CERI, CEFX, and CPI: Largely Improved Positive Controls for Testing Antigen-Specific T Cell Function in PBMC Compared to CEF


 ,
, 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. PBMC
2.2. CD4 and CD8 Depletion of PBMC
2.3. Positive Control Antigens
2.3.1. CEF
2.3.2. CEFX
2.3.3. CPI
2.3.4. CERI
2.3.5. Anti-CD3
2.4. Human Interferon-γ ImmunoSpot® Assay
2.5. Statistical Analysis of ImmunoSpot SFU Counts
3. Results
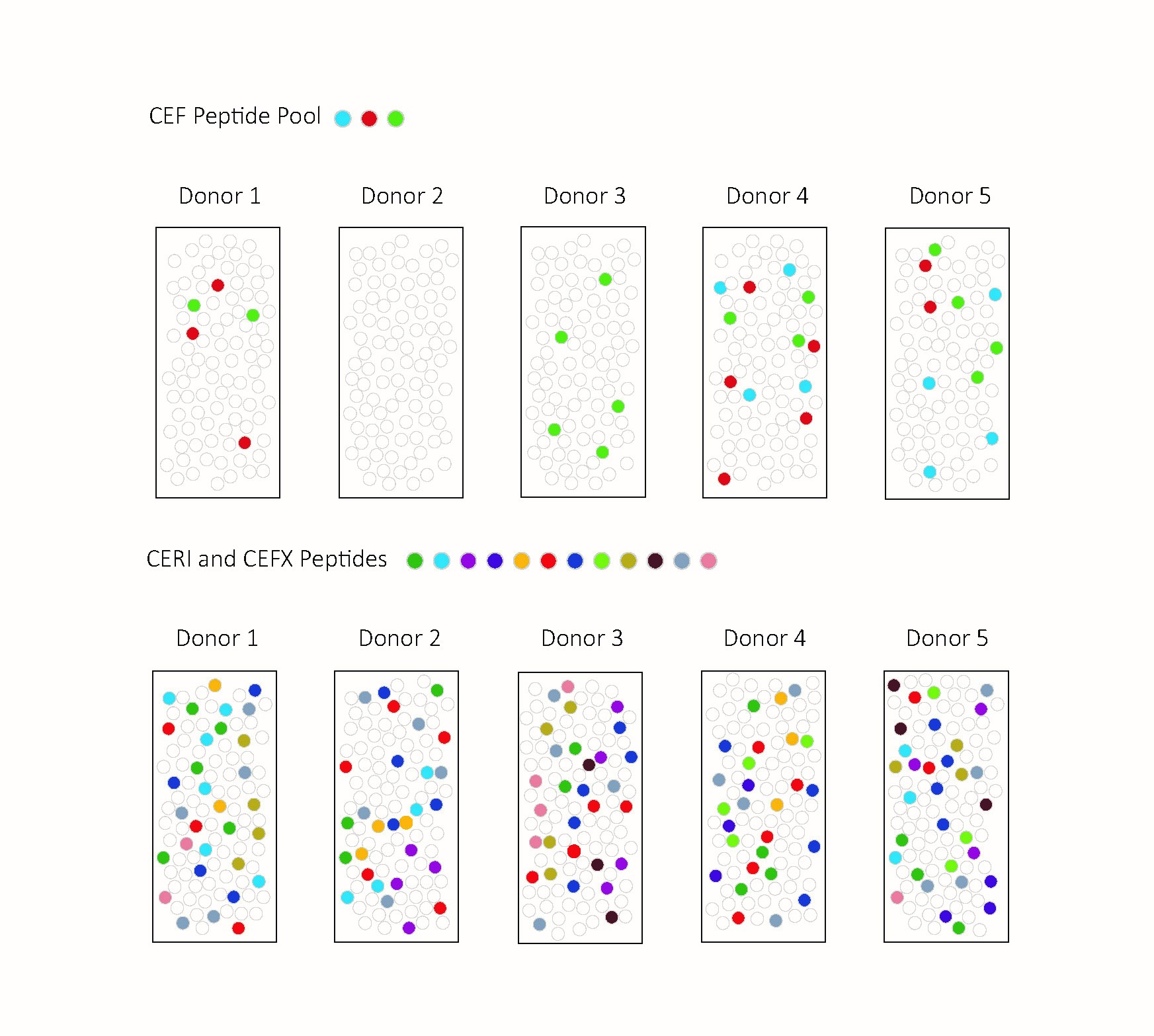
3.1. CEF and CERI Recall CD8+ T Cells, CPI Recalls CD4+ T Cells, and CEFX Recalls Both
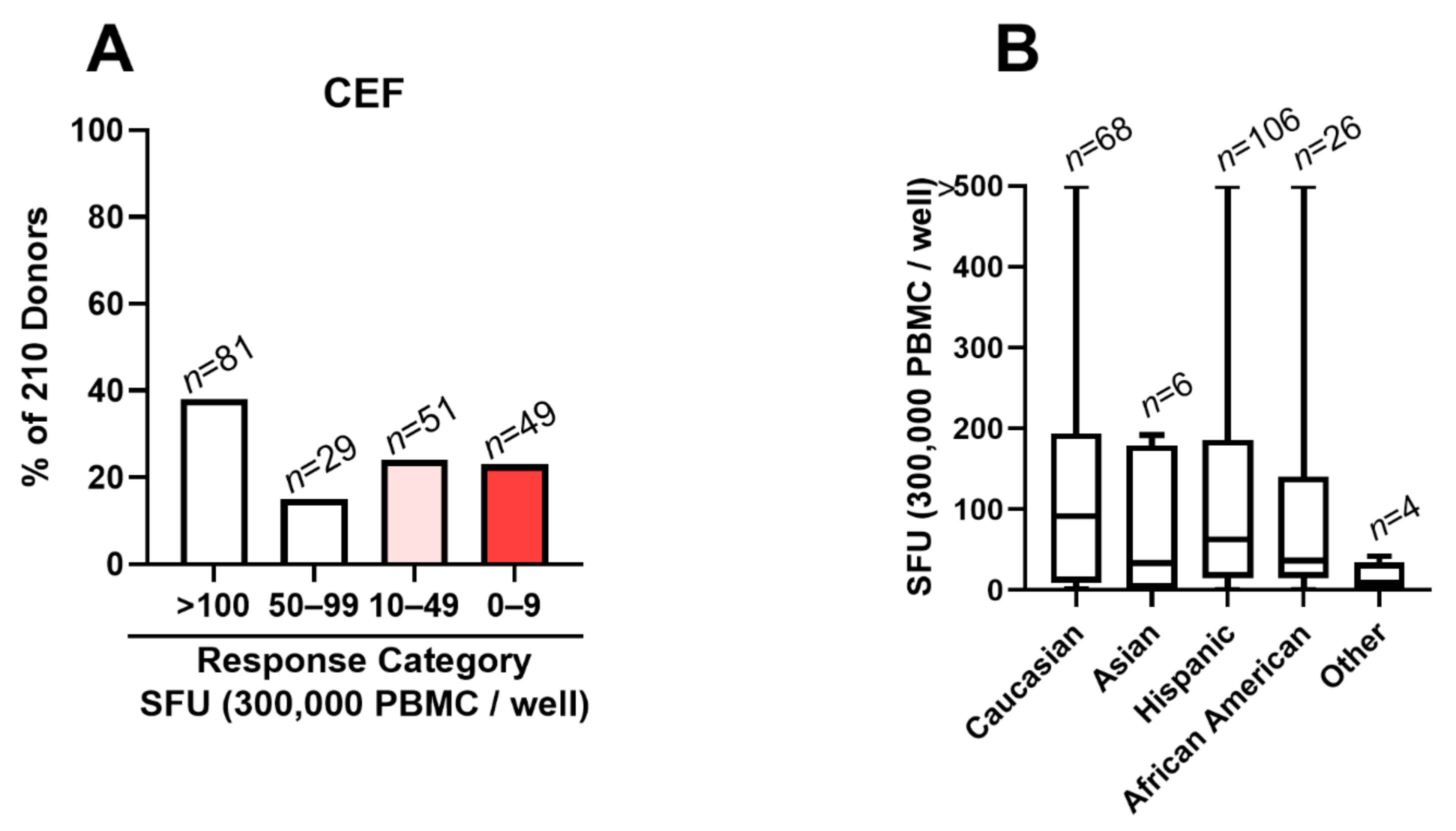
3.2. CEF Fails as a Positive Control in 48% of Test Subjects
3.3. CEF Non-Responder PBMCs Respond to Anti-CD3 Stimulation
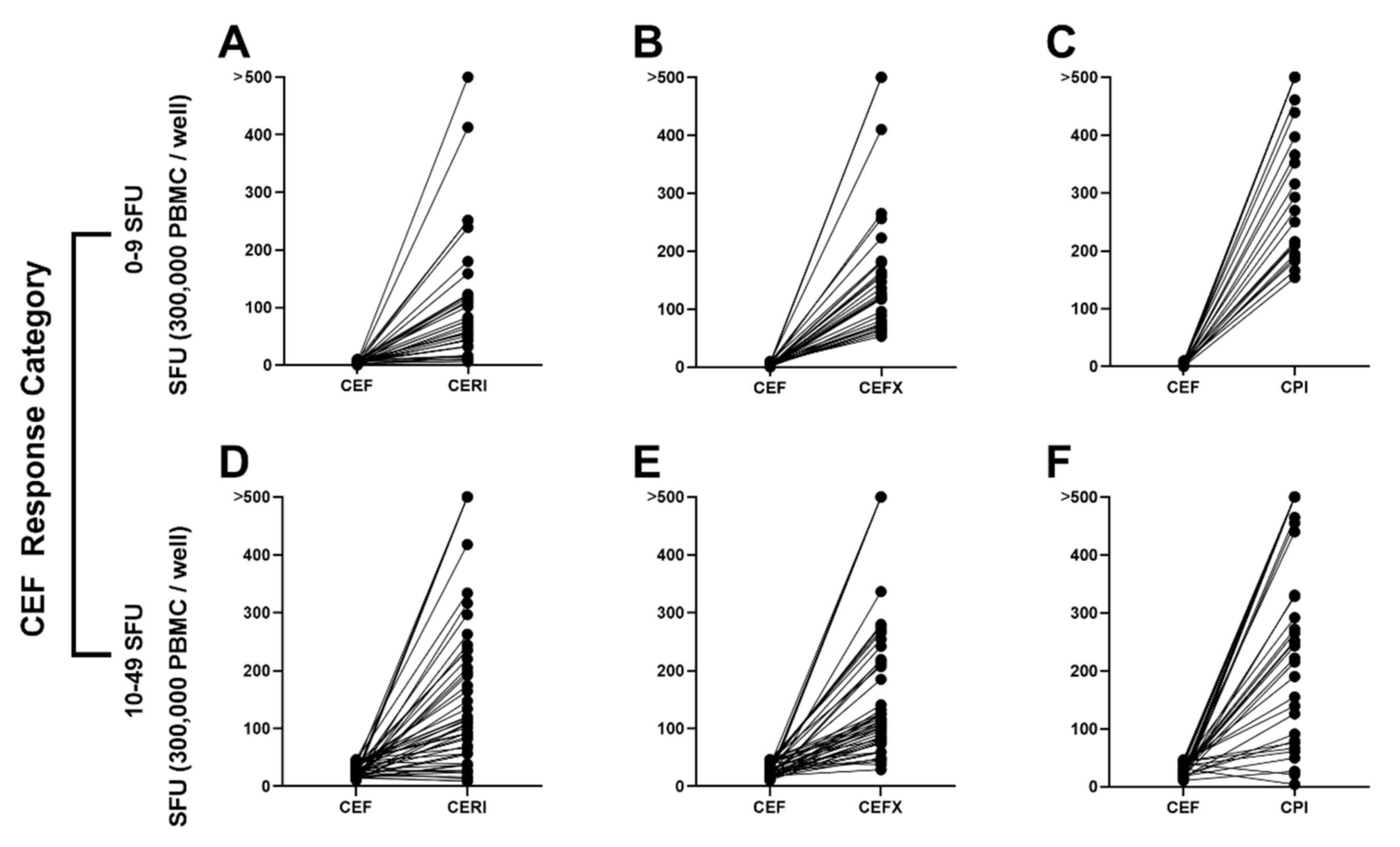
3.4. CEF Non-/Low-Responder PBMCs Respond to CERI, CEFX and CPI Stimulation
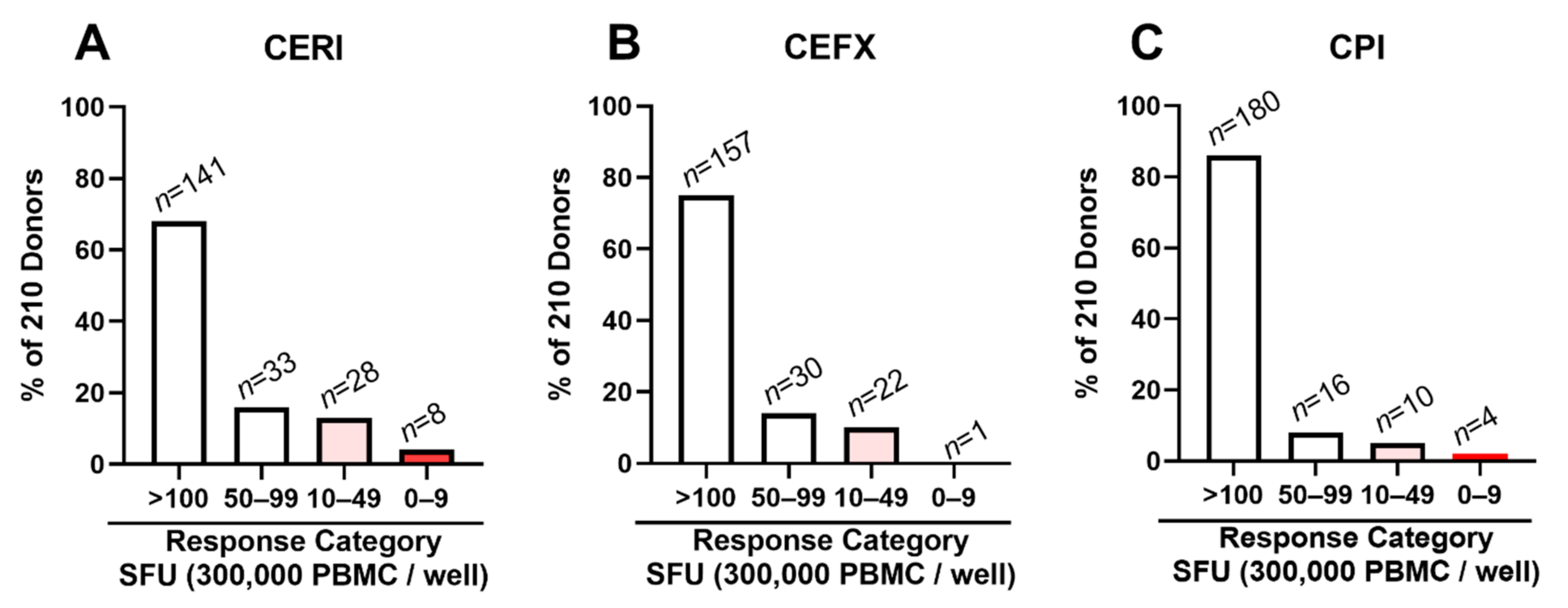
3.5. Low/Non-Responders to CERI, CEFX and CPI Are Rare
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tough, D.F.; Sprent, J. Life span of naive and memory T cells. Stem Cells 1995, 13, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Prabhakar, U.; Kelley, M. Validation of Cell-Based Assays in the GLP Setting: A Practical Guide; Wiley: Chichester, UK; John Wiley: Hoboken, NJ, USA, 2008. [Google Scholar]
- Kreher, C.R.; Dittrich, M.T.; Guerkov, R.; Boehm, B.O.; Tary-Lehmann, M. CD4+ and CD8+ cells in cryopreserved human PBMC maintain full functionality in cytokine ELISPOT assays. J. Immunol. Methods 2003, 278, 79–93. [Google Scholar] [CrossRef]
- Duechting, A.; Przybyla, A.; Kuerten, S.; Lehmann, P.V. Delayed Activation Kinetics of Th2- and Th17 Cells Compared to Th1 Cells. Cells 2017, 6, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramachandran, H.; Laux, J.; Moldovan, I.; Caspell, R.; Lehmann, P.V.; Subbramanian, R.A. Optimal thawing of cryopreserved peripheral blood mononuclear cells for use in high-throughput human immune monitoring studies. Cells 2012, 1, 313–324. [Google Scholar] [CrossRef]
- Neefjes, J.; Jongsma, M.L.; Paul, P.; Bakke, O. Towards a systems understanding of MHC class I and MHC class II antigen presentation. Nat. Rev. Immunol. 2011, 11, 823–836. [Google Scholar] [CrossRef]
- Pamer, E.; Cresswell, P. Mechanisms of MHC class I-restricted antigen processing. Annu. Rev. Immunol. 1998, 16, 323–358. [Google Scholar] [CrossRef]
- Reche, P.A.; Reinherz, E.L. Sequence variability analysis of human class I and class II MHC molecules: Functional and structural correlates of amino acid polymorphisms. J. Mol. Biol. 2003, 331, 623–641. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, P.V.; Lehmann, A.A. Aleatory epitope recognition prevails in human T cell responses? Crit Rev Immunol. 2020, 40, 225–235. [Google Scholar] [CrossRef]
- Currier, J.R.; Kuta, E.G.; Turk, E.; Earhart, L.B.; Loomis-Price, L.; Janetzki, S.; Ferrari, G.; Birx, D.L.; Cox, J.H. A panel of MHC class I restricted viral peptides for use as a quality control for vaccine trial ELISPOT assays. J. Immunol. Methods 2002, 260, 157–172. [Google Scholar] [CrossRef]
- NIH AIDS Reagent Program. CEF Control Peptide Pool NIH, Ed. 2018. Available online: aidreagent.org (accessed on 20 October 2020).
- Lehmann, A.A.; Zhang, T.; Reche, P.A.; Lehmann, P.V. Discordance between the predicted vs. the actually recognized CD8+ T cell epitopes of HCMV pp65 antigen and aleatory epitope dominance. Front. Immunol. 2021, in press. [Google Scholar] [CrossRef]
- Schiller, A.; Zhang, T.; Li, R.; Duechting, A.; Sundararaman, S.; Przybyla, A.; Kuerten, S.; Lehmann, P.V. A positive control for detection of functional CD4 T cells in PBMC: The CPI pool. Cells 2017, 6, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molero-Abraham, M.; Lafuente, E.M.; Flower, D.R.; Reche, P.A. Selection of conserved epitopes from hepatitis C virus for pan-populational stimulation of T-cell responses. Clin. Dev. Immunol. 2013, 2013, 601943. [Google Scholar] [CrossRef] [PubMed]
- Karulin, A.Y.; Karacsony, K.; Zhang, W.; Targoni, O.S.; Moldovan, I.; Dittrich, M.; Sundararaman, S.; Lehmann, P.V. ELISPOTs produced by CD8 and CD4 cells follow log normal size distribution permitting objective counting. Cells 2015, 4, 56–70. [Google Scholar] [CrossRef] [Green Version]
- Karulin, A.Y.; Caspell, R.; Dittrich, M.; Lehmann, P.V. Normal distribution of CD8+ T-cell-derived ELISPOT counts within replicates justifies the reliance on parametric statistics for identifying positive responses. Cells 2015, 4, 96–111. [Google Scholar] [CrossRef]
- Bjorkman, P.J.; Saper, M.A.; Samraoui, B.; Bennett, W.S.; Strominger, J.L.; Wiley, D.C. Structure of the human class I histocompatibility antigen, HLA-A2. Nature 1987, 329, 506–512. [Google Scholar] [CrossRef]
- Rock, K.L.; Reits, E.; Neefjes, J. Present yourself! By MHC class I and MHC class II molecules. Trends Immunol. 2016, 37, 724–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ott, P.A.; Tary-Lehmann, M.; Lehmann, P.V. The secretory IFN-gamma response of single CD4 memory cells after activation on different antigen presenting cell types. Clin. Immunol. 2007, 124, 267–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moldovan, I.; Targoni, O.; Zhang, W.; Sundararaman, S.; Lehmann, P.V. How frequently are predicted peptides actually recognized by CD8 cells? Cancer Immunol. Immunother. 2016, 65, 847–855. [Google Scholar] [CrossRef] [Green Version]
- Huppa, J.B.; Axmann, M.; Mörtelmaier, M.A.; Lillemeier, B.F.; Newell, E.W.; Brameshuber, M.; Klein, L.O.; Schütz, G.J.; Davis, M.M. TCR–peptide–MHC interactions in situ show accelerated kinetics and increased affinity. Nature 2010, 463, 963–967. [Google Scholar] [CrossRef] [Green Version]
- Norman, D.J. Mechanisms of action and overview of OKT3. Ther. Drug Monit. 1995, 17, 615–620. [Google Scholar] [CrossRef]
- Lehmann, P.V.; Suwansaard, M.; Zhang, T.; Roen, D.R.; Kirchenbaum, G.A.; Karulin, A.Y.; Lehmann, A.; Reche, P.A. Comprehensive evaluation of the expressed CD8+ T cell epitope space sing high-throughput epitope mapping. Front. Immunol. 2019, 10, 655. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subject | Media | CEF | CERI | CEFX | CPI | Anti-CD3 |
|---|---|---|---|---|---|---|
| ID 30 | 1 | 8 | 117 | 96 | 212 | >500 |
| ID 51 | 2 | 7 | 138 | 41 | 268 | >500 |
| ID 68 | 2 | 2 | 413 | 265 | 270 | >500 |
| ID 69 | 0 | 0 | 2 | 4 | 6 | 436 |
| ID 71 | 2 | 4 | 51 | 160 | 352 | >500 |
| ID 82 | 1 | 1 | 15 | 27 | 4 | 129 |
| ID 87 | 2 | 1 | 48 | 17 | 158 | >500 |
| ID 98 | 2 | 3 | 252 | 118 | 250 | >500 |
| ID 101 | 1 | 6 | 72 | 29 | 29 | 233 |
| ID 103 | 1 | 1 | 112 | 180 | 194 | >500 |
| ID 118 | 0 | 2 | 32 | 18 | 212 | 243 |
| ID 123 | 2 | 4 | 180 | 58 | 366 | 348 |
| ID 128 | 1 | 7 | 121 | 53 | 183 | 459 |
| ID 131 | 1 | 9 | 47 | 23 | 37 | 260 |
| ID 133 | 3 | 2 | 159 | 89 | 316 | >500 |
| ID 138 | 1 | 5 | 32 | 123 | 461 | >500 |
| ID 144 | 0 | 7 | 55 | 127 | 166 | >500 |
| ID 147 | 1 | 5 | 77 | 156 | >500 | >500 |
| ID 150 | 0 | 8 | 83 | 410 | 216 | >500 |
| ID 160 | 1 | 2 | 12 | 43 | 27 | 495 |
| ID 168 | 3 | 6 | 33 | 136 | 439 | >500 |
| ID 169 | 2 | 8 | 102 | 70 | 293 | 381 |
| ID 191 | 1 | 0 | 70 | 75 | 154 | 403 |
| ID 196 | 1 | 5 | 32 | 20 | 154 | >500 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lehmann, A.A.; Reche, P.A.; Zhang, T.; Suwansaard, M.; Lehmann, P.V. CERI, CEFX, and CPI: Largely Improved Positive Controls for Testing Antigen-Specific T Cell Function in PBMC Compared to CEF. Cells 2021, 10, 248. https://doi.org/10.3390/cells10020248
Lehmann AA, Reche PA, Zhang T, Suwansaard M, Lehmann PV. CERI, CEFX, and CPI: Largely Improved Positive Controls for Testing Antigen-Specific T Cell Function in PBMC Compared to CEF. Cells. 2021; 10(2):248. https://doi.org/10.3390/cells10020248
Chicago/Turabian StyleLehmann, Alexander A., Pedro A. Reche, Ting Zhang, Maneewan Suwansaard, and Paul V. Lehmann. 2021. "CERI, CEFX, and CPI: Largely Improved Positive Controls for Testing Antigen-Specific T Cell Function in PBMC Compared to CEF" Cells 10, no. 2: 248. https://doi.org/10.3390/cells10020248
APA StyleLehmann, A. A., Reche, P. A., Zhang, T., Suwansaard, M., & Lehmann, P. V. (2021). CERI, CEFX, and CPI: Largely Improved Positive Controls for Testing Antigen-Specific T Cell Function in PBMC Compared to CEF. Cells, 10(2), 248. https://doi.org/10.3390/cells10020248