
How Soon Hath Time… A History of Two “Seminal” Publications


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
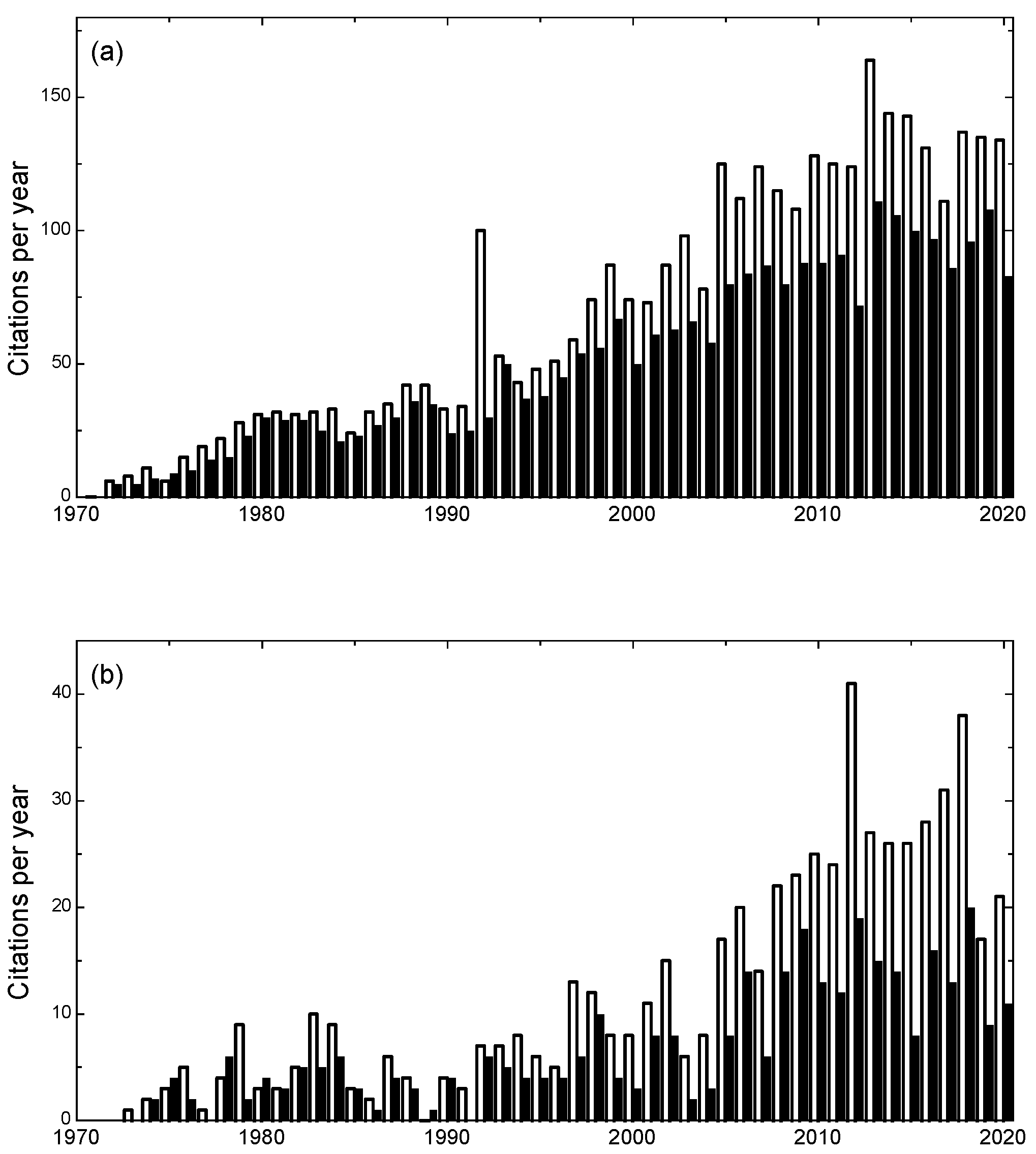
:1. Introduction
2. From Dung Flies to Sperm Competition, Anisogamy—And Other Things
Geoff Parker and I were very different characters, yet around 1964 we both found a lifetime’s inspiration from a single controversy. Outside of Bristol a war was raging in the world of evolutionary theory, triggered by V. C. Wynne-Edwards’ 1962 book extolling the power of group selection to bring about evolutionary change. People were taking sides, even in Bristol, with Geoff and I aligning with what seemed to be the minority at the time, the Individual Selectionists. In a bar or a refectory we would defend our cause against all-comers. We would even go to the then-extreme of claiming individual selection to be a force in the evolution of behavior. It was our first experience of the buzz to be gained from the defending of an academic idea that, apparently, was outrageous. At a time when few believed that behavior of any sort was even heritable, our backs were against the wall for much of the time.
…we shared a freezing apartment on the outskirts of Bristol. There, on those nights that we were both in, we would sit bent over the single gas fire, singeing gas-tainted toast on long forks and talking about sex—as postgraduates do. Unlike most postgraduates, our conversations revolved around dung-flies and Geoff’s ground-breaking notion of sperm competition. It is fun to think that for those few months that fire-lit room in Bristol could have been the only place in the world where sperm competition was being discussed…Or—another favorite sexual topic because it seemed so impenetrable—we would agonise over why there were two sexes, and only two. That question really stumped us, maybe even shook our faith a little. How could such a question possibly be answered by individual selection?
3. Sexual Selection and the Behavioural Ecology Revolution of the 1970s
Interesting to note that the 1:1 sex ratio…conflicts with the “best advantage to the species” philosophy. If…more females were produced, then reprod. [reproductive] rate of the species would increase.
This causes preponderance of males in competition for receptive females at any one time… [generating] often intense sexual selection on males.
May I say immediately that I have been extremely impressed with your paper on sexual selection…to my mind it is easily the most important contribution to the subject since Bateman’s work.
Your papers measuring opposing selection pressures for such traits as the passive [i.e., guarding] phase of males are a pleasure to read. As you know it’s extremely rare to find such measurements of opposing selection pressures. Someday, particularly for social traits, we will have to work out some more formal principles for applying natural selection than are commonly employed: you routinely think in terms of selection pressures operating simultaneously on several individuals at the same time, but this is not common, and it should be of value someday for someone to formulate in detail working rules by which one makes sophisticated functional arguments.
4. Sperm Competition
4.1. Writing and Publication
So far, [it] has been assumed that sexual selection concludes with the act of mating. This is an oversimplification…A better measure of sexual selection advantage is the fertilization rate in terms of the no. of offspring fertilized by a given male/unit time rather than no. of females inseminated/unit time.
Sperm competition may be defined as the competition, within a female, of the sperm from two or more males over the fertilization of that female’s ova.
I have attempted to analyse the intra-sexual selective pressures which arise in the insects as a result of the high level of sperm competition to which the group appears preadapted. Many copulatory and post-copulatory adaptations seem explicable in terms of this analysis. Previous considerations of sexual selection have tended to concentrate on pre-copulatory adaptations involving mechanisms by which males increase their chances of mating. I hope that the present review concerning sexual selection during and after mating will serve to link experimental genetics with the field behaviour approach.
…by bringing out the general principles perhaps more clearly…The treatment as it stands would be more suitable for a book…Will you please, without delay, see what you can do to comply with these ideas, and return the MS to me in Cambridge as soon as you possibly can. If we can get it to the printers in mid-July, there is a good chance that it will appear in the September number.
I hope you can do all this rather troublesome work without undue disruption—But I’m quite sure the referee is right in his comments, and in any case we are trying now to keep to the 20,000 word limit, because there are a very large number of reviews promised and space will have to be conserved.
I hope you will forgive me for not having retyped the rest…As time is limited I felt that these measures might be inadvisable.
I am thinking of making alterations and corrections as indicated on the enclosed sheet, and would like to have your blessing or veto.
4.2. Subsequent Developments
5. The Evolution of Gamete Dimorphism and Two Sexes: “PBS”
5.1. Writing and Publication
In a single memorable phone conversation with Geoff Parker, by then at Liverpool, I discovered that we had both dreamed-up a solution to our long-standing niggle at the conundrum of two sexes. The answer, we had both decided, had little to do with maleness and femaleness and everything to do with eggs and sperm. Explain the evolution of anisogamy, we encouraged each other, and everything else would fall into place.
Our referees have now returned the manuscript by you, Dr. Baker and Dr. Smith, which we are happy to accept. The paper has now been sent for publication.
5.2. Subsequent Developments
6. Final Thoughts
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Parker, G.A. Sperm competition and its evolutionary consequences in the insects. Biol. Rev. 1970, 45, 525–567. [Google Scholar] [CrossRef]
- Birkhead, T.R. How stupid not to have thought of that: Post-copulatory sexual selection. J. Zool. 2010, 281, 78–93. [Google Scholar] [CrossRef]
- Simmons, L.W.; Wedell, N. Fifty years of sperm competition: The structure of a scientific revolution. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2020, 375, 20200060. [Google Scholar] [CrossRef]
- Parker, G.A.; Baker, R.R.; Smith, V.G.F. The origin and evolution of gamete dimorphism and the male-female phenomenon. J. Theor. Biol. 1972, 36, 529–553. [Google Scholar] [CrossRef]
- Parker, G.A. Reflections before dusk. In Leaders in Animal Behavior: The Second Generation; Drickamer, L., Dewsbury, D.A., Eds.; Cambridge University press: Cambridge, UK, 2010; pp. 429–464. [Google Scholar]
- Parker, G.A. Golden flies, sunlit meadows: A tribute to the yellow dungfly. In Model Systems in Behavioural Ecology: Integrating Conceptual, Theoretical, and Empirical Approaches; Dugatkin, L.A., Ed.; Princeton University Press: Princeton, NJ, USA, 2001; pp. 3–26. [Google Scholar]
- Williams, G.C. Adaptation and Natural Selection: A Critique of Some Current Evolutionary Thought; Princeton University Press: Princeton, NJ, USA, 1966. [Google Scholar]
- Parker, G.A.; Geoff, A. Parker. Curr. Biol. 2007, 17, R111–R112. [Google Scholar] [CrossRef] [Green Version]
- Wynne-Edwards, V.C. Animal Dispersion in Relation to Social Behaviour; Oliver & Boyd: Edinburgh, Scotland, 1962. [Google Scholar]
- Baker, R.R. Foreword. In The Evolution of Sexuality; Shackelford, T.K., Hansen, R.D., Eds.; Springer: New York, NY, USA, 2015. [Google Scholar] [CrossRef]
- Dawkins, R. The Selfish Gene; Oxford University Press: Oxford, UK, 1976. [Google Scholar]
- Lehmann, L.; Keller, L.; West, S.; Roze, D. Group selection and kin selection: Two concepts but one process. Proc. Natl. Acad. Sci. USA 2007, 104, 6736. [Google Scholar] [CrossRef] [Green Version]
- Lehtonen, J. The Price equation and the unity of social evolution theory. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20190362. [Google Scholar] [CrossRef] [Green Version]
- Gardner, A. Price’s equation made clear. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20190361. [Google Scholar] [CrossRef] [Green Version]
- Baker, R.R. The Evolutionary Ecology of Animal Migration; Holmes & Meier Publishers Inc.: New York, NY, USA, 1978. [Google Scholar]
- Baker, R.R.; Bellis, M.A. Human Sperm Competition: Copulation, Masturbation and Infidelity; Chapman and Hall: London, UK, 1995. [Google Scholar]
- Baker, R.R. Robin Baker and Mark Bellis: Pioneers of research on human sperm competition. In Encyclopedia of Evolutionary Psychological Science; Shackelford, T.K., Weekes-Shackelford, V.A., Eds.; Springer International Publishing: New York, NY, USA, 2018. [Google Scholar] [CrossRef]
- Baker, R.R. Evolutionary psychology and the shaping of the novel Primal. Evol. Psychol. 2011, 9, 181–185. [Google Scholar] [CrossRef]
- Baker, R. Sexual whodunits and evolutionary psychology: The shaping of three novels. Evol. Psychol. 2013, 11, 147470491301100121. [Google Scholar] [CrossRef] [Green Version]
- Parker, G.A. In celebration of questions, past, present and future. In Social Behaviour: Genes, Ecology and Evolution, Székely, T., Moore, A.J., Komdeur, J., Eds.; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Parker, G.A. The reproductive behaviour and the nature of sexual selection in Scatophaga stercoraria L. (Diptera: Scatophagidae) II. The fertilization rate and the spatial and temporal relationships of each sex around the site of mating and oviposition. J. Anim. Ecol. 1970, 39, 205–228. [Google Scholar] [CrossRef]
- Parker, G.A. The reproductive behavior and nature of sexual selection in Scatophaga stercoraria L (Diptera-Scatophagidae). IX. Spatial distribution of fertilization rates and evolution of male search strategy within the reproductive area. Evolution 1974, 28, 93–108. [Google Scholar] [CrossRef]
- Parker, G.A. Sexual selection and sexual conflict. In Sexual Selection and Reproductive Competition in Insects; Blum, M.S., Blum, N.A., Eds.; Academic Press: New York, NY, USA, 1979; pp. 123–166. [Google Scholar]
- Parker, G.A. Foreword: A personal history of the development of animal contest theory and its role in the 1970s. In Animal Contests; Hardy, I.C.W., Briffa, M., Eds.; Cambridge University Press: Cambridge, UK, 2013; pp. Xi–Xx. [Google Scholar]
- Parker, G.A. The reproductive behaviour and the nature of sexual selection in Scatophaga stercoraria L. (Diptera: Scatophagidae) IV. Epigamic recognition and competition between males for the possession of females. Behaviour 1970, 37, 114–139. [Google Scholar]
- Parker, G.A. Sperm competition and its evolutionary effect on copula duration in the fly Scatophaga stercoraria. J. Insect. Physiol. 1970, 16, 1301–1328. [Google Scholar] [CrossRef]
- Parker, G.A. The reproductive behavior and the nature of sexual selection in Scatophaga stercoraria L. (Diptera: Scatophagidae). VII. The origin and evolution of the passive phase. Evolution 1970, 24, 774–788. [Google Scholar] [CrossRef] [Green Version]
- Parker, G.A.; Simmons, L.W.; Ward, P.I. Optimal copula duration in dungflies: Effects of frequency dependence and female mating status. Behav Ecol. Sociobiol. 1993, 32, 157–166. [Google Scholar] [CrossRef]
- Simmons, L.W.; Parker, G.A.; Hosken, D.J. Evolutionary insight from a humble fly: Sperm competition and the yellow dungfly. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2020, 375, 20200062. [Google Scholar] [CrossRef]
- Darwin, C.R. The Descent of Man, and Selection in Relation to Sex; John Murray: London, UK, 1871. [Google Scholar]
- Bateman, A.J. Intra-sexual selection in Drosophila. Heredity 1948, 2, 349–368. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, M.E. Studies on territorialism and sexual selection in dragonflies. Ecology 1955, 36, 566–586. [Google Scholar] [CrossRef]
- Parker, G.A. Behavioural ecology: Natural history as science. In Essays in Animal Behaviour: Celebrating 50 Years of Animal Behaviour; Lucas, J.R., Simmons, L.W., Eds.; Elsevier: Burlington, MA, USA, 2006; pp. 23–56. [Google Scholar]
- Darwin, C.R. On the Origin of Species by Means of Natural Selection; John Murray: London, UK, 1859. [Google Scholar]
- Parker, G.A.; Maynard Smith, J. Optimality theory in evolutionary biology. Nature 1990, 348, 27–33. [Google Scholar] [CrossRef]
- Maynard Smith, J. Optimization theory in evolution. Annu. Rev. Ecol. Syst. 1978, 9, 31–56. [Google Scholar] [CrossRef] [Green Version]
- Gowaty, P.A.; Hubbell, S.P. Chance, time allocation, and the evolution of adaptively flexible sex role behavior. Integr. Comp. Biol. 2005, 45, 931–944. [Google Scholar] [CrossRef] [PubMed]
- Trivers, R.L. Parental investment and sexual selection. In Sexual Selection and the Descent of Man, 1871–1971; Campbell, B., Ed.; Aldine: Chicago, IL, USA, 1972; pp. 136–179. [Google Scholar]
- Maynard Smith, J.; Price, G.R. The Logic of Animal Conflict. Nature 1973, 246, 15–18. [Google Scholar] [CrossRef]
- Andersson, M. Sexual Selection; Princeton University Press: Princeton, NJ, USA, 1994. [Google Scholar]
- Wallace, A.R. Darwinism: An Exposition of the Theory of Natural Selection with Some of its Applications; Macmillan and Co.: London, UK, 1889. [Google Scholar]
- Fisher, R.A. The Genetical Theory of Natural Selection; Oxford University Press: Oxford, UK, 1930. [Google Scholar]
- O’Donald, P. The theory of sexual selection. Heredity 1962, 17, 541. [Google Scholar] [CrossRef] [Green Version]
- Huxley, J. Darwin’s theory of sexual selection and the data subsumed by it, in the light of recent research. Am. Nat. 1938, 72, 416–433. [Google Scholar] [CrossRef]
- Huxley, J. The present standing of the theory of sexual selection. In Evolution: Essays on Aspects of Evolutionary Biology; de Beer, G.R., Ed.; Oxford University Press: Oxford, UK, 1938; pp. 11–42. [Google Scholar]
- Roughgarden, J. Sexual Selection: Is Anything Left? In Current Perspectives on Sexual Selection: What’s left after Darwin? Hoquet, T., Ed.; Springer: Dordrecht, The Netherlands, 2015; pp. 85–102. [Google Scholar] [CrossRef]
- Huxley, J. Evolution: The Modern Synthesis; George Allen and Unwin Ltd.: London, UK, 1942. [Google Scholar]
- Mayr, E. Sexual selection and natural selection. In Sexual Selection and the Descent of Man, 1871–1971; Campbell, B., Ed.; Aldine: Chicago, IL, USA, 1972; pp. 87–104. [Google Scholar]
- Hoquet, T. Bateman (1948): Rise and fall of a paradigm? Anim. Behav. 2020, 164, 223–231. [Google Scholar] [CrossRef]
- Parker, G.A.; Smith, J.L. Sperm competition and the evolution of the precopulatory passive phase behaviour in Locusta migratoria migratorioides. J. Entomol. Ser. A Gen. Entomol. 1975, 49, 155–171. [Google Scholar] [CrossRef]
- Waage, J.K. Dual function of the damselfly penis: Sperm removal and transfer. Science 1979, 203, 916–918. [Google Scholar] [CrossRef]
- Smith, R.L. Repeated copulation and sperm precedence: Paternity assurance for a male brooding water bug. Science 1979, 205, 1029–1031. [Google Scholar] [CrossRef]
- Sivinski, J.L. Sexual selection and insect sperm. Fla. Entomol. 1980, 63, 99–111. [Google Scholar] [CrossRef]
- Lloyd, J.E. Mating behavior and natural selection. Fla. Entomol. 1979, 62, 17–34. [Google Scholar] [CrossRef]
- Smith, R.L. (Ed.) Sperm Competition and the Evolution of Animal Mating Systems; Academic Press: New York, NY, USA, 1984. [Google Scholar]
- Thornhill, R. Cryptic female choice and its implications in the scorpionfly Harpobittacus nigriceps. Am. Nat. 1983, 122, 765–788. [Google Scholar] [CrossRef]
- Thornhill, R.; Alcock, J. The Evolution of Insect Mating Systems; Harvard University Press: Cambridge, MA, USA, 1983. [Google Scholar]
- Simmons, L.W. Female choice in the field cricket Gryllus bimaculatus (De Geer). Anim. Behav. 1986, 34, 1463–1470. [Google Scholar] [CrossRef]
- Eberhard, W.G. Female Control: Sexual Selection by Cryptic Female Choice; Princeton University Press: Cambridge, MA, USA, 1996. [Google Scholar]
- Parker, G.A. Why are there so many tiny sperm—Sperm competition and the maintenance of two sexes. J. Theor. Biol. 1982, 96, 281–294. [Google Scholar] [CrossRef]
- Charnov, E.L. The Theory of Sex Allocation; Princeton University Press: Princeton, NJ, USA, 1982. [Google Scholar]
- Parker, G.A. Sperm competition games—Raffles and roles. Proc. R. Soc. Lond. B Biol. Sci. 1990, 242, 120–126. [Google Scholar] [CrossRef]
- Parker, G.A. Sperm competition games—Sneaks and extra-pair copulations. Proc. R. Soc. Lond. B Biol. Sci. 1990, 242, 127–133. [Google Scholar] [CrossRef]
- Parker, G.A. Sperm competition and the evolution of ejaculates: Towards a theory base. In Sperm Competition and Sexual Selection; Birkhead, T.R., Møller, A.P., Eds.; Academic Press: London, UK, 1998; pp. 3–90. [Google Scholar]
- Parker, G.A.; Pizzari, T. Sperm competition and ejaculate economics. Biol. Rev. 2010, 85, 897–934. [Google Scholar] [CrossRef]
- Birkhead, T.R. Promiscuity; Faber and Faber: London, UK, 2000. [Google Scholar]
- Birkhead, T.R.; Møller, A.P. Sperm Competition in Birds: Evolutionary Causes and Consequences; Academic Press: London, UK, 1992. [Google Scholar]
- Birkhead, T.R.; Hosken, D.J.; Pitnick, S. Sperm Biology: An Evolutionary Perspective; Academic Press: London, UK, 2009. [Google Scholar]
- Simmons, L.W. Sperm Competition and Its Evolutionary Consequences in the Insects; Princeton University Press: Princeton, NJ, USA, 2001. [Google Scholar]
- Parker, G.A. Conceptual developments in sperm competition: A very brief synopsis. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2020, 375, 20200061. [Google Scholar] [CrossRef]
- Simmons, L.W.; Fitzpatrick, J.L. Sperm wars and the evolution of male fertility. Reproduction 2012, 144, 519–534. [Google Scholar] [CrossRef]
- Lüpold, S.; de Boer, R.A.; Evans, J.; Tomkins, J.; Fitzpatrick, J.L. How sperm competition shapes the evolution of testes and sperm: A meta-analysis. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2020, 375, 20200064. [Google Scholar] [CrossRef]
- Parker, G.A. The evolution of expenditure on testes. J. Zool. 2016, 298, 3–19. [Google Scholar] [CrossRef]
- Kelly, C.D.; Jennions, M.D. Sexual selection and sperm quantity: Meta-analyses of strategic ejaculation. Biol. Rev. 2011, 86, 863–884. [Google Scholar] [CrossRef] [PubMed]
- DelBarco-Trillo, J. Adjustment of sperm allocation under high risk of sperm competition across taxa: A meta-analysis. J. Evol. Biol. 2011, 24, 1706–1714. [Google Scholar] [CrossRef] [PubMed]
- Poiani, A. Complexity of seminal fluid: A review. Behav. Ecol. Sociobiol. 2006, 60, 289–310. [Google Scholar] [CrossRef]
- Avila, F.W.; Sirot, L.K.; LaFlamme, B.A.; Rubinstein, C.D.; Wolfner, M.F. Insect Seminal Fluid Proteins: Identification and Function. Annu. Rev. Entomol. 2011, 56, 21–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sirot, L.K.; Wong, A.; Chapman, T.; Wolfner, M.F. Sexual conflict and seminal fluid proteins: A dynamic landscape of sexual interactions. Cold Spring Harb. Perspect. Biol. 2015, 7, a017533. [Google Scholar] [CrossRef]
- Wigby, S.; Brown, N.; Allen, S.; Misra, S.; Sitnik, J.; Sepil, I.; Clark, A.; Wolfner, M. The Drosophila seminal proteome and its role in postcopulatory sexual selection. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2020, 375, 20200072. [Google Scholar] [CrossRef]
- Ramm, S.A. Seminal fluid and accessory male investment in sperm competition. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2020, 375, 20200068. [Google Scholar] [CrossRef]
- Snook, R.R. Sperm in competition: Not playing by the numbers. Trends Ecol. Evol. 2005, 20, 46–53. [Google Scholar] [CrossRef]
- Pitnick, S.; Hosken, D.J. Postcopulatory sexual selection. In Evolutionary Behavioral Ecology; Westneat, D.F., Fox, C.W., Eds.; Oxford University Press: Oxford, UK, 2010; pp. 379–399. [Google Scholar]
- Pitnick, S.; Wolfner, M.F.; Dorus, S. Post-ejaculatory modifications to sperm (PEMS). Biol. Rev. 2020, 95, 365–392. [Google Scholar] [CrossRef]
- Lüpold, S.; Pitnick, S. Sperm form and function: What do we know about the role of sexual selection? Reproduction 2018, 155, R229–R243. [Google Scholar] [CrossRef] [PubMed]
- Parker, G.A. The sexual cascade and the rise of pre-ejaculatory (Darwinian) sexual selection, sex roles, and sexual conflict. Cold Spring Harb. Perspect. Biol. 2014, 6, a017509. [Google Scholar] [CrossRef] [Green Version]
- Parker, G.A.; Pizzari, T. Sexual selection: The logical imperative. In Current Perspectives on Sexual Selection; Hoquet, T., Ed. Springer: Berlin, Germany, 2015; pp. 119–163. [Google Scholar] [CrossRef]
- Kalmus, H. Über den Erhaltungswet den phenotypishen (morphologishen) Anisogamie und die Entstehung der ersten Geshlectsuntershiede. Biol. Zent. 1932, 52, 716–736. [Google Scholar]
- Kalmus, H.; Smith, C.A.B. Evolutionary origin of sexual differentiation and the sex-ratio. Nature 1960, 186, 1004–1006. [Google Scholar] [CrossRef]
- Scudo, F.M. The adaptive value of sexual dimorphism. I. Anisogamy. Evolution 1967, 21, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Williams, G.C.; Mitton, J.B. Why reproduce sexually? J. Theor. Biol. 1973, 39, 545–554. [Google Scholar] [CrossRef]
- Williams, G.C. Sex and Evolution; Princeton University Press: Princeton, NJ, USA, 1975. [Google Scholar]
- Baker, R.R.; Parker, G.A. The origin and evolution of sexual reproduction up to the evolution of the male-female phenomenon. Acta Biotheor. 1973, 22, 49–77. [Google Scholar] [CrossRef]
- Lessells, C.M.; Snook, R.R.; Hosken, D.J. 2—The evolutionary origin and maintenance of sperm: Selection for a small, motile gamete mating type. In Sperm Biology; Birkhead, T.R., Hosken, D.J., Pitnick, S., Eds.; Academic Press: London, UK, 2009; pp. 43–67. [Google Scholar] [CrossRef]
- Maynard Smith, J. The Evolution of Sex; Cambridge University Press: Cambridge, UK, 1978. [Google Scholar]
- Maynard Smith, J. Evolution and the Theory of Games; Cambridge University Press: Cambridge, UK, 1982. [Google Scholar]
- Bell, G. The evolution of anisogamy. J. Theor. Biol. 1978, 73, 247–270. [Google Scholar] [CrossRef]
- Charlesworth, B. The population genetics of anisogamy. J. Theor. Biol. 1978, 73, 347–357. [Google Scholar] [CrossRef]
- Parker, G.A. Selection on non-random fusion of gametes during the evolution of anisogamy. J. Theor. Biol. 1978, 73, 1–28. [Google Scholar] [CrossRef]
- Cox, P.A.; Sethian, J.A. Search, encounter rates, and the evolution of anisogamy. Proc. Natl. Acad. Sci. USA 1984, 81, 6078–6079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Randerson, J.P.; Hurst, L.D. The uncertain evolution of the sexes. Trends. Ecol. Evol. 2001, 16, 571–579. [Google Scholar] [CrossRef]
- Bulmer, M.G.; Luttikhuizen, P.C.; Parker, G.A. Survival and anisogamy. Trends Ecol. Evol. 2002, 17, 357–358. [Google Scholar] [CrossRef]
- Bulmer, M.G.; Parker, G.A. The evolution of anisogamy: A game-theoretic approach. Proc. R. Soc. B-Biol. Sci. 2002, 269, 2381–2388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Togashi, T.; Cox, P.A. (Eds.) The Evolution of Anisogamy: A Fundamental Phenomenon Underlying Sexual Selection; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- Parker, G.A. The origin and maintenance of two sexes (anisogamy), and their gamete sizes by gamete competition. In The Evolution of Anisogamy: A Fundamental Phenomenon Underlying Sexual Selection; Togashi, T., Cox, P.A., Eds.; Cambridge University Press: Cambridge, UK, 2011; pp. 17–74. [Google Scholar]
- Levitan, D.R. Effects of gamete traits on fertilization in the sea and the evolution of sexual dimorphism. Nature 1996, 382, 153–155. [Google Scholar] [CrossRef]
- Iyer, P.; Roughgarden, J. Gametic conflict versus contact in the evolution of anisogamy. Popul Biol 2008, 73, 461–472. [Google Scholar] [CrossRef]
- Yang, J.-N. Cooperation and the evolution of anisogamy. J. Theor. Biol. 2010, 264, 24–36. [Google Scholar] [CrossRef]
- Lehtonen, J.; Kokko, H. Two roads to two sexes: Unifying gamete competition and gamete limitation in a single model of anisogamy evolution. Behav. Ecol. Sociobiol. 2011, 65, 445–459. [Google Scholar] [CrossRef]
- Parker, G.A.; Lehtonen, J. Gamete evolution and sperm numbers: Sperm competition versus sperm limitation. Proc. R. Soc. B-Biol. Sci. 2014, 281, 20140836. [Google Scholar] [CrossRef]
- Lehtonen, J.; Parker, G.A. Evolution of the two sexes under internal fertilization and alternative evolutionary pathways. Am. Nat. 2019, 193, 702–716. [Google Scholar] [CrossRef]
- Randerson, J.P.; Hurst, L.D. A comparative test of a theory for the evolution of anisogamy. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2001, 268, 879–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanschen, E.R.; Herron, M.D.; Wiens, J.J.; Nozaki, H.; Michod, R.E. Multicellularity drives the evolution of sexual traits. Am. Nat. 2018, 192, E93–E105. [Google Scholar] [CrossRef] [PubMed]
- da Silva, J. The evolution of sexes: A specific test of the disruptive selection theory. Ecol. Evol. 2018, 8, 207–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, G. The Masterpiece of Nature: The Evolution and Genetics of Sexuality; Croom Helm: London, UK, 1982. [Google Scholar]
- da Silva, J.; Drysdale, V.L. Isogamy in large and complex volvocine algae is consistent with the gamete competition theory of the evolution of anisogamy. Proc. R. Soc. B Biol. Sci. 2018, 285, 20181954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biological Reviews, 200th Anniversary Collection. Available online: https://onlinelibrary.wiley.com/page/journal/1469185x/homepage/200th-anniversary-collection (accessed on 31 January 2021).
- Behavioural Ecology, Collection: Sperm Competition and its Evolutionary Consequences, Virtual Issues. Available online: https://academic.oup.com/beheco/pages/sperm-competition (accessed on 31 January 2021).
- Philosophical Transactions of the Royal Society B, Theme Issue: Fifty Years of Sperm Competition. Available online: https://royalsocietypublishing.org/toc/rstb/375/1813 (accessed on 31 January 2021).
- Cells, Special Issue: Origin and Evolution of Sperm Cells—An Issue in Honor of Geoff A. Parker. Available online: https://www.mdpi.com/journal/cells/special_issues/honorary_sperm (accessed on 31 January 2021).




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parker, G.A. How Soon Hath Time… A History of Two “Seminal” Publications. Cells 2021, 10, 287. https://doi.org/10.3390/cells10020287
Parker GA. How Soon Hath Time… A History of Two “Seminal” Publications. Cells. 2021; 10(2):287. https://doi.org/10.3390/cells10020287
Chicago/Turabian StyleParker, Geoff A. 2021. "How Soon Hath Time… A History of Two “Seminal” Publications" Cells 10, no. 2: 287. https://doi.org/10.3390/cells10020287
APA StyleParker, G. A. (2021). How Soon Hath Time… A History of Two “Seminal” Publications. Cells, 10(2), 287. https://doi.org/10.3390/cells10020287