
Tributyrin, a Butyrate Pro-Drug, Primes Satellite Cells for Differentiation by Altering the Epigenetic Landscape

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
3. Results
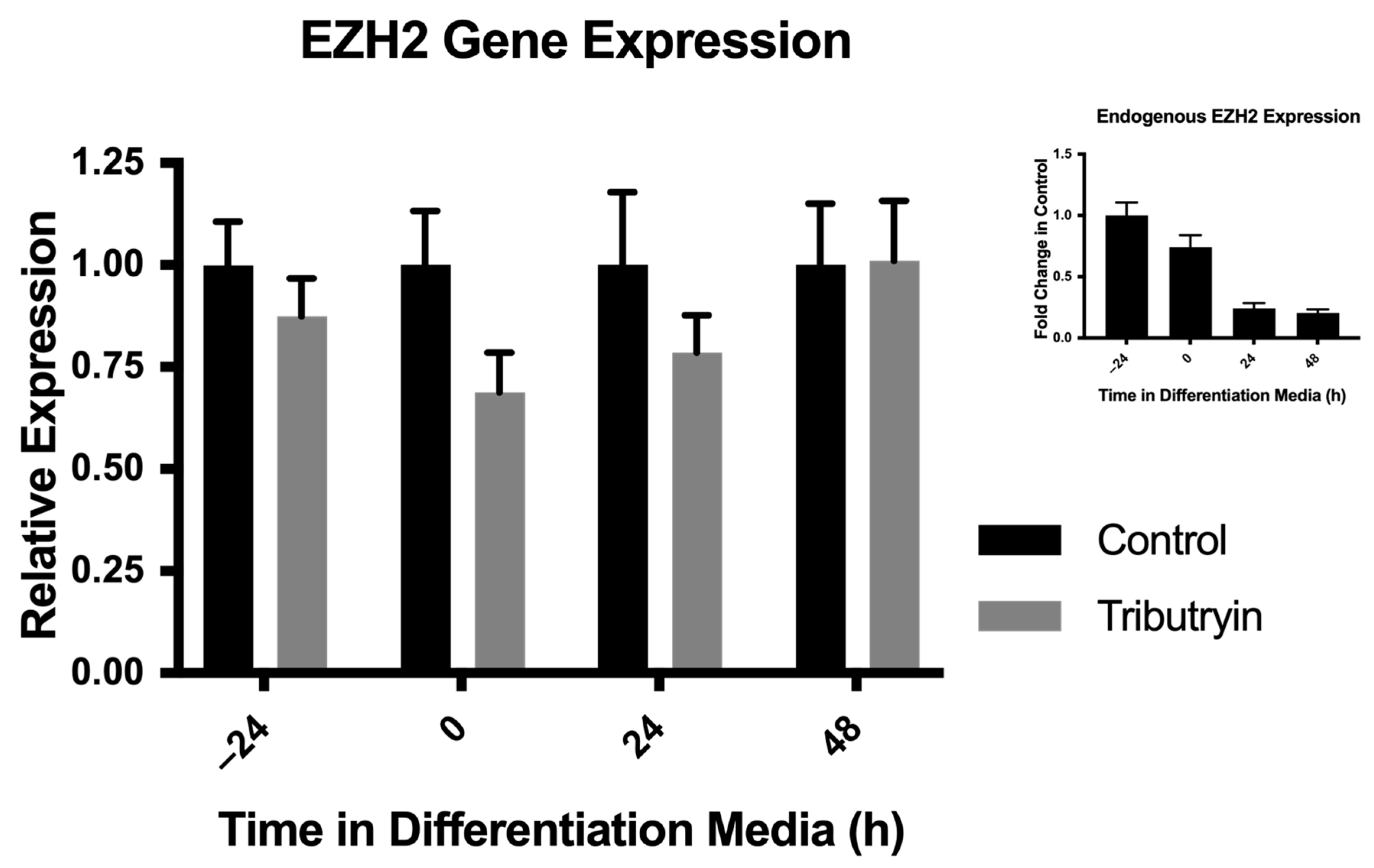
3.1. Tributyrin’s Effect on EZH2 Gene Expression in Satellite Cells
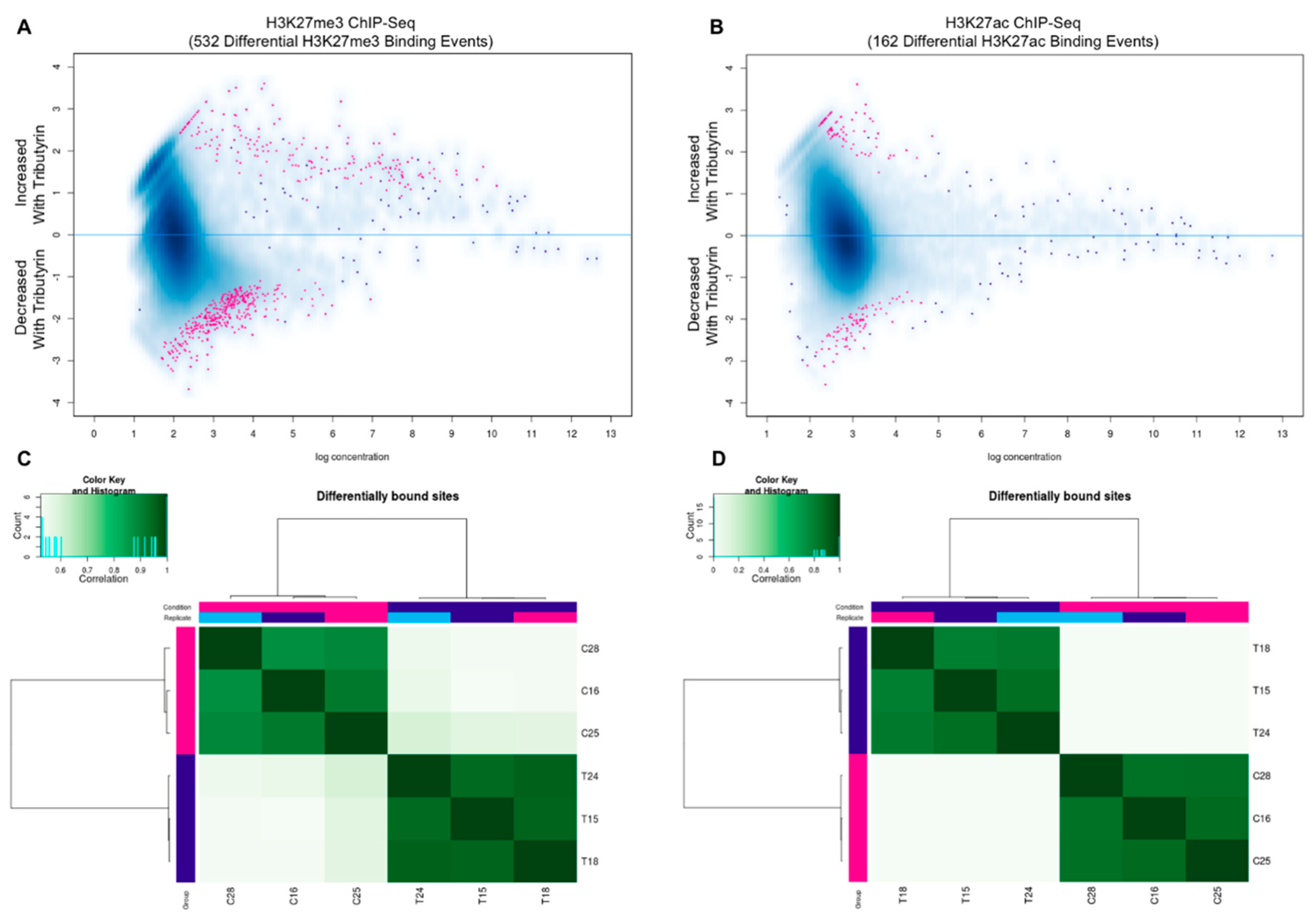
3.2. Genomic Landscape of H3K27me3 and H3K27ac
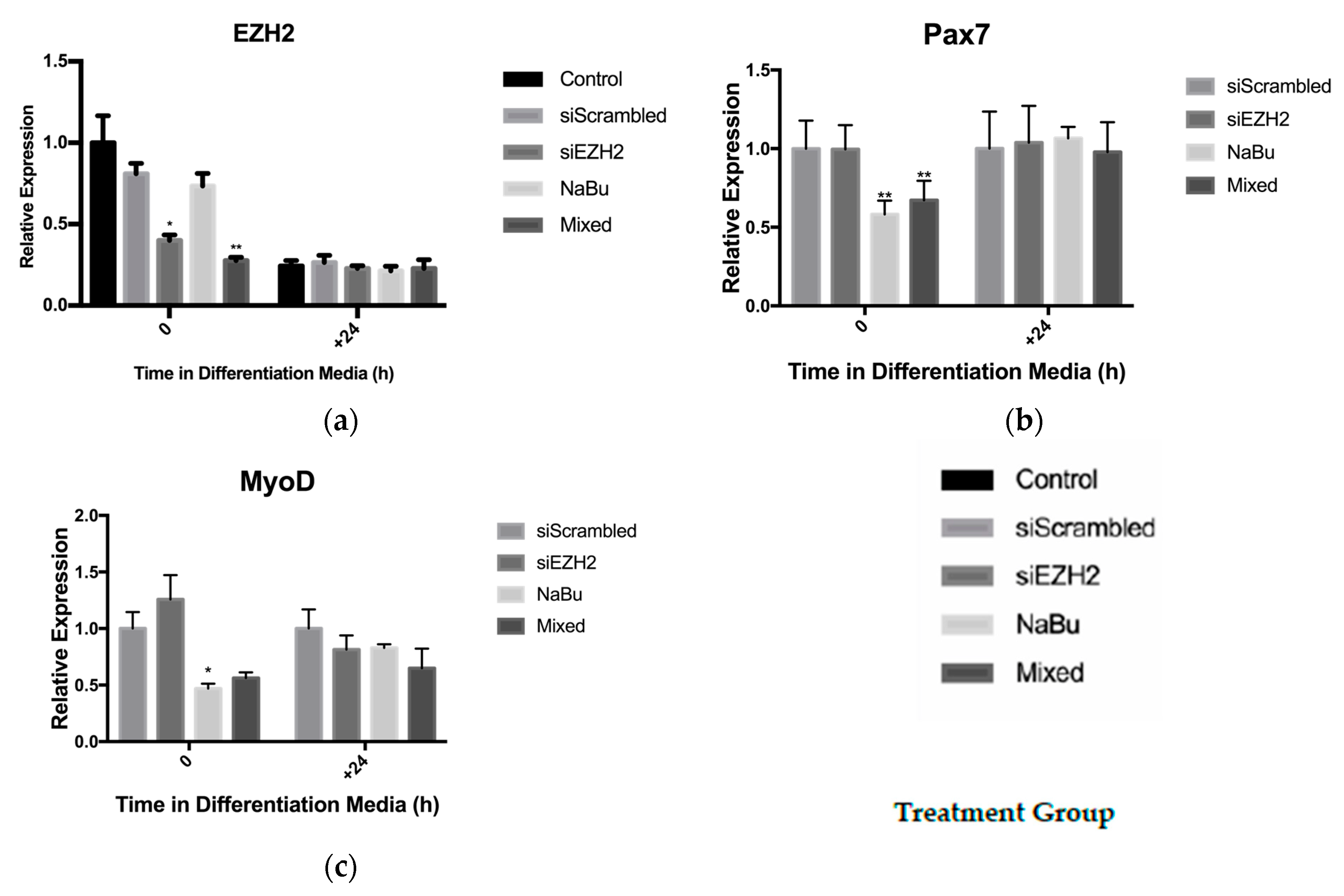
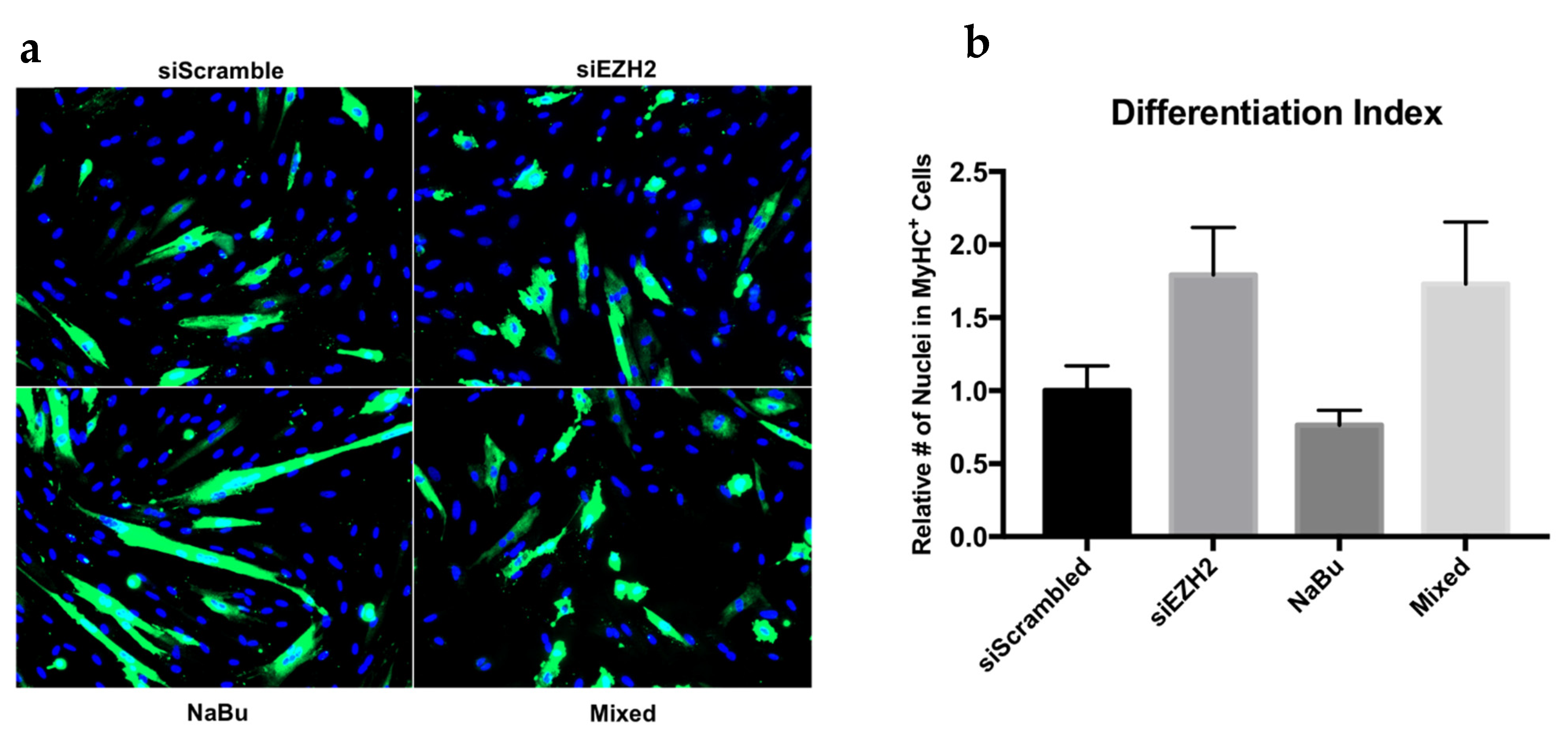
3.3. Butyrate’s Impact on EZH2 Activity and Satellite Cell Myogenesis
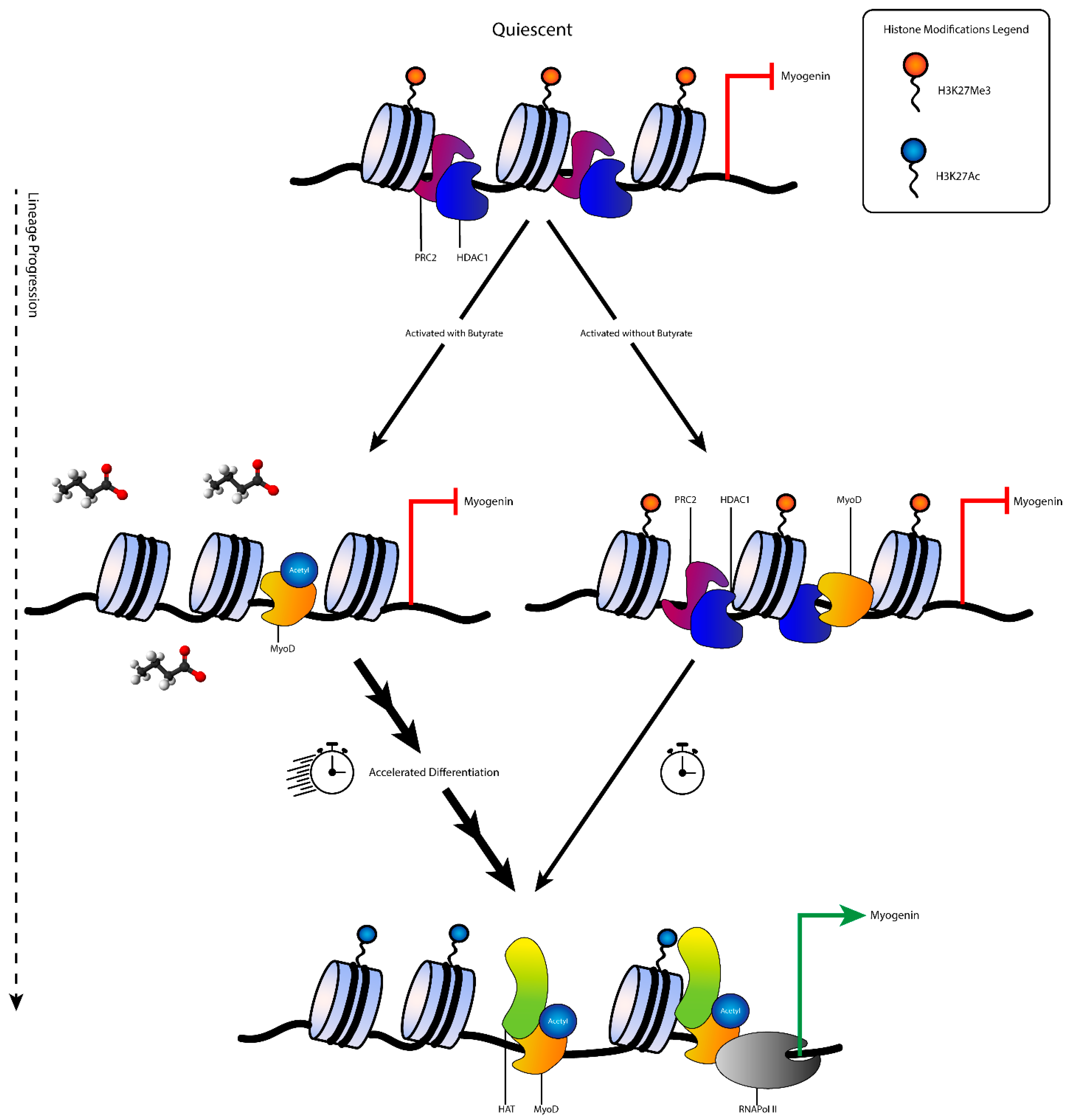
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Bentzinger, C.F.; von Maltzahn, J.; Dumont, N.A.; Stark, D.A.; Wang, Y.X.; Nhan, K.; Frenette, J.; Cornelison, D.; Rudnicki, M.A. Wnt7a stimulates myogenic stem cell motility and engraftment resulting in improved muscle strength. J. Cell Biol. 2014, 205, 97–111. [Google Scholar] [CrossRef]
- Dumont, N.A.; Bentzinger, C.F.; Sincennes, M.C.; Rudnicki, M.A. Satellite cells and skeletal muscle regeneration. Compr. Physiol. 2015, 5, 1027–1059. [Google Scholar] [CrossRef]
- Kuang, S.; Kuroda, K.; Le Grand, F.; Rudnicki, M.A. Asymmetric self-renewal and commitment of satellite stem cells in muscle. Cell 2007, 129, 999–1010. [Google Scholar] [CrossRef] [Green Version]
- Moresi, V.; Marroncelli, N.; Adamo, S. New insights into the epigenetic control of satellite cells. World J. Stem Cells 2015, 7, 945–955. [Google Scholar] [CrossRef] [Green Version]
- Sincennes, M.C.; Brun, C.E.; Rudnicki, M.A. Concise review: Epigenetic regulation of myogenesis in health and disease. Stem Cells Transl. Med. 2016, 5, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Caretti, G.; Di Padova, M.; Micales, B.; Lyons, G.E.; Sartorelli, V. The polycomb Ezh2 methyltransferase regulates muscle gene expression and skeletal muscle differentiation. Genes Dev. 2004, 18, 2627–2638. [Google Scholar] [CrossRef] [Green Version]
- Palacios, D.; Mozzetta, C.; Consalvi, S.; Caretti, G.; Saccone, V.; Proserpio, V.; Marquez, V.E.; Valente, S.; Mai, A.; Forcales, S.V.; et al. TNF/p38alpha/polycomb signaling to Pax7 locus in satellite cells links inflammation to the epigenetic control of muscle regeneration. Cell Stem Cell 2010, 7, 455–469. [Google Scholar] [CrossRef] [Green Version]
- Asp, P.; Blum, R.; Vethantham, V.; Parisi, F.; Micsinai, M.; Cheng, J.; Bowman, C.; Kluger, Y.; Dynlacht, B.D. Genome-wide remodeling of the epigenetic landscape during myogenic differentiation. Proc. Natl. Acad. Sci. USA 2011, 108, E149–E158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dilworth, F.J.; Blais, A. Epigenetic regulation of satellite cell activation during muscle regeneration. Stem Cell. Res. Ther. 2011, 2, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iezzi, S.; Cossu, G.; Nervi, C.; Sartorelli, V.; Puri, P.L. Stage-specific modulation of skeletal myogenesis by inhibitors of nuclear deacetylases. Proc. Natl. Acad. Sci. USA 2002, 99, 7757–7762. [Google Scholar] [CrossRef] [Green Version]
- Iezzi, S.; Di Padova, M.; Serra, C.; Caretti, G.; Simone, C.; Maklan, E.; Minetti, G.; Zhao, P.; Hoffman, E.P.; Puri, P.L.; et al. Deacetylase inhibitors increase muscle cell size by promoting myoblast recruitment and fusion through induction of follistatin. Dev. Cell 2004, 6, 673–684. [Google Scholar] [CrossRef] [Green Version]
- Minetti, G.C.; Colussi, C.; Adami, R.; Serra, C.; Mozzetta, C.; Parente, V.; Fortuni, S.; Straino, S.; Sampaolesi, M.; Di Padova, M.; et al. Functional and morphological recovery of dystrophic muscles in mice treated with deacetylase inhibitors. Nat. Med. 2006, 12, 1147–1150. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.W.; Lee, S.; Seo, M.S.; Park, S.B.; Kurtz, A.; Kang, S.K.; Kang, K.S. Histone deacetylase controls adult stem cell aging by balancing the expression of polycomb genes and jumonji domain containing 3. Cell. Mol. Life Sci. 2010, 67, 1165–1176. [Google Scholar] [CrossRef] [Green Version]
- Murray, R.L.; Zhang, W.; Iwaniuk, M.; Grilli, E.; Stahl, C.H. Dietary tributyrin, an HDAC inhibitor, promotes muscle growth through enhanced terminal differentiation of satellite cells. Physiol. Rep. 2018, 6, e13706. [Google Scholar] [CrossRef] [PubMed]
- Mauro, A. Satellite cell of skeletal muscle fibers. J. Biophys. Biochem. Cytol. 1961, 9, 493–495. [Google Scholar] [CrossRef] [PubMed]
- Blais, A.; Tsikitis, M.; Acosta-Alvear, D.; Sharan, R.; Kluger, Y.; Dynlacht, B.D. An initial blueprint for myogenic differentiation. Genes Dev. 2005, 19, 553–569. [Google Scholar] [CrossRef] [Green Version]
- Crist, C.G.; Montarras, D.; Buckingham, M. Muscle satellite cells are primed for myogenesis but maintain quiescence with sequestration of Myf5 mRNA targeted by microRNA-31 in mRNP granules. Cell Stem Cell 2012, 11, 118–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blais, A. Myogenesis in the genomics era. J. Mol. Biol. 2015, 427, 2023–2038. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.X.; Dumont, N.A.; Rudnicki, M.A. Muscle stem cells at a glance. J. Cell Sci. 2014, 127, 4543–4548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, M.H.; Seale, P.; Rudnicki, M.A. Looking back to the embryo: Defining transcriptional networks in adult myogenesis. Nat. Rev. Genet. 2003, 4, 497–507. [Google Scholar] [CrossRef] [PubMed]
- Seale, P.; Sabourin, L.A.; Girgis-Gabardo, A.; Mansouri, A.; Gruss, P.; Rudnicki, M.A. Pax7 is required for the specification of myogenic satellite cells. Cell 2000, 102, 777–786. [Google Scholar] [CrossRef] [Green Version]
- Olguin, H.C.; Pisconti, A. Marking the tempo for myogenesis: Pax7 and the regulation of muscle stem cell fate decisions. J. Cell. Mol. Med. 2012, 16, 1013–1025. [Google Scholar] [CrossRef]
- Tapscott, S.J. The circuitry of a master switch: Myod and the regulation of skeletal muscle gene transcription. Development 2005, 132, 2685–2695. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, N.; Yoshida, S.; Koishi, K.; Masuda, K.; Nabeshima, Y. Cell heterogeneity upon myogenic differentiation: Down-regulation of MyoD and Myf-5 generates ‘reserve cells’. J. Cell Sci. 1998, 111, 769–779. [Google Scholar] [CrossRef] [PubMed]
- Olguin, H.C.; Yang, Z.; Tapscott, S.J.; Olwin, B.B. Reciprocal inhibition between Pax7 and muscle regulatory factors modulates myogenic cell fate determination. J. Cell Biol. 2007, 177, 769–779. [Google Scholar] [CrossRef]
- Mal, A.; Harter, M.L. MyoD is functionally linked to the silencing of a muscle-specific regulatory gene prior to skeletal myogenesis. Proc. Natl. Acad. Sci. USA 2003, 100, 1735–1739. [Google Scholar] [CrossRef] [Green Version]
- Bergstrom, D.A.; Penn, B.H.; Strand, A.; Perry, R.L.; Rudnicki, M.A.; Tapscott, S.J. Promoter-specific regulation of MyoD binding and signal transduction cooperate to pattern gene expression. Mol. Cell 2002, 9, 587–600. [Google Scholar] [CrossRef]
- Hasty, P.; Bradley, A.; Morris, J.H.; Edmondson, D.G.; Venuti, J.M.; Olson, E.N.; Klein, W.H. Muscle deficiency and neonatal death in mice with a targeted mutation in the myogenin gene. Nature 1993, 364, 501–506. [Google Scholar] [CrossRef]
- De la Serna, I.L.; Ohkawa, Y.; Berkes, C.A.; Bergstrom, D.A.; Dacwag, C.S.; Tapscott, S.J.; Imbalzano, A.N. MyoD targets chromatin remodeling complexes to the myogenin locus prior to forming a stable DNA-bound complex. Mol. Cell. Biol. 2005, 25, 3997–4009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venuti, J.M.; Morris, J.H.; Vivian, J.L.; Olson, E.N.; Klein, W.H. Myogenin is required for late but not early aspects of myogenesis during mouse development. J. Cell Biol. 1995, 128, 563–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davie, J.K.; Cho, J.-H.; Meadows, E.; Flynn, J.M.; Knapp, J.R.; Klein, W.H. Target gene selectivity of the myogenic basic helix–loop–helix transcription factor myogenin in embryonic muscle. Dev. Biol. 2007, 311, 650–664. [Google Scholar] [CrossRef] [Green Version]
- Millay, D.P.; O’Rourke, J.R.; Sutherland, L.B.; Bezprozvannaya, S.; Shelton, J.M.; Bassel-Duby, R.; Olson, E.N. Myomaker is a membrane activator of myoblast fusion and muscle formation. Nature 2013, 499, 301–305. [Google Scholar] [CrossRef] [Green Version]
- Brack, A.S.; Conboy, I.M.; Conboy, M.J.; Shen, J.; Rando, T.A. A temporal switch from notch to Wnt signaling in muscle stem cells is necessary for normal adult myogenesis. Cell Stem Cell 2008, 2, 50–59. [Google Scholar] [CrossRef] [Green Version]
- Olguin, H.C.; Olwin, B.B. Pax-7 up-regulation inhibits myogenesis and cell cycle progression in satellite cells: A potential mechanism for self-renewal. Dev. Biol. 2004, 275, 375–388. [Google Scholar] [CrossRef] [Green Version]
- Spivakov, M.; Fisher, A.G. Epigenetic signatures of stem-cell identity. Nat. Rev. Genet. 2007, 8, 263–271. [Google Scholar] [CrossRef]
- Liu, L.; Cheung, T.H.; Charville, G.W.; Hurgo, B.M.; Leavitt, T.; Shih, J.; Brunet, A.; Rando, T.A. Chromatin modifications as determinants of muscle stem cell quiescence and chronological aging. Cell Rep. 2013, 4, 189–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mal, A.; Sturniolo, M.; Schiltz, R.L.; Ghosh, M.K.; Harter, M.L. A role for histone deacetylase HDAC1 in modulating the transcriptional activity of MyoD: Inhibition of the myogenic program. EMBO J. 2001, 20, 1739–1753. [Google Scholar] [CrossRef] [PubMed]
- Puri, P.L.; Iezzi, S.; Stiegler, P.; Chen, T.T.; Schiltz, R.L.; Muscat, G.E.; Giordano, A.; Kedes, L.; Wang, J.Y.; Sartorelli, V. Class I histone deacetylases sequentially interact with MyoD and pRb during skeletal myogenesis. Mol. Cell 2001, 8, 885–897. [Google Scholar] [CrossRef]
- Blum, R.; Dynlacht, B.D. The role of MyoD1 and histone modifications in the activation of muscle enhancers. Epigenetics 2013, 8, 778–784. [Google Scholar] [CrossRef] [Green Version]
- Stojic, L.; Jasencakova, Z.; Prezioso, C.; Stützer, A.; Bodega, B.; Pasini, D.; Klingberg, R.; Mozzetta, C.; Margueron, R.; Puri, P.L.; et al. Chromatin regulated interchange between polycomb repressive complex 2 (PRC2)-Ezh2 and PRC2-Ezh1 complexes controls myogenin activation in skeletal muscle cells. Epigenet. Chromatin 2011, 4, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodhouse, S.; Pugazhendhi, D.; Brien, P.; Pell, J.M. Ezh2 maintains a key phase of muscle satellite cell expansion but does not regulate terminal differentiation. J. Cell Sci. 2013, 126, 565–579. [Google Scholar] [CrossRef] [Green Version]
- Dong, L.; Zhong, X.; He, J.; Zhang, L.; Bai, K.; Xu, W.; Wang, T.; Huang, X. Supplementation of tributyrin improves the growth and intestinal digestive and barrier functions in intrauterine growth-restricted piglets. Clin. Nutr. 2016, 35, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Song, P.; Fan, P.; Hou, C.; Thacker, P.; Ma, X. Dietary sodium butyrate decreases postweaning diarrhea by modulating intestinal permeability and changing the bacterial communities in weaned piglets. J. Nutr. 2015, 145, 2774–2780. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Su, S.; Ajuwon, K.M. Butyrate supplementation to gestating sows and piglets induces muscle and adipose tissue oxidative genes and improves growth performance. J. Anim. Sci. 2012, 90 (Suppl. S4), 430–432. [Google Scholar] [CrossRef] [Green Version]
- Doumit, M.E.; Merkel, R.A. Conditions for isolation and culture of porcine myogenic satellite cells. Tissue Cell 1992, 24, 253–262. [Google Scholar] [CrossRef]
- Allen, R.E.; Temm-Grove, C.J.; Sheehan, S.M.; Rice, G. Skeletal muscle satellite cell cultures. Methods Cell Biol. 1997, 52, 155–176. [Google Scholar] [PubMed]
- Li, B.J.; Li, P.H.; Huang, R.H.; Sun, W.X.; Wang, H.; Li, Q.F.; Chen, J.; Wu, W.J.; Liu, H.L. Isolation, culture and identification of porcine skeletal muscle satellite cells. Asian-Australas. J. Anim. Sci. 2015, 28, 1171–1177. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, D. ChIP-seq: Using high-throughput sequencing to discover protein-DNA interactions. Methods 2009, 48, 240–248. [Google Scholar] [CrossRef] [Green Version]
- Schmieder, R.; Edwards, R. Quality control and preprocessing of metagenomic datasets. Bioinformatics 2011, 27, 863–864. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liu, T.; Meyer, C.A.; Eeckhoute, J.; Johnson, D.S.; Bernstein, B.E.; Nusbaum, C.; Myers, R.M.; Brown, M.; Li, W.; et al. Model-based analysis of ChIP-Seq (MACS). Genome Biol. 2008, 9, R137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross-Innes, C.S.; Stark, R.; Teschendorff, A.E.; Holmes, K.A.; Ali, H.R.; Dunning, M.J.; Brown, G.D.; Gojis, O.; Ellis, I.O.; Green, A.R.; et al. Differential oestrogen receptor binding is associated with clinical outcome in breast cancer. Nature 2012, 481, 389–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.J.; Gazin, C.; Lawson, N.D.; Pages, H.; Lin, S.M.; Lapointe, D.S.; Green, M.R. ChIPpeakAnno: A Bioconductor package to annotate ChIP-seq and ChIP-chip data. BMC Bioinform. 2010, 11, 237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang da, W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Consalvi, S.; Brancaccio, A.; Dall’Agnese, A.; Puri, P.L.; Palacios, D. Praja1 E3 ubiquitin ligase promotes skeletal myogenesis through degradation of EZH2 upon p38alpha activation. Nat. Commun. 2017, 8, 13956. [Google Scholar] [CrossRef] [Green Version]
- Juan, A.H.; Kumar, R.M.; Marx, J.G.; Young, R.A.; Sartorelli, V. Mir-214-dependent regulation of the polycomb protein Ezh2 in skeletal muscle and embryonic stem cells. Mol. Cell 2009, 36, 61–74. [Google Scholar] [CrossRef] [Green Version]
- Le Gall, M.; Gallois, M.; Seve, B.; Louveau, I.; Holst, J.J.; Oswald, I.P.; Lalles, J.P.; Guilloteau, P. Comparative effect of orally administered sodium butyrate before or after weaning on growth and several indices of gastrointestinal biology of piglets. Br. J. Nutr. 2009, 102, 1285–1296. [Google Scholar] [CrossRef] [Green Version]
- Wang, J. The Msx1 homeoprotein recruits polycomb to the nuclear periphery during development. Dev. Cell 2011, 21, 575–588. [Google Scholar] [CrossRef] [Green Version]
- Prasad, K.N. Butyric acid: A small fatty acid with diverse biological functions. Life Sci. 1980, 27, 1351–1358. [Google Scholar] [CrossRef]
- Chen, J.F.; Tao, Y.; Li, J.; Deng, Z.; Yan, Z.; Xiao, X.; Wang, D.Z. microRNA-1 and microRNA-206 regulate skeletal muscle satellite cell proliferation and differentiation by repressing Pax7. J. Cell Biol. 2010, 190, 867–879. [Google Scholar] [CrossRef] [Green Version]
- Naguibneva, I.; Ameyar-Zazoua, M.; Polesskaya, A.; Ait-Si-Ali, S.; Groisman, R.; Souidi, M.; Cuvellier, S.; Harel-Bellan, A. The microRNA miR-181 targets the homeobox protein Hox-A11 during mammalian myoblast differentiation. Nat. Cell Biol. 2006, 8, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Cicchillitti, L.; Di Stefano, V.; Isaia, E.; Crimaldi, L.; Fasanaro, P.; Ambrosino, V.; Antonini, A.; Capogrossi, M.C.; Gaetano, C.; Piaggio, G.; et al. Hypoxia-inducible factor 1-alpha induces miR-210 in normoxic differentiating myoblasts. J. Biol. Chem. 2012, 287, 44761–44771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, K.; Sha, J.; Harter, M.L. Activation of Cdc6 by MyoD is associated with the expansion of quiescent myogenic satellite cells. J. Cell Biol. 2010, 188, 39–48. [Google Scholar] [CrossRef]
- Von Maltzahn, J.; Jones, A.E.; Parks, R.J.; Rudnicki, M.A. Pax7 is critical for the normal function of satellite cells in adult skeletal muscle. Proc. Natl. Acad. Sci. USA 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GO Categories Enriched in H3K27me3 from Satellite Cells Treated with Tributyrin | ||||
|---|---|---|---|---|
| GO Term | Associated Gene ID | Gene Name/Function | Fold Change | p-Value |
| Skeletal Muscle Cell/Myoblast Differentiation/Cell Differentiation | MYOD1 | myogenic differentiation 1 | −2.059 | 4.2 × 10−4 |
| SOX11 | SRY-box 11 | −2.653 | 3.0 × 10−4 | |
| QKI | KH domain containing RNA binding | −1.398 | 4.7 × 10−4 | |
| NTRK3 | neurotrophic tyrosine kinase, receptor, type 3 | −2.023 | 1.9 × 10−4 | |
| Negative Regulation of Cell Proliferation | DRD2 | dopamine receptor D2 | −2.508 | 3.1 × 10−4 |
| BTG4 | BTG anti-proliferation factor 4 | −2.002 | 1.3 × 10−5 | |
| INSM1 | INSM transcriptional repressor 1 | −1.486 | 3.0 × 10−4 | |
| Negative Regulation of Cell Response to Growth Factor Stimulus | SLIT2 | slit guidance ligand 2 | −2.227 | 4.7 × 10−4 |
| CASK | calcium/calmodulin dependent serine protein kinase | −1.308 | 2.5 × 10−4 | |
| Cell Cycle Arrest | THBS1 | thrombospondin 1 | −1.753 | 4.9 × 10−4 |
| IL-12B | interleukin 12B | −2.251 | 1.8 × 10−7 | |
| Homophilic cell adhesion via plasma membrane adhesion molecules | CDH6 | cadherin 6 | −2.002 | 1.5 × 10−5 |
| CDH10 | cadherin 10 | −1.853 | 2.5 × 10−4 | |
| FAT3 | FAT atypical cadherin 3 | −2.201 | 3.3 × 10−4 | |
| Micro-RNAs | MIR206 | ----- | −2.574 | 4.1 × 10−4 |
| MIR214 | ----- | −1.757 | 4.5 × 10−4 | |
| MIR208B | ----- | −1.525 | 5.9 × 10−4 | |
| MIR130A | ----- | 2.862 | 2.0 × 10−4 | |
| Positive Regulation of Cell Proliferation | FGF17 | fibroblast growth factor 17 | 2.496 | 1.2 × 10−8 |
| RASGRP4 | RAS guanylyl releasing protein 4 | 1.743 | 2.9 × 10−5 | |
| Germ-line stem cell population maintenance | PIWIL2 | piwi-like RNA-mediated gene silencing 2 | 2.502 | 1.1 × 10−6 |
| GO Categories Enriched in H3K27ac from Satellite Cells Treated with Tributyrin | ||||
|---|---|---|---|---|
| GO Term | Associated Gene ID | Gene Name/Function | Fold Change | p-Value |
| Regulation of Cell Proliferation | DMNT1 | DNA (cytosine-5-)-methyltransferase | −1.732 | 1.2 × 10−4 |
| ATP Binding | MYO5A | myosin VA | −3.563 | 3.8 × 10−5 |
| PLK2 | polo like kinase 2 | −2.695 | 9.1 × 10−7 | |
| EGFR | epidermal growth factor receptor | −2.643 | 1.3 × 10−4 | |
| Calcium Ion Binding | SCUBE3 | signal peptide, CUB domain, EGF-like 3 | −2.407 | 1.3 × 10−4 |
| ANXA10 | annexin A10 | −2.233 | 4.2 × 10−5 | |
| ITPR3 | inositol 1,4,5-triphosphate receptor 3 | −2.022 | 2.7 × 10−5 | |
| Micro-RNAs | MIR128-1 | ----- | −2.360 | 1.4 × 10−4 |
| MIR181A-1 | ----- | 1.964 | 3.8 × 10−6 | |
| MIR210 | ----- | 2.922 | 5.7 × 10−7 | |
| Cell Adhesion Via Plasma Membrane | CDH7 | cadherin 7 | 2.819 | 5.6 × 10−5 |
| PCDH15 | protocadherin 15 | 3.133 | 4.6 × 10−7 | |
| TORC1 Complex | RPTOR | regulatory associated protein of MTOR, complex 1 | 1.916 | 9.8 × 10−7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Murray, R.L.; Zhang, W.; Liu, J.; Cooper, J.; Mitchell, A.; Buman, M.; Song, J.; Stahl, C.H. Tributyrin, a Butyrate Pro-Drug, Primes Satellite Cells for Differentiation by Altering the Epigenetic Landscape. Cells 2021, 10, 3475. https://doi.org/10.3390/cells10123475
Murray RL, Zhang W, Liu J, Cooper J, Mitchell A, Buman M, Song J, Stahl CH. Tributyrin, a Butyrate Pro-Drug, Primes Satellite Cells for Differentiation by Altering the Epigenetic Landscape. Cells. 2021; 10(12):3475. https://doi.org/10.3390/cells10123475
Chicago/Turabian StyleMurray, Robert L., Wei Zhang, Jianan Liu, Jason Cooper, Alex Mitchell, Maria Buman, Jiuzhou Song, and Chad H. Stahl. 2021. "Tributyrin, a Butyrate Pro-Drug, Primes Satellite Cells for Differentiation by Altering the Epigenetic Landscape" Cells 10, no. 12: 3475. https://doi.org/10.3390/cells10123475
APA StyleMurray, R. L., Zhang, W., Liu, J., Cooper, J., Mitchell, A., Buman, M., Song, J., & Stahl, C. H. (2021). Tributyrin, a Butyrate Pro-Drug, Primes Satellite Cells for Differentiation by Altering the Epigenetic Landscape. Cells, 10(12), 3475. https://doi.org/10.3390/cells10123475