, 3283. https://doi.org/10.3390/cells10123283
Yariuchi Y, Okamoto T, Noutoshi Y, Takahashi T. Responses of Polyamine-Metabolic Genes to Polyamines and Plant Stress Hormones in Arabidopsis Seedlings. Cells. 2021; 10(12):3283. https://doi.org/10.3390/cells10123283
Chicago/Turabian StyleYariuchi, Yusaku, Takashi Okamoto, Yoshiteru Noutoshi, and Taku Takahashi. 2021. "Responses of Polyamine-Metabolic Genes to Polyamines and Plant Stress Hormones in Arabidopsis Seedlings" Cells 10, no. 12: 3283. https://doi.org/10.3390/cells10123283
APA StyleYariuchi, Y., Okamoto, T., Noutoshi, Y., & Takahashi, T. (2021). Responses of Polyamine-Metabolic Genes to Polyamines and Plant Stress Hormones in Arabidopsis Seedlings. Cells, 10(12), 3283. https://doi.org/10.3390/cells10123283