
Drosophila melanogaster: A Model System to Study Distinct Genetic Programs in Myoblast Fusion


Abstract
:1. Introduction
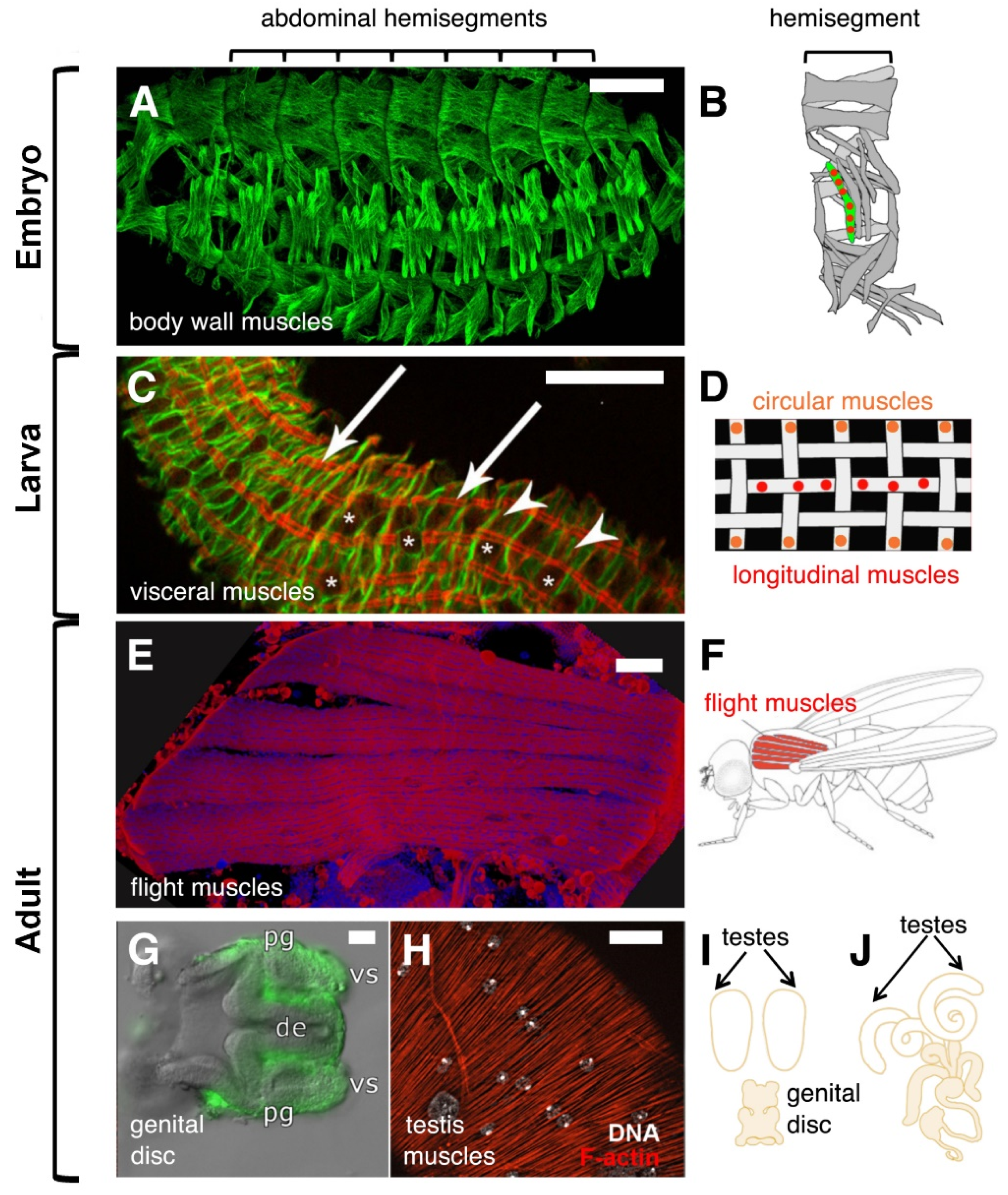
2. Muscle Formation during Drosophila Development
2.1. The Formation of the Larval Bodywall Muscles during Embryogenesis
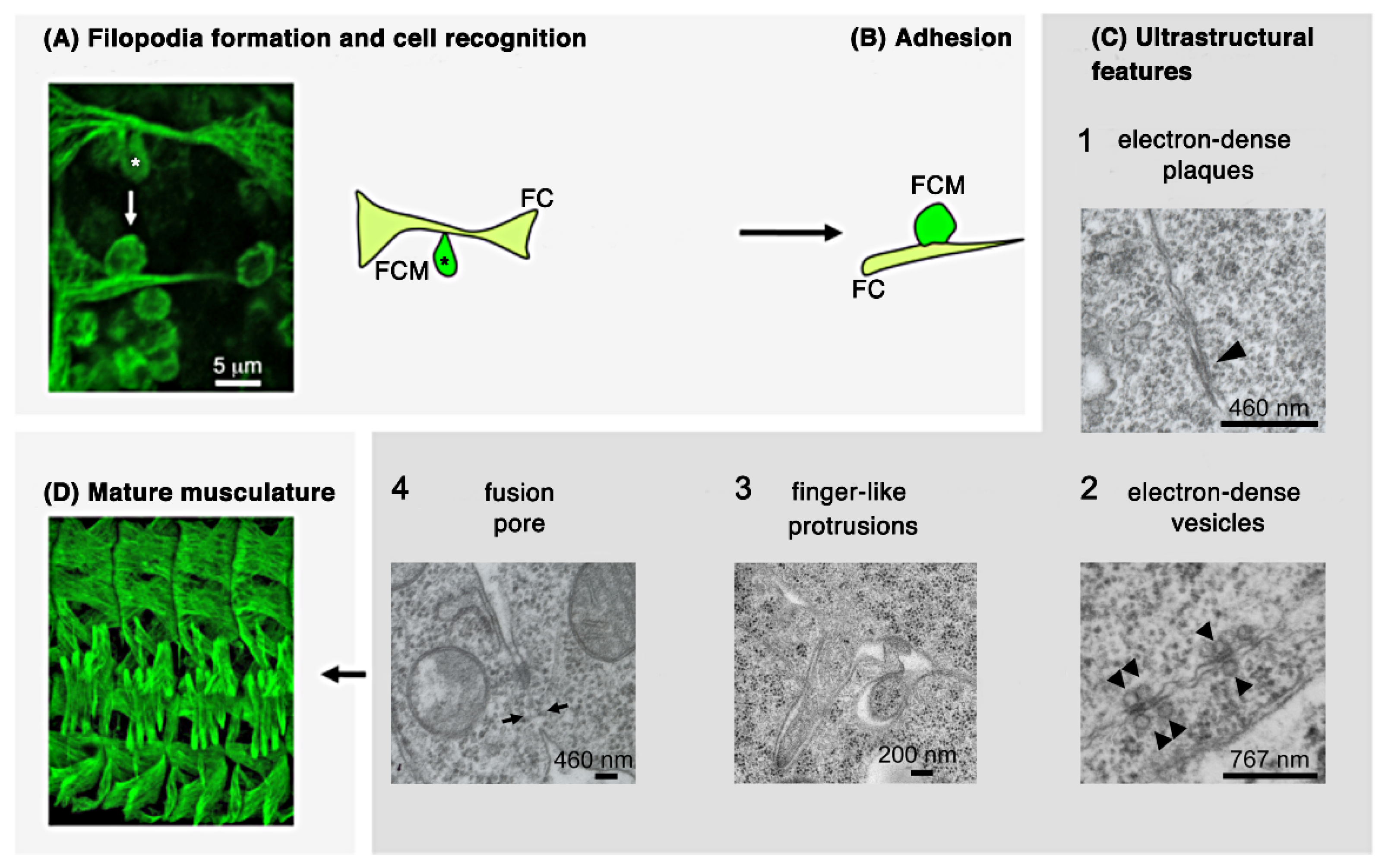
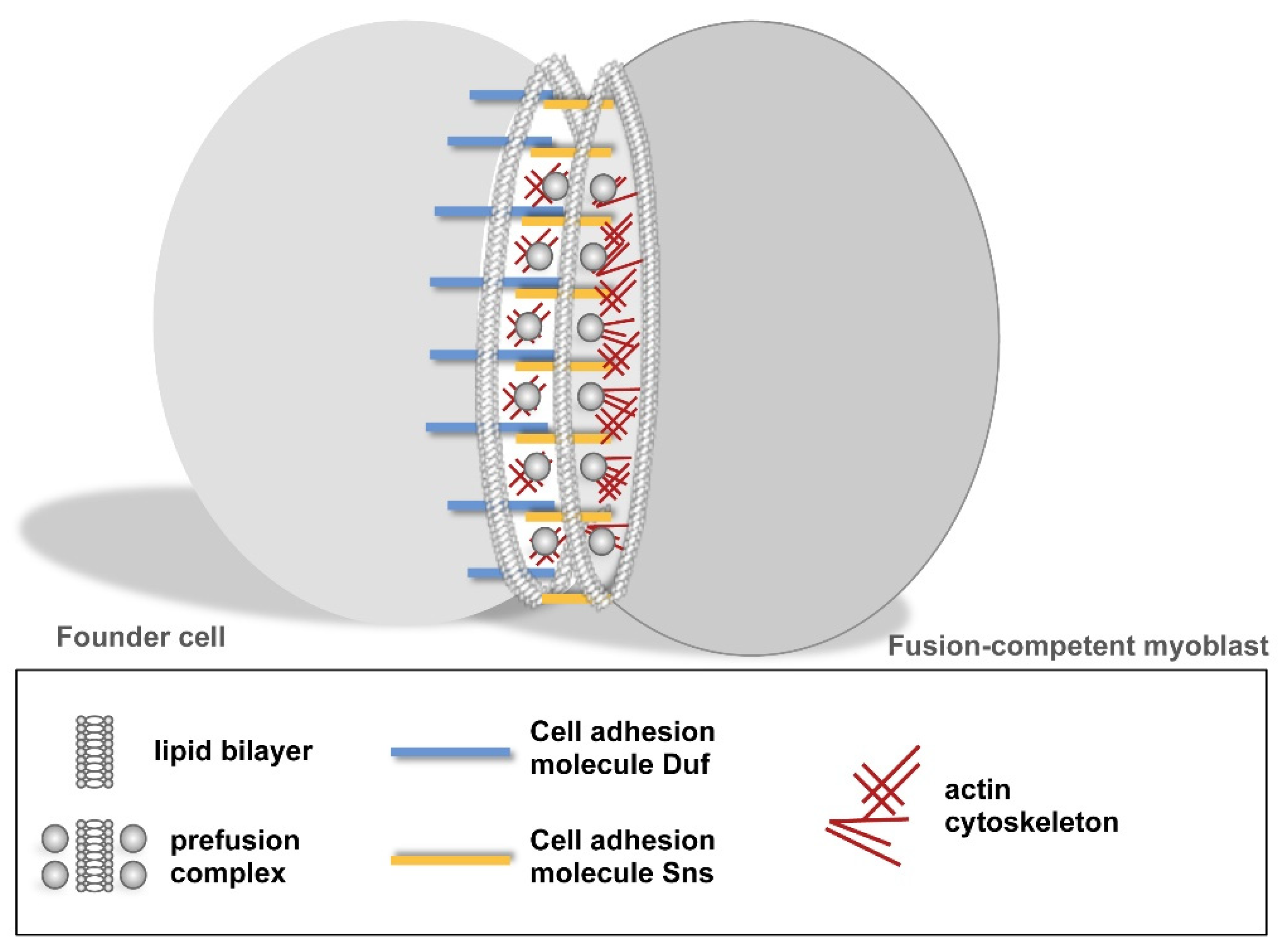
2.1.1. Founder Cells and Fusion Competent Myoblasts
2.1.2. Cellular Steps of Embryonic Myoblast Fusion
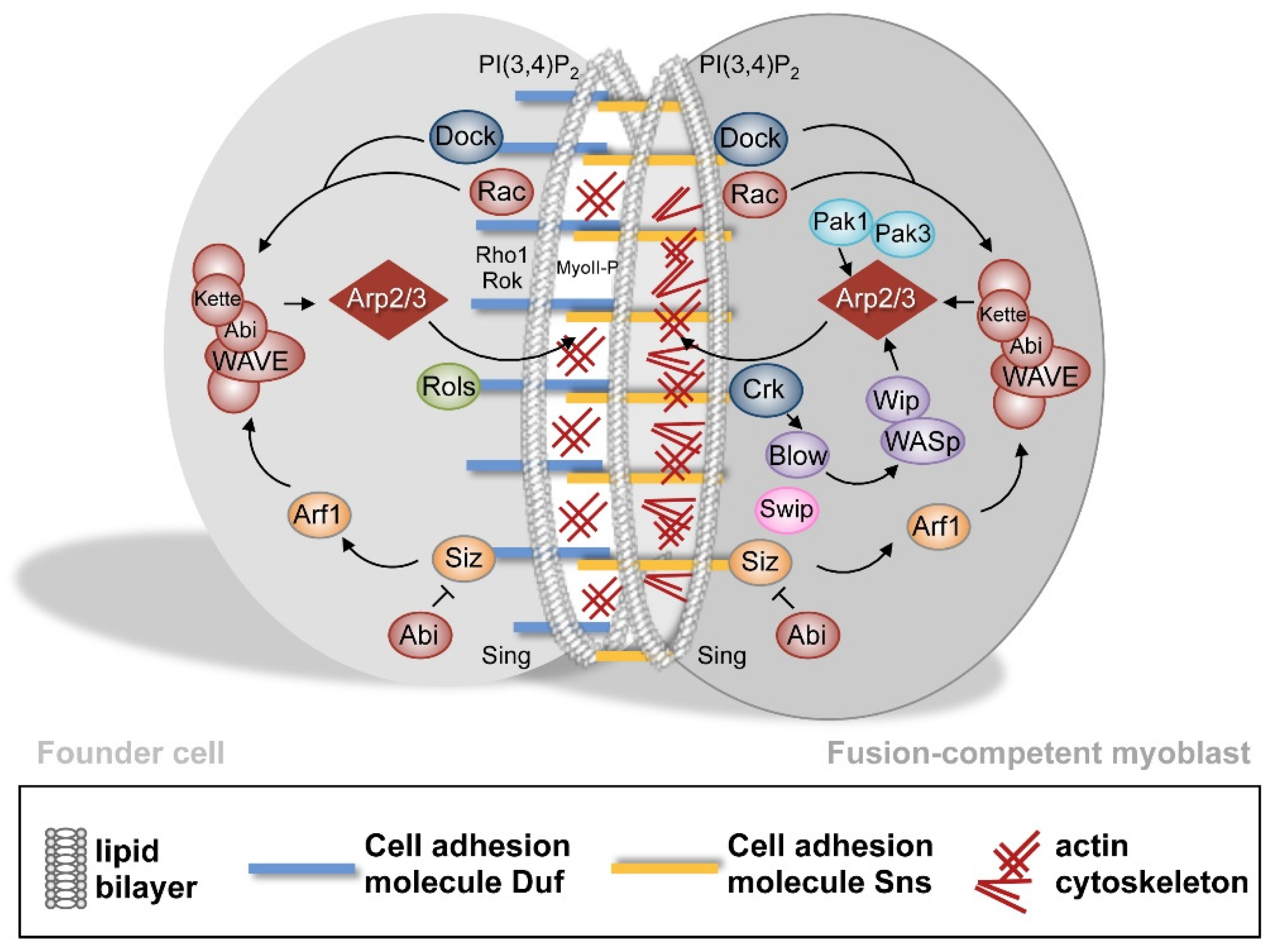
2.1.3. Establishment of a Cell–Cell Communication Structure after the Recognition and Adhesion of Myoblasts
2.1.4. Intracellular Signaling and Branched F-Actin Formations during Myoblast Fusion
3. Common and Different Players in Visceral and Flight Muscle Development
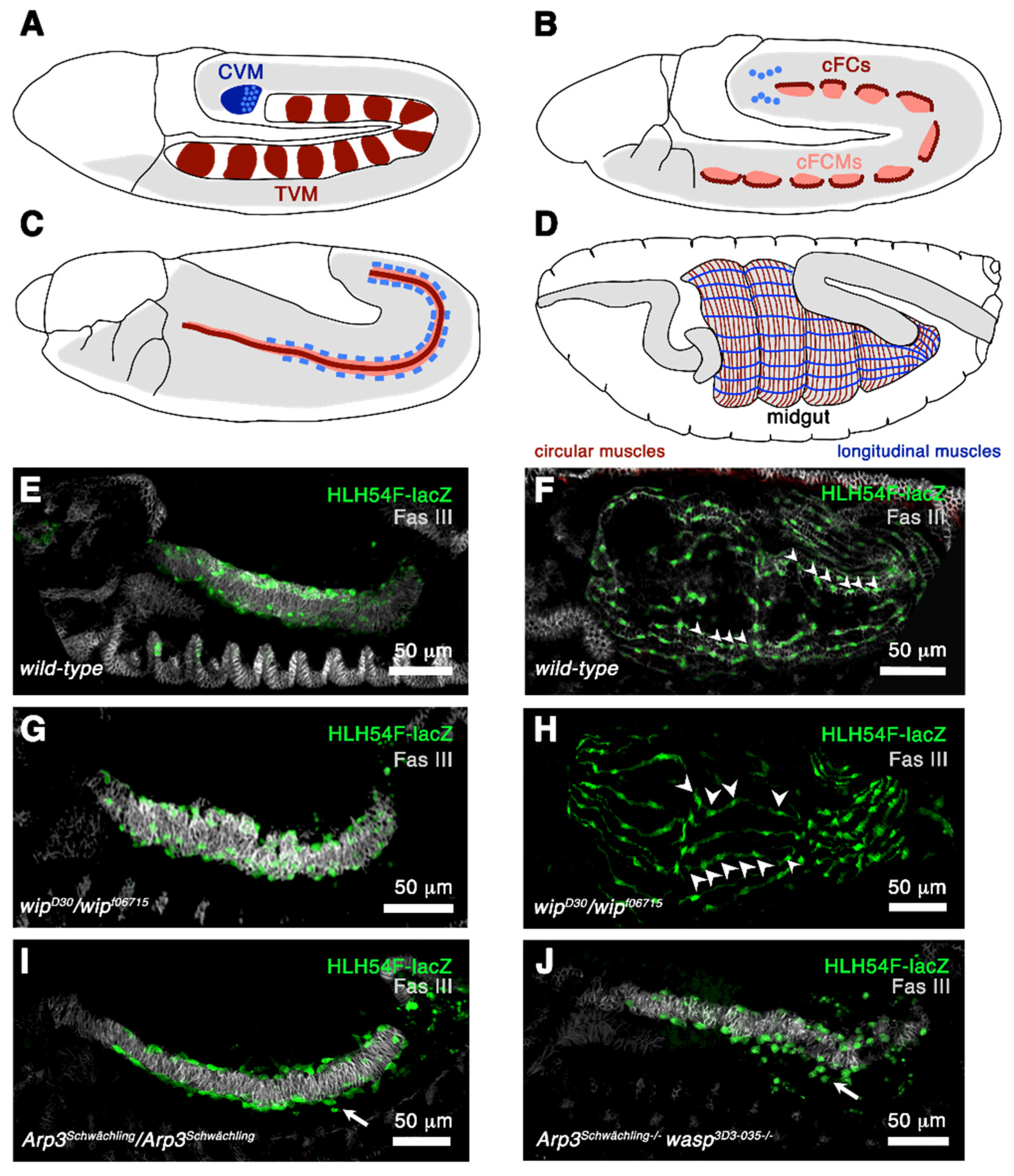
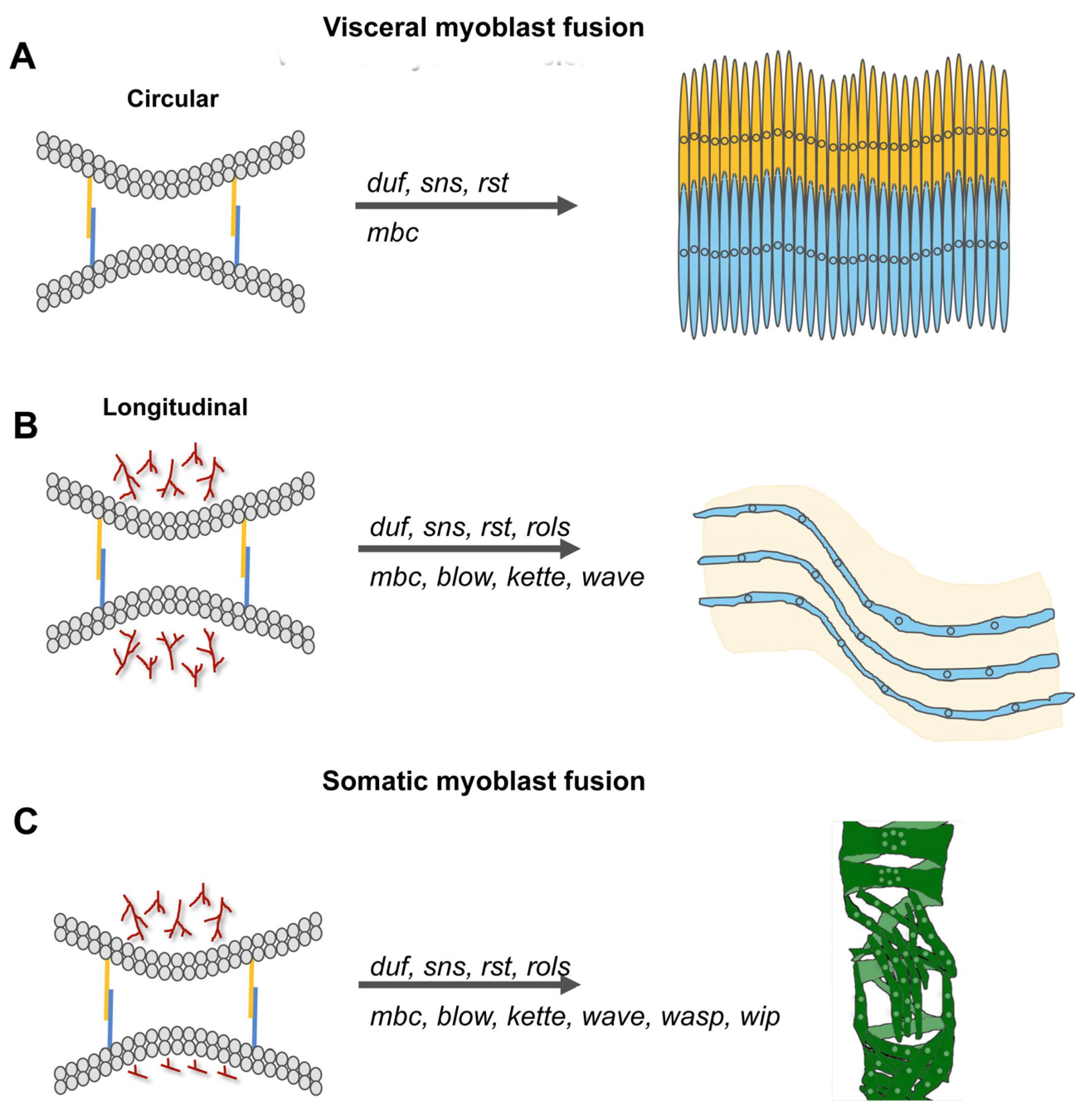
3.1. The Formation of the Visceral Muscles during Embryogenesis
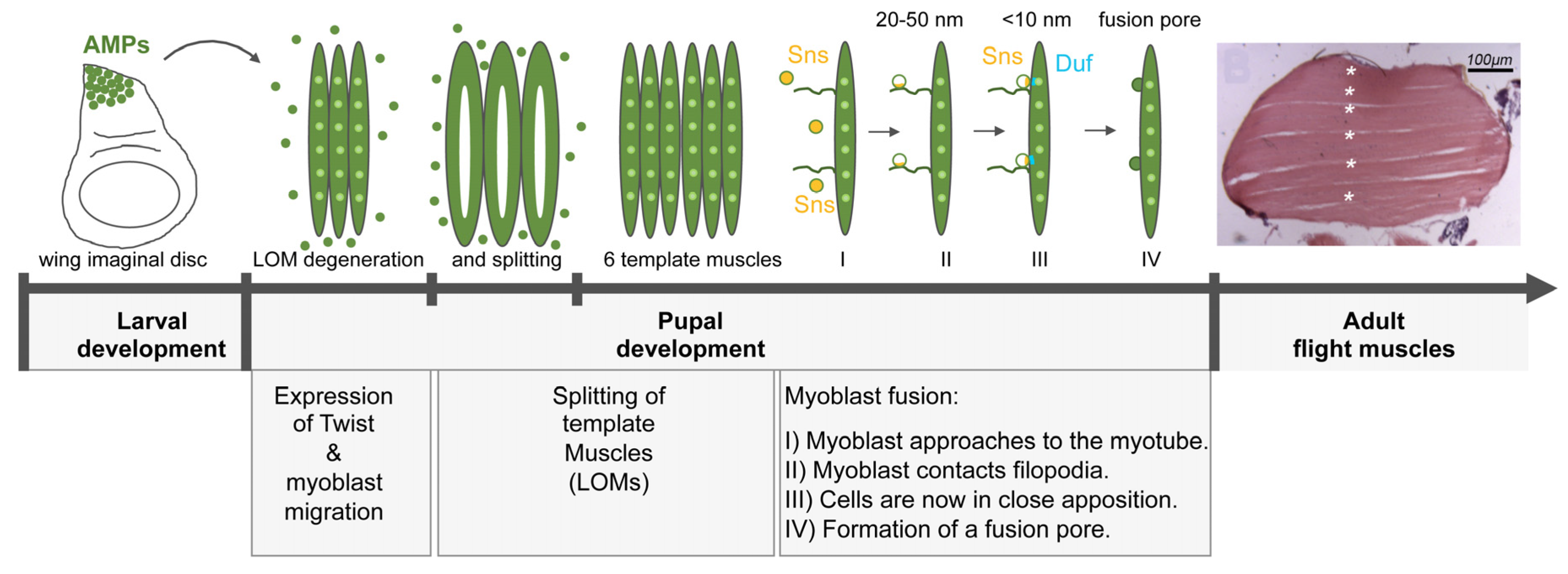
3.2. The Formation of Flight Muscles during Metamorphosis
3.3. Summary between Visceral and Flight Muscle Development
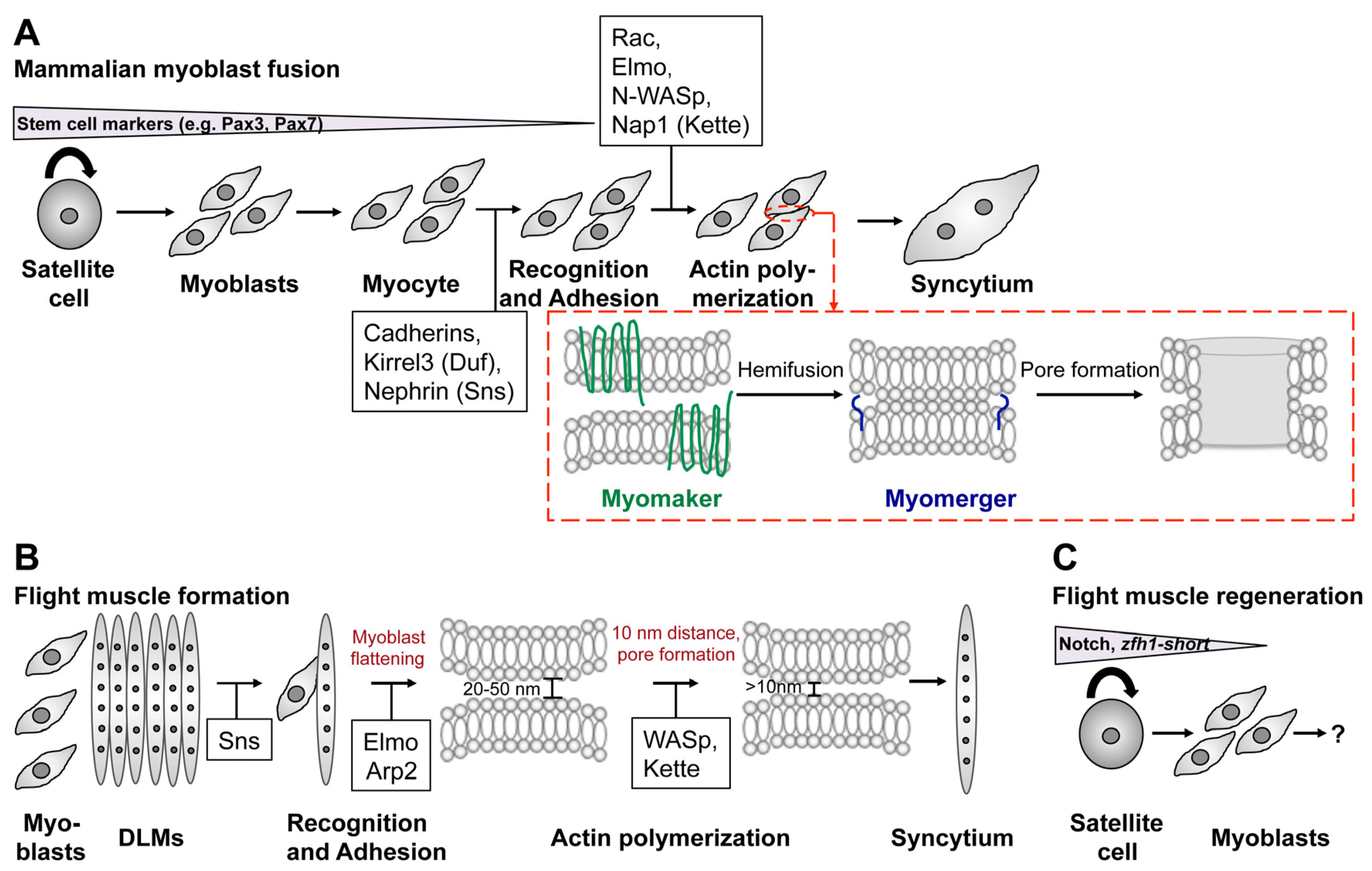
4. Myoblast Fusion in Mammals
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Rüegg, M.A.; Glass, D.J. Molecular mechanisms and treatment options for muscle wasting diseases. Annu. Rev. Pharmacol. Toxicol. 2011, 51, 373–395. [Google Scholar] [CrossRef] [PubMed]
- Ugur, B.; Chen, K.; Bellen, H.J. Drosophila tools and assays for the study of human diseases. Dis. Model. Mech. 2016, 9, 235244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozlov, M.M.; Chernomordik, L.V. Membrane tension and membrane fusion. Curr. Opin. Struct. Biol. 2015, 33, 61–67. [Google Scholar] [CrossRef] [Green Version]
- Hernández, J.M.; Podbilewicz, B. The hallmarks of cell-cell fusion. Development 2017, 144, 4481–4495. [Google Scholar] [CrossRef] [Green Version]
- Deng, S.; Azevedo, M.; Baylies, M. Acting on identity: Myoblast fusion and the formation of the syncytial muscle fiber. Semin. Cell Dev. Biol. 2017, 72, 45–55. [Google Scholar] [CrossRef]
- Kim, J.H.; Jin, P.; Duan, R.; Chen, E.H. Mechanisms of myoblast fusion during muscle development. Curr. Opin. Genet. Dev. 2015, 32, 162–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobi, K.C.; Schulman, V.K.; Baylies, M.K. Specification of the somatic musculature in Drosophila. Wiley Interdiscip. Rev. Dev. Biol. 2015, 4, 357–375. [Google Scholar] [CrossRef] [Green Version]
- Carmena, A.; Bate, M.; Jiménez, F. Lethal of scute, a proneural gene, participates in the specification of muscle progenitors during Drosophila embryogenesis. Genes Dev. 1995, 9, 2373–2383. [Google Scholar] [CrossRef] [Green Version]
- Artero, R.; Furlong, E.E.; Beckett, K.; Scott, M.P.; Baylies, M. Notch and Ras signaling pathway effector genes expressed in fusion competent and founder cells during Drosophila myogenesis. Development 2003, 130, 6257–6272. [Google Scholar] [CrossRef] [Green Version]
- Broadie, K.S.; Bate, M. The development of adult muscles in Drosophila: Ablation of identified muscle precursor cells. Development 1991, 113, 103–118. [Google Scholar] [CrossRef]
- Tixier, V.; Bataillé, L.; Jagla, K. Diversification of muscle types: Recent insights from Drosophila. Exp. Cell Res. 2010, 316, 3019–3027. [Google Scholar] [CrossRef]
- Duan, H.; Skeath, J.B.; Nguyen, H.T. Drosophila Lame duck, a novel member of the Gli superfamily, acts as a key regulator of myogenesis by controlling fusion-competent myoblast development. Development 2001, 128, 4489–4500. [Google Scholar] [CrossRef] [PubMed]
- Poovathumkadavil, P.; Jagla, K. Genetic Control of Muscle Diversification and Homeostasis: Insights from. Cells 2020, 9, 1543. [Google Scholar] [CrossRef] [PubMed]
- Kesper, D.A.; Stute, C.; Buttgereit, D.; Kreiskother, N.; Vishnu, S.; Fischbach, K.F.; Renkawitz-Pohl, R. Myoblast fusion in Drosophila melanogaster is mediated through a fusion-restricted myogenic-adhesive structure (FuRMAS). Dev. Dyn. 2007, 236, 404–415. [Google Scholar] [CrossRef] [PubMed]
- Richardson, B.; Beckett, K.; Baylies, M. Visualizing new dimensions in Drosophila myoblast fusion. Bioessays 2008, 30, 423–431. [Google Scholar] [CrossRef] [Green Version]
- Gildor, B.; Massarwa, R.; Shilo, B.Z.; Schejter, E.D. The SCAR and WASp nucleation-promoting factors act sequentially to mediate Drosophila myoblast fusion. EMBO Rep. 2009, 10, 1043–1050. [Google Scholar] [CrossRef] [Green Version]
- Doberstein, S.K.; Fetter, R.D.; Mehta, A.Y.; Goodman, C.S. Genetic analysis of myoblast fusion: Blown fuse is required for progression beyond the prefusion complex. J. Cell Biol. 1997, 136, 1249–1261. [Google Scholar] [CrossRef]
- Kim, S.; Shilagardi, K.; Zhang, S.L.; Hong, S.N.; Sens, K.L.; Bo, J.; Gonzalez, G.A.; Chen, E.H. A critical function for the actin cytoskeleton in targeted exocytosis of prefusion vesicles during myoblast fusion. Dev. Cell 2007, 12, 571–586. [Google Scholar] [CrossRef] [Green Version]
- Estrada, B.; Maeland, A.D.; Gisselbrecht, S.S.; Bloor, J.W.; Brown, N.H.; Michelson, A.M. The MARVEL domain protein, Singles Bar, is required for progression past the pre-fusion complex stage of myoblast fusion. Dev. Biol. 2007, 307, 328–339. [Google Scholar] [CrossRef] [Green Version]
- Berger, S.; Schäfer, G.; Kesper, D.A.; Holz, A.; Eriksson, T.; Palmer, R.H.; Beck, L.; Klämbt, C.; Renkawitz-Pohl, R.; Önel, S.F. WASP and SCAR have distinct roles in activating the Arp2/3 complex during myoblast fusion. J. Cell Sci. 2008, 121, 1303–1313. [Google Scholar] [CrossRef] [Green Version]
- Sens, K.L.; Zhang, S.L.; Jin, P.; Duan, R.; Zhang, G.F.; Luo, F.B.; Parachini, L.; Chen, E.H. An invasive podosome-like structure promotes fusion pore formation during myoblast fusion. J. Cell Biol. 2010, 191, 1013–1027. [Google Scholar] [CrossRef] [Green Version]
- Hamp, J.; Löwer, A.; Dottermusch-Heidel, C.; Beck, L.; Moussian, B.; Flötenmeyer, M.; Önel, S.F. Drosophila Kette coordinates myoblast junction dissolution and the ratio of Scar-to-WASp during myoblast fusion. J. Cell Sci. 2016, 129, 3426–3436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Önel, S.F.; Rust, M.B.; Jacob, R.; Renkawitz-Pohl, R. Tethering membrane fusion: Common and different players in myoblasts and at the synapse. J. Neurogenet. 2014, 28, 302–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haralalka, S.; Shelton, C.; Cartwright, H.N.; Guo, F.; Trimble, R.; Kumar, R.P.; Abmayr, S.M. Live imaging provides new insights on dynamic F-actin filopodia and differential endocytosis during myoblast fusion in Drosophila. PLoS ONE 2014, 9, e114126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, R.; Lee, D.M.; Jimah, J.R.; Gerassimov, N.; Yang, C.; Kim, S.; Luvsanjav, D.; Winkelman, J.; Mettlen, M.; Abrams, M.E.; et al. Dynamin regulates the dynamics and mechanical strength of the actin cytoskeleton as a multifilament actin-bundling protein. Nat. Cell Biol. 2020, 22, 674–688. [Google Scholar] [CrossRef]
- Ruiz-Gomez, M.; Coutts, N.; Price, A.; Taylor, M.V.; Bate, M. Drosophila dumbfounded: A myoblast attractant essential for fusion. Cell 2000, 102, 189–198. [Google Scholar] [CrossRef] [Green Version]
- Bour, B.A.; Chakravarti, M.; West, J.M.; Abmayr, S.M. Drosophila SNS, a member of the immunoglobulin superfamily that is essential for myoblast fusion. Genes Dev. 2000, 14, 1498–1511. [Google Scholar] [CrossRef]
- Dworak, H.A.; Charles, M.A.; Pellerano, L.B.; Sink, H. Characterization of Drosophila hibris, a gene related to human nephrin. Development 2001, 128, 4265–4276. [Google Scholar] [CrossRef]
- Önel, S.-F.; Dottermusch, C.; Sickmann, A.; Buttgereit, D.; Renkawitz-Pohl, R. Role of the Actin Cytoskeleton Within FuRMAS During Drosophila Myoblast Fusion and First Functionally Conserved Factors in Vertebrates. In Cell Fusions: Regulation and Control; Larsson, I., Ed.; Springer: Berlin/Heidelberg, Germany, 2011. [Google Scholar]
- Önel, S.F.; Renkawitz-Pohl, R. FuRMAS: Triggering Myoblast Fusion in Drosophila. Dev. Dyn. 2009, 238, 15131–15525. [Google Scholar] [CrossRef]
- Oakes, S.A. Mitochondria control calcium entry at the immunological synapse. Proc. Natl. Acad. Sci. USA 2007, 104, 15171–15172. [Google Scholar] [CrossRef] [Green Version]
- Dottermusch-Heidel, C.; Groth, V.; Beck, L.; Oenel, S.-F. The Arf-GEF Schizo/Loner regulates N-cadherin to induce fusion competence of Drosophila myoblasts. Dev. Biol. 2012, 368, 18–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lübke, S.; Braukmann, C.; Rexer, K.-H.; Cigoja, L.; Önel, S.F. The Abl-interactor Abi suppresses the function of the BRAG2 GEF family member Schizo. Open Biol. 2021. [Google Scholar] [CrossRef]
- Abmayr, S.M.; Pavlath, G.K. Myoblast fusion: Lessons from flies and mice. Development 2012, 139, 641–656. [Google Scholar] [CrossRef] [Green Version]
- Kaipa, B.R.; Shao, H.; Schäfer, G.; Trinkewitz, T.; Groth, V.; Liu, J.; Beck, L.; Bogdan, S.; Abmayr, S.M.; Önel, S.F. Dock mediates Scar- and WASp-dependent actin polymerization through interaction with cell adhesion molecules in founder cells and fusion-competent myoblasts. J. Cell Sci. 2013, 126, 360–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, P.; Duan, R.; Luo, F.; Zhang, G.; Hong, S.N.; Chen, E.H. Competition between Blown Fuse and WASP for WIP Binding Regulates the Dynamics of WASP-Dependent Actin Polymerization In Vivo. Dev. Cell 2011, 20, 623–638. [Google Scholar] [CrossRef] [Green Version]
- Schäfer, G.; Weber, S.; Holz, A.; Bogdan, S.; Schumacher, S.; Müller, A.; Renkawitz-Pohl, R.; Önel, S.F. The Wiskott-Aldrich syndrome protein (WASP) is essential for myoblast fusion in Drosophila. Dev. Biol. 2007, 304, 664–674. [Google Scholar] [CrossRef] [Green Version]
- Vijayakumar, V.; Monypenny, J.; Chen, X.J.; Machesky, L.M.; Lilla, S.; Thrasher, A.J.; Antón, I.M.; Calle, Y.; Jones, G.E. Tyrosine phosphorylation of WIP releases bound WASP and impairs podosome assembly in macrophages. J. Cell Sci. 2015, 128, 251–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massarwa, R.; Carmon, S.; Shilo, B.Z.; Schejter, E.D. WIP/WASp-based actin-polymerization machinery is essential for myoblast fusion in Drosophila. Dev. Cell 2007, 12, 557–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brinkmann, K.; Winterhoff, M.; Önel, S.F.; Schultz, J.; Faix, J.; Bogdan, S. WHAMY is a novel actin polymerase promoting myoblast fusion, macrophage cell motility and sensory organ development in Drosophila. J. Cell Sci. 2016, 129, 604–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollard, T.D. Regulation of actin filament assembly by Arp2/3 complex and formins. Annu. Rev. Biophys. Biomol. Struct. 2007, 36, 451–477. [Google Scholar] [CrossRef]
- Richardson, B.E.; Beckett, K.; Nowak, S.J.; Baylies, M.K. SCAR/WAVE and Arp2/3 are crucial for cytoskeletal remodeling at the site of myoblast fusion. Development 2007, 134, 4357–4367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollitt, A.Y.; Insall, R.H. WASP and SCAR/WAVE proteins: The drivers of actin assembly. J. Cell Sci. 2009, 122, 2575–2578. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Borek, D.; Padrick, S.B.; Gomez, T.S.; Metlagel, Z.; Ismail, A.M.; Umetani, J.; Billadeau, D.D.; Otwinowski, Z.; Rosen, M.K. Structure and control of the actin regulatory WAVE complex. Nature 2010, 468, 533–538. [Google Scholar] [CrossRef]
- Hakeda-Suzuki, S.; Ng, J.; Tzu, J.; Dietzl, G.; Sun, Y.; Harms, M.; Nardine, T.; Luo, L.; Dickson, B.J. Rac function and regulation during Drosophila development. Nature 2002, 416, 438–442. [Google Scholar] [CrossRef]
- Haralalka, S.; Shelton, C.; Cartwright, H.N.; Katzfey, E.; Janzen, E.; Abmayr, S.M. Asymmetric Mbc, active Rac1 and F-actin foci in the fusion-competent myoblasts during myoblast fusion in Drosophila. Development 2011, 138, 1551–1562. [Google Scholar] [CrossRef] [Green Version]
- Geisbrecht, E.R.; Haralalka, S.; Swanson, S.K.; Florens, L.; Washburn, M.P.; Abmayr, S.M. Drosophila ELMO/CED-12 interacts with Myoblast city to direct myoblast fusion and ommatidial organization. Dev. Biol. 2008, 314, 137–149. [Google Scholar] [CrossRef] [Green Version]
- Duan, R.; Jin, P.; Luo, F.; Zhang, G.; Anderson, N.; Chen, E.H. Group I PAKs function downstream of Rac to promote podosome invasion during myoblast fusion in vivo. J. Cell Biol. 2012, 199, 169–185. [Google Scholar] [CrossRef] [Green Version]
- Chong, C.; Tan, L.; Lim, L.; Manser, E. The mechanism of PAK activation. Autophosphorylation events in both regulatory and kinase domains control activity. J. Biol. Chem. 2001, 276, 17347–17353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bothe, I.; Deng, S.; Baylies, M. PI(4,5)P2 regulates myoblast fusion through Arp2/3 regulator localization at the fusion site. Development 2014, 141, 2289–2301. [Google Scholar] [CrossRef] [Green Version]
- Deng, S.; Bothe, I.; Baylies, M.K. The Formin Diaphanous Regulates Myoblast Fusion through Actin Polymerization and Arp2/3 Regulation. PLoS Genet. 2015, 11, e1005381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, R.; Kim, J.H.; Shilgardi, S.; Schiffhauer, E.S.; Lee, D.M.; Son, S.; Li, S.; Thomas, C.; Luo, T.; Flechter, D.A.; et al. Spectrin is a mechanoresponsive protein shaping fusogenic synapse architecture during myoblast fusion. Nat. Cell Biol. 2018, 20, 688–698. [Google Scholar] [CrossRef]
- Antonny, B.; Burd, C.; De Camilli, P.; Chen, E.; Daumke, O.; Faelber, K.; Ford, M.; Frolov, V.A.; Frost, A.; Hinshaw, J.E.; et al. Membrane fission by dynamin: What we know and what we need to know. EMBO J. 2016, 35, 2270–2284. [Google Scholar] [CrossRef]
- Seugnet, L.; Simpson, P.; Haenlin, M. Requirement for dynamin during notch signaling in Drosophila neurogenesis. Dev. Biol. 1997, 192, 585–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuang, M.-C.; Lin, S.-S.; Ohniwa, R.L.; Lee, G.-H.; Su, Y.-A.; Chang, Y.-C.; Tang, M.-J.; Liu, Y.-W. Tks5 and Dynamin-2 enhance actin bundle rigidity in invadosomes to promote myoblast fusion. J. Cell Biol. 2019, 218, 1670–1685. [Google Scholar] [CrossRef] [Green Version]
- Rau, A.; Buttgereit, D.; Holz, A.; Fetter, R.; Doberstein, S.K.; Paululat, A.; Staudt, N.; Skeath, J.; Michelson, A.M.; Renkawitz-Pohl, R. Rolling pebbles (rols) is required in Drosophila muscle precursors for recruitment of myoblasts for fusion. Development 2001, 128, 5061–5073. [Google Scholar] [CrossRef]
- Menon, S.D.; Chia, W. Drosophila rolling pebbles: A multidomain protein required for myoblast fusion that recruits D-titin in response to the myoblast attractant dumbfounded. Dev. Cell 2001, 1, 691–703. [Google Scholar] [CrossRef] [Green Version]
- Chen, E.H.; Olson, E.N. Antisocial, an intracellular adaptor protein, is required for myoblast fusion in Drosophila. Dev. Cell 2001, 1, 705–715. [Google Scholar] [CrossRef] [Green Version]
- Bulchand, S.; Menon, S.D.; George, S.E.; Chia, W. The intracellular domain of Dumbfounded affects myoblast fusion efficiency and interacts with Rolling pebbles and Loner. PLoS ONE 2010, 5, e9374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avirneni-Vadlamudi, U.; Galindo, K.A.; Endicott, T.R.; Paulson, V.; Cameron, S.; Galindo, R.L. Drosophila and mammalian models uncover a role for the myoblast fusion gene TANC1 in rhabdomyosarcoma. J. Clin. Investig. 2012, 122, 403–407. [Google Scholar] [CrossRef] [Green Version]
- Hornbruch-Freitag, C.; Griemert, B.; Buttgereit, D.; Renkawitz-Pohl, R. Drosophila Swiprosin-1/EFHD2 accumulates at the prefusion complex stage during Drosophila myoblast fusion. J. Cell Sci. 2011, 124, 3266–3278. [Google Scholar] [CrossRef] [Green Version]
- Estrada, B.; Choe, S.E.; Gisselbrecht, S.S.; Michaud, S.; Raj, L.; Busser, B.W.; Halfon, M.S.; Church, G.M.; Michelson, A.M. An integrated strategy for analyzing the unique developmental programs of different myoblast subtypes. PLoS Genet. 2006, 2, e16. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Pulido, L.; Martín-Belmonte, F.; Valencia, A.; Alonso, M.A. MARVEL: A conserved domain involved in membrane apposition events. Trends Biochem. Sci. 2002, 27, 599–601. [Google Scholar] [CrossRef]
- San Martin, B.; Bate, M. Hindgut visceral mesoderm requires an ectodermal template for normal development in Drosophila. Development 2001, 128, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Klapper, R.; Stute, C.; Schomaker, O.; Strasser, T.; Janning, W.; Renkawitz-Pohl, R.; Holz, A. The formation of syncytia within the visceral musculature of the Drosophila midgut is dependent on duf, sns and mbc. Mech Dev. 2002, 110, 85–96. [Google Scholar] [CrossRef]
- Englund, C.; Lorén, C.E.; Grabbe, C.; Varshney, G.K.; Deleuil, F.; Hallberg, B.; Palmer, R.H. Jeb signals through the Alk receptor tyrosine kinase to drive visceral muscle fusion. Nature 2003, 425, 512–516. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.H.; Norris, A.; Weiss, J.B.; Frasch, M. Jelly belly protein activates the receptor tyrosine kinase Alk to specify visceral muscle pioneers. Nature 2003, 425, 507–512. [Google Scholar] [CrossRef] [PubMed]
- Stute, C.; Schimmelpfeng, K.; Renkawitz-Pohl, R.; Palmer, R.H.; Holz, A. Myoblast determination in the somatic and visceral mesoderm depends on Notch signalling.g as well as on milliways (mili(Alk)) as receptor for Jeb signalling. Development 2004, 131, 743–754. [Google Scholar] [CrossRef] [Green Version]
- Ismat, A.; Schaub, C.; Reim, I.; Kirchner, K.; Schultheis, D.; Frasch, M. HLH54F is required for the specification and migration of longitudinal gut muscle founders from the caudal mesoderm of Drosophila. Development 2010, 137, 3107–3117. [Google Scholar] [CrossRef] [Green Version]
- Rudolf, A.; Buttgereit, D.; Jacobs, M.; Wolfstetter, G.; Kesper, D.; Pütz, M.; Berger, S.; Renkawitz-Pohl, R.; Holz, A.; Önel, S.F. Distinct genetic programs guide Drosophila circular and longitudinal visceral myoblast fusion. BMC Cell Biol. 2014, 15, 27. [Google Scholar] [CrossRef] [Green Version]
- Gunage, R.D.; Dhanyasi, N.; Reichert, H.; VijayRaghavan, K. Drosophila adult muscle development and regeneration. Semin Cell Dev. Biol. 2017, 72, 56–66. [Google Scholar] [CrossRef]
- Roy, S.; VijayRaghavan, K. Patterning muscles using organizers: Larval muscle templates and adult myoblasts actively interact to pattern the dorsal longitudinal flight muscles of Drosophila. J. Cell Biol. 1998, 141, 1135–1145. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, J.; Bate, M.; Vijayraghavan, K. Development of the indirect flight muscles of Drosophila. Development 1991, 113, 67–77. [Google Scholar] [CrossRef]
- Farrell, E.R.; Fernandes, J.; Keshishian, H. Muscle organizers in Drosophila: The role of persistent larval fibers in adult flight muscle development. Dev. Biol. 1996, 176, 220–229. [Google Scholar] [CrossRef]
- Fernandes, J.J.; Celniker, S.E.; VijayRaghavan, K. Development of the indirect flight muscle attachment sites in Drosophila: Role of the PS integrins and the stripe gene. Dev. Biol. 1996, 176, 166–184. [Google Scholar] [CrossRef] [Green Version]
- Gunage, R.D.; Reichert, H.; VijayRaghavan, K. Identification of a new stem cell population that generates Drosophila flight muscles. Elife 2014, 3, e03126. [Google Scholar] [CrossRef] [PubMed]
- Bernard, F.; Krejci, A.; Housden, B.; Adryan, B.; Bray, S.J. Specificity of Notch pathway activation: Twist controls the transcriptional output in adult muscle progenitors. Development 2010, 137, 2633–2642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elgar, S.J.; Han, J.; Taylor, M.V. mef2 activity levels differentially affect gene expression during Drosophila muscle development. Proc. Natl. Acad. Sci. USA 2008, 105, 918–923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segal, D.; Dhanyasi, N.; Schejter, E.D.; Shilo, B.Z. Adhesion and Fusion of Muscle Cells Are Promoted by Filopodia. Dev. Cell 2016, 38, 291–304. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, P.; Gildor, B.; Shilo, B.Z.; VijayRaghavan, K.; Schejter, E.D. The actin nucleator WASp is required for myoblast fusion during adult Drosophila myogenesis. Development 2011, 138, 2347–2357. [Google Scholar] [CrossRef] [Green Version]
- Dhanyasi, N.; Segal, D.; Shimoni, E.; Shinder, V.; Shilo, B.Z.; VijayRaghavan, K.; Schejter, E.D. Surface apposition and multiple cell contacts promote myoblast fusion in Drosophila flight muscles. J. Cell Biol. 2015, 211, 191–203. [Google Scholar] [CrossRef] [Green Version]
- Chaturvedi, D.; Reichert, H.; Gunage, R.D.; VijayRaghavan, K. Identification and functional characterization of muscle satellite cells in. Elife 2017, 6, e30107. [Google Scholar] [CrossRef]
- Boukhatmi, H.; Bray, S. A population of adult satellite-like cells in. Elife 2018, 7, e35954. [Google Scholar] [CrossRef]
- Chen, B.; You, W.; Wang, Y.; Shan, T. The regulatory role of myomaker and Myomixer-Myomerger-Minio in muscle development and regeneration. Cell. Mol. Life Sci. 2020, 77, 1551–1569. [Google Scholar] [CrossRef] [PubMed]
- Petrany, M.J.; Millay, D.P. Cell Fusion: Merging Membranes and Making Muscle. Trends Cell Biol. 2019, 29, 964–973. [Google Scholar] [CrossRef]
- Hindi, S.M.; Tajrishi, M.M.; Kumar, A. Signaling mechanism in mammalian myoblast fusion. Sci. Signal. 2013, 6, re2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millay, D.P.; O’Rourke, J.R.; Sutherland, L.B.; Bezprozvannaya, S.; Shelton, J.M.; Bassel-Duby, R.; Olson, E.N. Myomaker is a membrane activator of myoblast fusion and muscle formation. Nature 2013, 499, 301–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bi, P.; Ramirez-Martinez, A.; Li, H.; Cannavino, J.; McAnally, J.R.; Shelton, J.M.; Sánchez-Ortiz, E.; Bassel-Duby, R.; Olson, E.N. Control of muscle formation by the fusogenic micropeptide myomixer. Science 2017, 356, 323–327. [Google Scholar] [CrossRef] [Green Version]
- Quinn, M.E.; Goh, Q.; Kurosaka, M.; Gamage, D.G.; Petrany, M.J.; Prasad, V.; Millay, D.P. Myomerger induces fusion of non-fusogenic cells and is required for skeletal muscle development. Nat. Commun. 2017, 8, 15665. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Vashisht, A.A.; O’Rourke, J.; Corbel, S.Y.; Moran, R.; Romero, A.; Miraglia, L.; Zhang, J.; Durrant, E.; Schmedt, C.; et al. The microprotein Minion controls cell fusion and muscle formation. Nat. Commun. 2017, 8, 15664. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Wen, J.; Bigot, A.; Chen, J.; Shang, R.; Mouly, V.; Bi, P. Human myotube formation is determined by MyoD-Myomixer/Myomaker axis. Sci. Adv. 2020, 18, eabc4062. [Google Scholar] [CrossRef]
- Luo, W.; Li, E.; Nie, Q.; Zhang, X. Myomaker, regulated by MyoD and MyoG and miR-140-3p, promote schicken myoblast fusion. Int. J. Mol. Sci. 2015, 16, 26186–26201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castiglioni, I.; Caccia, R.; Garcia-Manteiga, J.M.; Ferri, G.; Caretti, G.; Molineris, I.; Nishioka, K.; Gabellini, D. The Trithorax protein Ash1L promotes myoblast fusion by activating Cdon expression. Nat. Commun. 2018, 28, 5026. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Embryo | Adult | |
|---|---|---|
| Longitudinal muscles | Dorsal longitudinal muscles | |
| Cell adhesion/Protein class | ||
| Kirre/Duf | required | required |
| Hbs | n.d. | required |
| Rst/IrreC | n.d. | required |
| Sing | n.d. | required |
| Sns | required | required |
| Signaling molecules/Protein class | ||
| Blow (antagonist of WASp) | FCM | n.d. |
| Elmo (GEF) | n.d. | required |
| Mbc (GEF) | required | n.d. |
| Rols | required | n.d. |
| Actin regulators/Protein class | ||
| Arp3 | required | n.d. |
| Arp2 | n.d. | required |
| Kette | required | required |
| Scar/WAVE | required | n.d. |
| WASp | Not required | required |
| Wip/Sltr | Not required | required |
| Cell adhesion Proteins | ADAM12, CD36, M-cadherin, N-cadherin, Disintegrin, α3-Integrin, α9-Integrin, β-Integrin, Kirrel3 (Duf homologue), Neogenin, Nephrin (Sns homologue) |
| Transmembrane Lipids | Cholesterol, Phosphatidyl serine |
| Signaling Proteins | Arf6, Brag-2, β-Catenin, Creatine kinase B, Crk, Crkl, diacylglycerol kinase ξ, Dock1, Dock5, EB3, focal adhesion kinase, Kindlin-2, Myoferlin, Stab2, Syntropin Trio |
| Actin Proteins | BAI1, BAI3, Cdc42, Filamin C, Nap1/Hem, Non-muscle myosin 2A, Rac1, N-WASP, Wip |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rout, P.; Preußner, M.; Önel, S.F. Drosophila melanogaster: A Model System to Study Distinct Genetic Programs in Myoblast Fusion. Cells 2022, 11, 321. https://doi.org/10.3390/cells11030321
Rout P, Preußner M, Önel SF. Drosophila melanogaster: A Model System to Study Distinct Genetic Programs in Myoblast Fusion. Cells. 2022; 11(3):321. https://doi.org/10.3390/cells11030321
Chicago/Turabian StyleRout, Pratiti, Mathieu Preußner, and Susanne Filiz Önel. 2022. "Drosophila melanogaster: A Model System to Study Distinct Genetic Programs in Myoblast Fusion" Cells 11, no. 3: 321. https://doi.org/10.3390/cells11030321
APA StyleRout, P., Preußner, M., & Önel, S. F. (2022). Drosophila melanogaster: A Model System to Study Distinct Genetic Programs in Myoblast Fusion. Cells, 11(3), 321. https://doi.org/10.3390/cells11030321