
Adolescent THC Treatment Does Not Potentiate the Behavioral Effects in Adulthood of Maternal Immune Activation


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
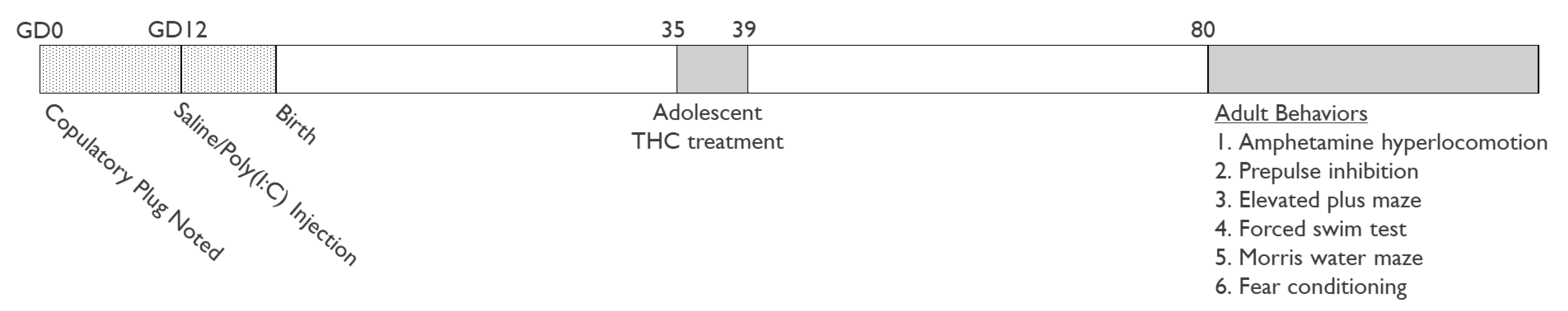
2.1. Animals
2.2. Drugs and Treatment
2.2.1. Maternal Immune Activation
2.2.2. THC Treatment
2.3. Behavioral Assays
2.3.1. Behavior in the Open Field and Amphetamine Hyperlocomotion
2.3.2. Prepulse Inhibition of the Acoustic Startle Reflex
2.3.3. Morris Water Maze (MWM)
2.3.4. Fear Conditioning and Extinction
2.3.5. Elevated Plus Maze
2.3.6. Forced Swim Test
2.4. Statistical Analyses
3. Results
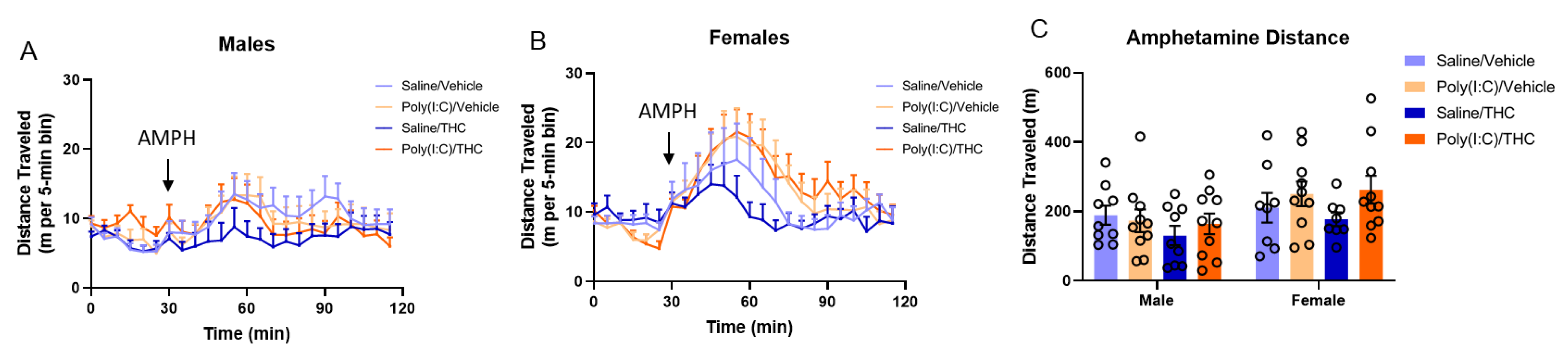
3.1. Amphetamine Hyperlocomotion
3.2. Prepulse Inhibition of the Acoustic Startle Response
3.3. Morris Water Maze
3.4. Fear Conditioning and Extinction
3.5. Elevated Plus Maze and Forced Swim Test
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brown, A.S. The environment and susceptibility to schizophrenia. Prog. Neurobiol. 2011, 93, 23–58. [Google Scholar] [CrossRef] [Green Version]
- Mednick, S.A.; Machon, R.A.; Huttunen, M.O.; Bonett, D. Adult Schizophrenia Following Prenatal Exposure to an Influenza Epidemic. Arch. Gen. Psychiatry 1988, 45, 189–192. [Google Scholar] [CrossRef]
- Barr, C.E.; Mednick, S.A.; Munk-Jorgensen, P. Exposure to influenza epidemics during gestation and adult schizophrenia. A 40-year study. Arch. Gen. Psychiatry 1990, 47, 869–874. [Google Scholar] [CrossRef]
- Adams, W.; Kendell, R.E.; Hare, E.H.; Munk-Jørgensen, P. Epidemiological Evidence that Maternal Influenza Contributes to the Aetiology of Schizophrenia. Br. J. Psychiatry 1993, 163, 522–534. [Google Scholar] [CrossRef]
- Izumoto, Y.; Inoue, S.; Yasuda, N. Schizophrenia and the influenza epidemics of 1957 in Japan. Biol. Psychiatry 1999, 46, 119–124. [Google Scholar] [CrossRef]
- Suvisaari, J.; Haukka, J.; Tanskanen, A.; Hovi, T.; Lönnqvist, J. Association between prenatal exposure to poliovirus infection and adult schizophrenia. Am. J. Psychiatry 1999, 156, 1100–1102. [Google Scholar] [CrossRef] [PubMed]
- Buka, S.L.; Tsuang, M.T.; Torrey, E.F.; Klebanoff, M.A.; Bernstein, D.; Yolken, R.H. Maternal Infections and Subsequent Psychosis Among Offspring. Arch. Gen. Psychiatry 2001, 58, 1032–1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, A.S.; Derkits, E.J. Prenatal Infection and Schizophrenia: A Review of Epidemiologic and Translational Studies. Am. J. Psychiatry 2010, 167, 261–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, A.S.; Meyer, U. Maternal Immune Activation and Neuropsychiatric Illness: A Translational Research Perspective. Am. J. Psychiatry 2018, 175, 1073–1083. [Google Scholar] [CrossRef] [Green Version]
- Estes, M.L.; McAllister, A.K. Maternal immune activation: Implications for neuropsychiatric disorders. Science 2016, 353, 772–777. [Google Scholar] [CrossRef] [Green Version]
- Meyer, U.; Feldon, J.; Schedlowski, M.; Yee, B.K. Towards an immuno-precipitated neurodevelopmental animal model of schizophrenia. Neurosci. Biobehav. Rev. 2005, 29, 913–947. [Google Scholar] [CrossRef] [PubMed]
- Meyer, U. Prenatal Poly(I:C) Exposure and Other Developmental Immune Activation Models in Rodent Systems. Biol. Psychiatry 2014, 75, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Van Der Steur, S.J.; Batalla, A.; Bossong, M.G. Factors Moderating the Association between Cannabis Use and Psychosis Risk: A Systematic Review. Brain Sci. 2020, 10, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marconi, A.; Di Forti, M.; Lewis, C.; Murray, R.; Vassos, E. Meta-analysis of the Association Between the Level of Cannabis Use and Risk of Psychosis. Schizophr. Bull. 2016, 42, 1262–1269. [Google Scholar] [CrossRef] [PubMed]
- Renard, J.; Rosen, L.G.; Loureiro, M.; De Oliveira, C.; Schmid, S.; Rushlow, W.J.; LaViolette, S.R. Adolescent Cannabinoid Exposure Induces a Persistent Sub-Cortical Hyper-Dopaminergic State and Associated Molecular Adaptations in the Prefrontal Cortex. Cereb. Cortex 2016, 27, 1297–1310. [Google Scholar] [CrossRef] [Green Version]
- McCutcheon, R.A.; Reis Marques, T.; Howes, O.D. Schizophrenia—An Overview. JAMA Psychiatry 2019, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zhong, L.; Brown, J.; Kramer, A.; Kaleka, K.; Petersen, A.; Krueger, J.N.; Florence, M.; Muelbl, M.J.; Battle, M.; Murphy, G.G.; et al. Increased Prefrontal Cortex Neurogranin Enhances Plasticity and Extinction Learning. J. Neurosci. 2015, 35, 7503–7508. [Google Scholar] [CrossRef]
- Braff, D.L.; Geyer, M.A. Sensorimotor gating and schizophrenia. Human and animal model studies. Arch. Gen. Psychiatry 1990, 47, 181–188. [Google Scholar] [CrossRef]
- Meyer, U.; Nyffeler, M.; Yee, B.K.; Knuesel, I.; Feldon, J. Adult brain and behavioral pathological markers of prenatal immune challenge during early/middle and late fetal development in mice. Brain Behav. Immun. 2008, 22, 469–486. [Google Scholar] [CrossRef]
- Weber-Stadlbauer, U.; Richetto, J.; Zwamborn, R.A.J.; Slieker, R.C.; Meyer, U. Transgenerational modification of dopaminergic dysfunctions induced by maternal immune activation. Neuropsychopharmacology 2021, 46, 404–412. [Google Scholar] [CrossRef]
- Senst, L.; Baimoukhametova, D.; Sterley, T.-L.; Bains, J.S. Sexually dimorphic neuronal responses to social isolation. eLife 2016, 5, 5904. [Google Scholar] [CrossRef] [Green Version]
- Giovanoli, S.; Engler, H.; Engler, A.; Richetto, J.; Voget, M.; Willi, R.; Winter, C.; Riva, M.A.; Mortensen, P.B.; Feldon, J.; et al. Stress in Puberty Unmasks Latent Neuropathological Consequences of Prenatal Immune Activation in Mice. Science 2013, 339, 1095–1099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; van Praag, H. Maternal immune activation differentially impacts mature and adult-born hippocampal neurons in male mice. Brain Behav. Immun. 2015, 45, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Haddad, F.L.; Lu, L.; Baines, K.J.; Schmid, S. Sensory filtering disruption caused by poly I:C—Timing of exposure and other experimental considerations. Brain Behav. Immun. Health 2020, 9, 100156. [Google Scholar] [CrossRef] [PubMed]
- Jaehne, E.J.; Chong, E.M.S.; Sbisa, A.; Gillespie, B.; Hill, R.; Gogos, A.; Buuse, M.V.D. TrkB agonist 7,8-dihydroxyflavone reverses an induced prepulse inhibition deficit selectively in maternal immune activation offspring: Implications for schizophrenia. Behav. Pharmacol. 2021, 32, 404–412. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, Y.; Inaba, H.; Iwakura, Y.; Namba, H.; Sotoyama, H.; Murata, Y.; Iwamoto, K.; Nawa, H. Inter-breeder differences in prepulse inhibition deficits of C57BL/6J mice in a maternal immune activation model. Neuropsychopharmacol. Rep. 2021, 41, 416–421. [Google Scholar] [CrossRef]
- Mueller, F.S.; Polesel, M.; Richetto, J.; Meyer, U.; Weber-Stadlbauer, U. Mouse models of maternal immune activation: Mind your caging system! Brain Behav. Immun. 2018, 73, 643–660. [Google Scholar] [CrossRef]
- Mueller, F.S.; Scarborough, J.; Schalbetter, S.M.; Richetto, J.; Kim, E.; Couch, A.; Yee, Y.; Lerch, J.P.; Vernon, A.C.; Weber-Stadlbauer, U.; et al. Behavioral, neuroanatomical, and molecular correlates of resilience and susceptibility to maternal immune activation. Mol. Psychiatry 2021, 26, 396–410. [Google Scholar] [CrossRef]
- Wilk, C.M. Brief cognitive assessment in schizophrenia: Normative data for the Repeatable Battery for the Assessment of Neuropsychological Status. Schizophr. Res. 2004, 70, 175–186. [Google Scholar] [CrossRef]
- Keefe, R.S.E.; Fenton, W.S. How Should DSM-V Criteria for Schizophrenia Include Cognitive Impairment? Schizophr. Bull. 2007, 33, 912–920. [Google Scholar] [CrossRef] [Green Version]
- Harvey, P.D.; Howanitz, E.; Parrella, M.; White, L.; Davidson, M.; Mohs, R.C.; Hoblyn, J.; Davis, K.L. Symptoms, Cognitive Functioning, and Adaptive Skills in Geriatric Patients With Lifelong Schizophrenia: A Comparison Across Treatment Sites. Am. J. Psychiatry 1998, 155, 1080–1086. [Google Scholar] [CrossRef]
- Vuillermot, S.; Luan, W.; Meyer, U.; Eyles, D. Vitamin D treatment during pregnancy prevents autism-related phenotypes in a mouse model of maternal immune activation. Mol. Autism 2017, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Weber-Stadlbauer, U.; Richetto, J.; Labouesse, M.; Bohacek, J.; Mansuy, I.M.; Meyer, U. Transgenerational transmission and modification of pathological traits induced by prenatal immune activation. Mol. Psychiatry 2016, 22, 102–112. [Google Scholar] [CrossRef]
- Labouesse, M.; Langhans, W.; Meyer, U. Abnormal context–reward associations in an immune-mediated neurodevelopmental mouse model with relevance to schizophrenia. Transl. Psychiatry 2015, 5, e637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vigli, D.; Palombelli, G.; Fanelli, S.; Calamandrei, G.; Canese, R.; Mosca, L.; Scattoni, M.L.; Ricceri, L. Maternal Immune Activation in Mice Only Partially Recapitulates the Autism Spectrum Disorders Symptomatology. Neuroscience 2020, 445, 109–119. [Google Scholar] [CrossRef]
- Sangha, S.; Greba, Q.; Robinson, P.D.; Ballendine, S.A.; Howland, J.G. Heightened fear in response to a safety cue and extinguished fear cue in a rat model of maternal immune activation. Front. Behav. Neurosci. 2014, 8, 168. [Google Scholar] [CrossRef] [Green Version]
- Yee, N.; Schwarting, R.K.; Fuchs, E.; Wöhr, M. Increased affective ultrasonic communication during fear learning in adult male rats exposed to maternal immune activation. J. Psychiatr. Res. 2012, 46, 1199–1205. [Google Scholar] [CrossRef] [PubMed]
- Zuckerman, L.; Weiner, I. Maternal immune activation leads to behavioral and pharmacological changes in the adult offspring. J. Psychiatr. Res. 2005, 39, 311–323. [Google Scholar] [CrossRef]
- Vorhees, C.V.; Graham, D.; Braun, A.; Schaefer, T.L.; Skelton, M.; Richtand, N.M.; Williams, M. Prenatal immune challenge in rats: Altered responses to dopaminergic and glutamatergic agents, prepulse inhibition of acoustic startle, and reduced route-based learning as a function of maternal body weight gain after prenatal exposure to poly IC. Synapse 2012, 66, 725–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poulia, N.; Delis, F.; Brakatselos, C.; Polissidis, A.; Koutmani, Y.; Kokras, N.; Dalla, C.; Politis, P.K.; Antoniou, K. Detrimental effects of adolescent escalating low-dose Δ(9) -tetrahydrocannabinol leads to a specific bio-behavioural profile in adult male rats. Br. J. Pharm. 2021, 178, 1722–1736. [Google Scholar] [CrossRef]
- Cha, Y.M.; White, A.M.; Kuhn, C.M.; Wilson, W.A.; Swartzwelder, H. Differential effects of delta9-THC on learning in adolescent and adult rats. Pharmacol. Biochem. Behav. 2006, 83, 448–455. [Google Scholar] [CrossRef]
- Cha, Y.M.; Jones, K.H.; Kuhn, C.M.; Wilson, W.A.; Swartzwelder, H.S. Sex differences in the effects of delta9-tetrahydrocannabinol on spatial learning in adolescent and adult rats. Behav Pharm. 2007, 18, 563–569. [Google Scholar] [CrossRef]
- Ruiz, C.M.; Torrens, A.; Castillo, E.; Perrone, C.R.; Cevallos, J.; Inshishian, V.C.; Harder, E.V.; Justeson, D.N.; Huestis, M.A.; Swarup, V.; et al. Pharmacokinetic, behavioral, and brain activity effects of Δ9-tetrahydrocannabinol in adolescent male and female rats. Neuropsychopharmacology 2021, 46, 959–969. [Google Scholar] [CrossRef] [PubMed]
- Torrens, A.; Vozella, V.; Huff, H.; McNeil, B.; Ahmed, F.; Ghidini, A.; Mahler, S.V.; Huestis, M.A.; Das, A.; Piomelli, D. Comparative Pharmacokinetics of Δ9-Tetrahydrocannabinol in Adolescent and Adult Male Mice. J. Pharmacol. Exp. Ther. 2020, 374, 151–160. [Google Scholar] [CrossRef]
- Schaefer, J.D.; Jang, S.-K.; Vrieze, S.; Iacono, W.G.; McGue, M.; Wilson, S. Adolescent cannabis use and adult psychoticism: A longitudinal co-twin control analysis using data from two cohorts. J. Abnorm. Psychol. 2021, 130, 691–701. [Google Scholar] [CrossRef] [PubMed]
- Karcher, N.R.; Barch, D.M.; Demers, C.H.; Baranger, D.A.A.; Heath, A.C.; Lynskey, M.T.; Agrawal, A. Genetic Predisposition vs Individual-Specific Processes in the Association Between Psychotic-like Experiences and Cannabis Use. JAMA Psychiatry 2019, 76, 87. [Google Scholar] [CrossRef]
- Nesvåg, R.; Reichborn-Kjennerud, T.; Gillespie, N.A.; Knudsen, G.P.; Bramness, J.G.; Kendler, K.S.; Ystrom, E. Genetic and Environmental Contributions to the Association Between Cannabis Use and Psychotic-Like Experiences in Young Adult Twins. Schizophr. Bull. 2016, 43, 644–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lecca, S.; Luchicchi, A.; Scherma, M.; Fadda, P.; Muntoni, A.L.; Pistis, M. Δ9-Tetrahydrocannabinol During Adolescence Attenuates Disruption of Dopamine Function Induced in Rats by Maternal Immune Activation. Front. Behav. Neurosci. 2019, 13, 202. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stollenwerk, T.M.; Hillard, C.J. Adolescent THC Treatment Does Not Potentiate the Behavioral Effects in Adulthood of Maternal Immune Activation. Cells 2021, 10, 3503. https://doi.org/10.3390/cells10123503
Stollenwerk TM, Hillard CJ. Adolescent THC Treatment Does Not Potentiate the Behavioral Effects in Adulthood of Maternal Immune Activation. Cells. 2021; 10(12):3503. https://doi.org/10.3390/cells10123503
Chicago/Turabian StyleStollenwerk, Todd M., and Cecilia J. Hillard. 2021. "Adolescent THC Treatment Does Not Potentiate the Behavioral Effects in Adulthood of Maternal Immune Activation" Cells 10, no. 12: 3503. https://doi.org/10.3390/cells10123503
APA StyleStollenwerk, T. M., & Hillard, C. J. (2021). Adolescent THC Treatment Does Not Potentiate the Behavioral Effects in Adulthood of Maternal Immune Activation. Cells, 10(12), 3503. https://doi.org/10.3390/cells10123503